
Развитие нервной системы у беспозвоночных животных
3. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ФИЛОГЕНЕЗЕ
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. У одного и того же типа животных нервные клетки могут одновременно и независимо происходить из трех разных зародышевых листков. Полигенез нервных клеток беспозвоночных является основой разнообразия медиаторных механизмов их нервной системы.
Нервная система впервые появляется у кишечнополостных животных. Кишечнополостные — это двухслойные животные. Их тело представляет собой полый мешок, внутренняя полость которого является пищеварительной полостью. Нервная система кишечнополостных принадлежит к диффузному типу. Каждая нервная клетка в ней длинными отростками соединена с несколькими соседними, образуя нервную сеть. Нервные клетки кишечнополостных не имеют специализированных поляризованных отростков. Их отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками диффузной нервной системы бывают нескольких типов. Существуют плазматические контакты, обеспечивающие непрерывность сети (анастомозы). Появляются и щелевидные контакты между отростками нервных клеток, подобные синапсам. Причем среди них существуют контакты, в которых синаптические пузырьки располагаются по обе стороны контакта — так называемые симметричные синапсы, а есть и несимметричные синапсы: в них везикулы располагаются только с одной стороны щели.
Нервные клетки типичного кишечнополостного животного гидры равномерно распределены по поверхности тела, образуя некоторые скопления в районе ротового отверстия и подошвы (рис. 8). Диффузная нервная сеть проводит возбуждение во всех направлениях. При этом волну распространяющегося возбуждения сопровождает волна мышечного сокращения.
Рис. 8. Схема строения диффузной нервной системы кишечнополостного животного:
1 — ротовое отверстие; 2 — щупальце; 3 — подошва
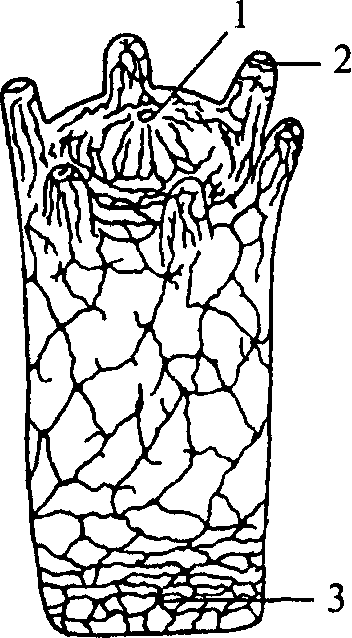
Рис. 9. Схема строения диффузностволовой нервной системы турбеллярии:
1 — нервный узел; 2 — глотка; 3 — брюшной продольный ствол; 4 — боковой нервный ствол
Следующим этапом развития беспозвоночных является появление трехслойных животных — плоских червей. Подобно кишечнополостным они имеют кишечную полость, сообщающуюся с внешней средой ротовым отверстием. Однако у них появляется третий зародышевый слой — мезодерма и двусторонний тип симметрии. Нервная система низших плоских червей принадлежит диффузному типу. Однако из диффузной сети уже обособляются несколько нервных стволов (рис. 9, 3, 4).
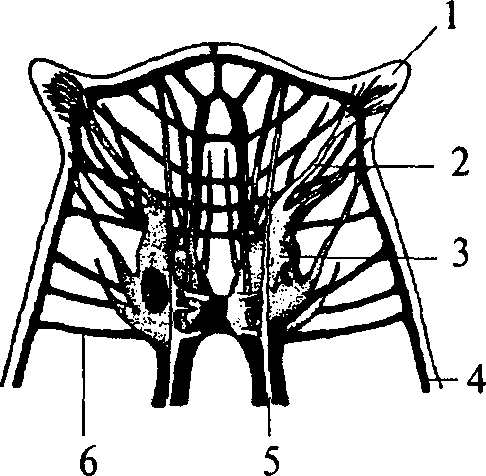
У свободно живущих плоских червей нервный аппарат приобретает черты централизации. Нервные элементы собираются в несколько продольных стволов (рис. 10, 4, 5) (для самых высокоорганизованных животных характерно наличие двух стволов), которые соединяются между собой поперечными волокнами (комиссурами) (рис. 10, 6). Упорядоченная таким образом нервная система называется ортогоном. Стволы ортогона представляют собой совокупность нервных клеток и их отростков (рис. 10).
1 — щупальцевидный вырост; 2 — нерв, иннервирующий вырост; 3 — мозговой ганглий; 4 — боковой продольный нервный ствол; 5 — брюшной продольный нервный ствол; 6 — комиссура
ганглия появляются длинные отростки, идущие в продольные стволы ортогона (рис. 10, 4, 5).
Таким образом, ортогон представляет собой первый шаг к централизации нервного аппарата и его цефализации (появлению мозга). Централизация и цефализация являются результатом развития сенсорных (чувствительных) структур.
Следующим этапом развития беспозвоночных животных является появление сегментированных животных — кольчатых червей. Их тело метамерно, т.е. состоит из сегментов. Структурной основой нервной системы кольчатых червей является ганглий — парное скопление нервных клеток, расположенных по одному в каждом сегменте. Нервные клетки в ганглии размещаются по периферии. Центральную его часть занимает нейропиль — переплетение отростков нервных клеток и глиальные клетки. Ганглий расположен на брюшной стороне сегмента под кишечной трубкой. Он посылает свои чувствительные и двигательные волокна в свой сегмент и в два соседних. Таким образом, каждый ганглий имеет три пары боковых нервов, каждый из которых является смешанным и иннервирует свой сегмент. Приходящие с периферии чувствительные волокна попадают в ганглий через вентральные корешки нервов. Двигательные волокна выходят из ганглия по дорсальным корешкам нервов. Соответственно этому чувствительные нейроны расположены в вентральной части ганглия, а двигательные — в дорсальной. Кроме того, в ганглии есть мелкие клетки, иннервирующие внутренние органы (вегетативные элементы), они расположены латерально — между чувствительными и двигательными нейронами. Среди нейронов чувствительной, двигательной или ассоциативной зон ганглиев кольчатых червей не обнаружено группирования элементов, нейроны распределены диффузно, т.е. не образуют центров.
Ганглии кольчатых червей соединены между собой в цепочку. Каждый последующий ганглий связан с предыдущим при помощи
1 — надглоточный нервный ганглий;
2 — подглоточный нервный ганглий;
3 — сложный слившийся ганглий грудного сегмента; 4 — брюшной ганглий; 5 — периферический нерв; 6 — коннектива
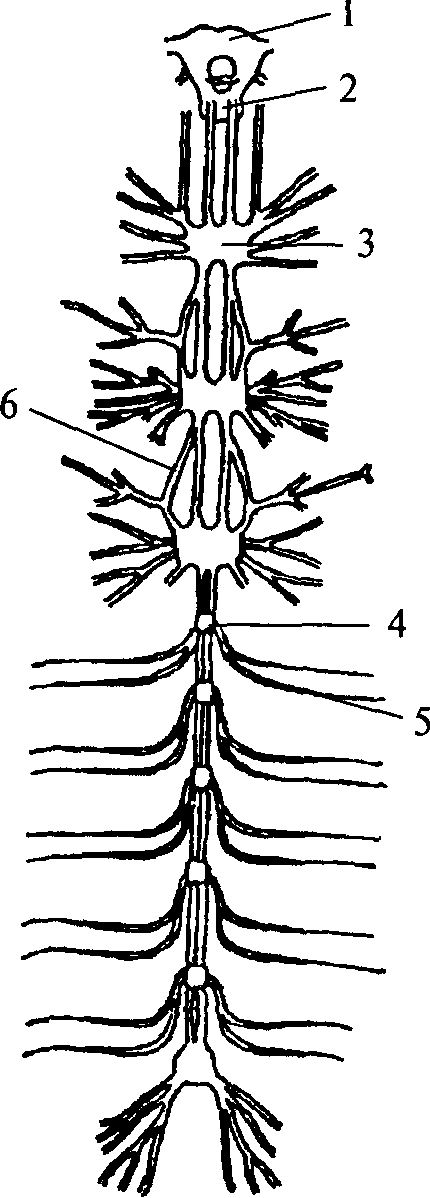
нервных стволов, которые называются коннективами. На переднем конце тела кольчатых червей два слившихся ганглия образуют крупный подглоточный нервный узел. Коннективы от подглоточного нервного узла, огибая глотку, вливаются в надглоточный нервный узел, который является самой ростральной (передней) частью нервной системы. В состав надглоточного нервного ганглия входят только чувствительные и ассоциативные нейроны. Двигательных элементов там не обнаружено. Таким образом, надглоточный ганглий кольчатых червей является высшим ассоциативным центром, он осуществляет контроль над подглоточным ганглием. Подглоточный ганглий контролирует нижележащие узлы, он имеет связи с двумя-тремя последующими ганглиями, тогда как остальные ганглии брюшной нервной цепочки не образуют связей длинней, чем до соседнего ганглия.
В филогенетическом ряду кольчатых червей есть группы с хорошо развитыми органами чувств (многощетинковые черви). У этих животных в надглоточном ганглии обособляются три отдела. Передний отдел иннервирует щупальца, средняя часть иннервирует глаза и антенны. И наконец, задняя часть развивается в связи с совершенствованием химических органов чувств.
Сходную структуру имеет нервная система членистоногих, т.е. построена по типу брюшной нервной цепочки, однако может достигать высокого уровня развития (рис. 11). Она включает в себя значительно развитый надглоточный ганглий, выполняющий функ-
1 — грибовидное тело; 2 — протоцеребрум; 3 — зрительная лопасть; 4 — дейтоцеребрум; 5 — тритоцеребрум
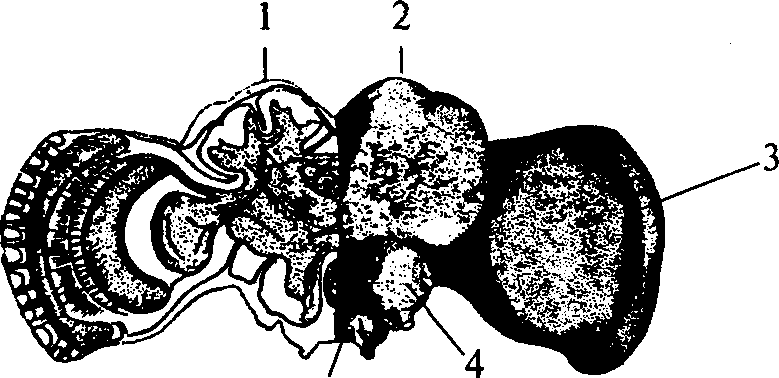
цию мозга, подглоточный ганглий, управляющий органами ротового аппарата, и сегментарные ганглии брюшной нервной цепочки. Ганглии брюшной нервной цепочки могут сливаться между собой, образуя сложные ганглиозные массы.
Головной мозг членистоногих состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Сложным строением отличается мозг насекомых. Особенно важными ассоциативными центрами насекомых являются грибовидные тела, располагающиеся на поверхности протоцеребрума, причем чем более сложным поведением характеризуется вид, тем более развиты у него грибовидные тела. Поэтому наибольшего развития грибовидные тела достигают у общественных насекомых (рис. 12).
Практически во всех отделах нервной системы членистоногих существуют нейросекреторные клетки. Нейросекреты играют важную регулирующую роль в гормональных процессах членистоногих.
В процессе эволюции первоначально диффузно расположенные биполярные нейросекреторные клетки воспринимали сигналы либо отростками, либо всей поверхностью клетки, затем сформировались нейросекреторные центры, нейросекреторные тракты и нейросекреторные контактные области. В последующем произошла специализация нервных центров, увеличилась степень надежности во взаимоотношениях двух основных регуляторных систем (нервной и гуморальной) и сформировался принципиально новый этап регуляции — подчинение нейросекреторным центрам периферических эндокринных желез.
1 — церебральная комиссура; 2 — церебральные ганглии; 3 — педальные ганглии; 4 — коннектива; 5 — висцеральные ганглии
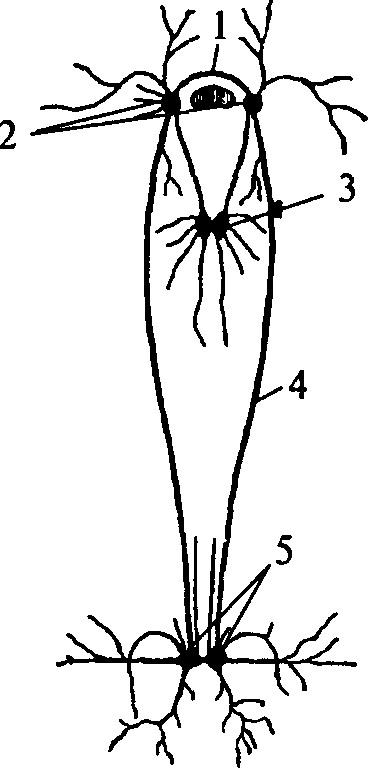
Нервная система моллюсков также имеет ганглионарное строение (рис. 13). У простейших представителей типа она состоит из нескольких пар ганглиев. Каждая пара ганглиев управляет определенной группой органов: ногой, висцеральными органами, легкими и т.д. — и расположена рядом с иннервируемыми органами или внутри их. Одноименные ганглии попарно соединены между собой комиссурами. Кроме того, каждый ганглий связан длинными коннективами с церебральным комплексом ганглиев.
У более высокоорганизованных моллюсков (головоногие) нервная система преобразуется (рис. 14). Ганглии ее сливаются и образуют общую окологлоточную массу — головной мозг. От заднего отдела головного мозга отходят два крупных мантийных нерва и образуют два больших звездчатых ганглия. Таким образом, у головоногих наблюдается высокая степень цефализации.
Этапы развития центральной нервной системы
Появление многоклеточных организмов явилось первичным стимулом для дифференциации систем связи, которые обеспечивают целостность реакций организма, взаимодействие между его тканями и органами. Это взаимодействие может осуществляться как гуморальным путем посредством поступления гормонов и продуктов метаболизма в кровь, лимфу и тканевую жидкость, так и за счет функции нервной системы, которая обеспечивает быструю передачу возбуждения, адресованного к вполне определенным мишеням.
Нервная система беспозвоночных животных
Нервная система как специализированная система интеграции на пути структурного и функционального развития проходит через несколько этапов, которые у первично- и вторичноротых животных могут характеризоваться чертами параллелизма и филогенетической пластичностью выбора.
Среди беспозвоночных наиболее примитивный тип нервной системы в виде диффузной нервной сети встречается у типа кишеч-нополостных. Их нервная сеть представляет собой скопление муль-типолярных и биполярных нейронов, отростки которых могут перекрещиваться, прилегать друг к другу и лишены функциональной дифференциации на аксоны и дендриты. Диффузная нервная сеть не разделена на центральный и периферический отделы и может быть локализована в эктодерме и энтодерме.
Эпидермальные нервные сплетения, напоминающие нервные сети кишечнополостных, могут быть обнаружены и у более высоко организованных беспозвоночных (плоские и кольчатые черви), однако здесь они занимают соподчиненное положение по отношению к центральной нервной системе (ЦНС), которая выделяется как самостоятельный отдел.
В качестве примера такой централизации и концентрации нервных элементов можно привести ортогональную нервную систему плоских червей. Ортогон высших турбеллярий представляет собой упорядоченную структуру, которая состоит из ассоциативных и двигательных клеток, формирующих вместе несколько пар продольных тяжей, или стволов, соединенных большим числом поперечных и кольцевых комиссуральных стволов. Концентрация нервных элементов сопровождается их погружением в глубь тела.
Плоские черви являются билатерально симметричными животными с четко выраженной продольной осью тела. Движение у свободноживущих форм осуществляется преимущественно в сторону головного конца, где концентрируются рецепторы, сигнализирующие о приближении источника раздражения. К числу таких рецепторов турбеллярий относятся пигментные глазки, обонятельные ямки, ста-тоцист, чувствительные клетки покровов , наличие которых способствует концентрации нервной ткани на переднем конце тела. Этот процесс приводит к формированию головного ганглия, который, по меткому выражению Ч. Шеррингтона, можно рассматривать как ганглиозную надстройку над системами рецепции на расстоянии.
Ганглионизация нервных элементов получает дальнейшее развитие у высших беспозвоночных, кольчатых червей, моллюсков и членистоногих. У большинства кольчатых червей брюшные стволы ганглионизированы таким образом, что в каждом сегменте тела формируется по одной паре ганглиев, соединенных коннективами с другой парой, расположенной в соседнем сегменте.
Ганглии одного сегмента у примитивных аннелид соединены между собой поперечными комиссурами, и это приводит к образованию лестничной нервной системы,. В более продвинутых отрядах кольчатых червей наблюдается тенденция к сближению брюшных стволов вплоть до полного слияния ганглиев правой и левой сторон и перехода от лестничной к цепочечной нервной системе. Идентичный, цепочечный тип строения нервной системы существует и у членистоногих с различной выраженностью концентрации нервных элементов, которая может осуществляться не только за счет слияния соседних ганглиев одного сегмента, но и при слиянии последовательных ганглиев различных сегментов.
Эволюция нервной системы беспозвоночных идет не только по пути концентрации нервных элементов, но и в направлении усложнения структурных взаимоотношений в пределах ганглиев. Не случайно в современной литературе отмечается тенденция сравнивать брюшную нервную цепочку со спинным мозгом позвоночных животных. Как и в спинном мозгу, в ганглиях обнаруживается поверхностное расположение проводящих путей, дифференциация нейропиля на моторную, чувствительную и ассоциативные области. Это сходство, являющееся примером параллелизма в эволюции тканевых структур, не исключает, однако, своеобразия анатомической организации. Так, например, расположение туловищного мозга кольчатых червей и членистоногих на брюшной стороне тела обусловило локализацию моторного нейропиля на дорсальной стороне ганглия, а не на вентральной, как это имеет место у позвоночных животных.
Процесс ганглионизации у беспозвоночных может привести к формированию нервной системы разбросанно-узлового типа, которая встречается у моллюсков. В пределах этого многочисленного типа имеются филогенетически примитивные формы с нервной системой, сопоставимой с ортогоном плоских червей (боконервные моллюски), и продвинутые классы (головоногие моллюски), у которых слившиеся ганглии формируют дифференцированный на отделы мозг.
Прогрессивное развитие мозга у головоногих моллюсков и насекомых создает предпосылку для возникновения своеобразной иерархии командных систем управления поведением. Низший уровень интеграции в сегментарных ганглиях насекомых и в подглоточной массе мозга моллюсков служит основой для автономной деятельности и координации элементарных двигательных актов. В то же время мозг представляет собой следующий, более высокий уровень интеграции, где могут осуществляться межанализаторный синтез и оценка биологической значимости информации. На основе этих процессов формируются нисходящие команды, обеспечивающие вариантность запуска нейронов сегментарных центров. Очевидно, взаимодействие двух уровней интеграции лежит в основе пластичности поведения высших беспозвоночных, включающего врожденные и приобретенные реакции.
В целом, говоря об эволюции нервной системы беспозвоночных, было бы упрощением представлять ее как линейный процесс. Факты, полученные в нейроонтогенетических исследованиях беспозвоночных, позволяют допустить множественное (полигенетическое) происхождение нервной ткани беспозвоночных. Следовательно, эволюция нервной системы беспозвоночных могла идти широким фронтом от нескольких источников с изначальным многообразием.
На ранних этапах филогенетического развития сформировался второй ствол эволюционного древа, который дал начало иглокожим и хордовым. Основным критерием для выделения типа хордовых является наличие хорды, глоточных жаберных щелей и дорсального нервного тяжа — нервной трубки, представляющей собой производное наружного зародышевого листка — эктодермы. Трубчатый тип нервной системы позвоночных по основным принципам организации отличен от ганглионарного или узлового типа нервной системы высших беспозвоночных.
Нервная система, служащая для проведения раздражений и интеграции деятельности организма, возникает на основе проведения раздражений и интеграции поведения простейших посредством градиентов: функции проведения и интеграции в ходе эволюции откладываются у организмов в самой их структуре.
Нервная система появляется впервые у кишечнополостных. В своём развитии она проходит несколько этапов, или ступеней. Первоначальным, наиболее примитивным типом нервной системы является диффузная нервная система.Она порождает недифференцированный способ реагирования на раздражение, который встречается, например, у медузы.
Диффузная нервная система медузы
В дальнейшем развитии живых существ и их нервной системы начинается процесс централизации нервной системы (у червей), который далее идёт по двум расходящимся линиям: из них одна ведёт к высшим беспозвоночным, другая — к позвоночным. Эволюция приводит, с одной стороны, к образованию сначала так называемой узловой нервной системы. Для неё характерно сплетение, концентрация нервных клеток в узлах, которые по преимуществу осуществляют регуляцию реакций животного. Этот тип нервной системы отчётливо представлен у кольчатых червей. У некоторых из них, например у дождевого червя, отдельные нервные узлы соединены между собой, образуя как бы цепочку. Такая цепочечная нервная система отличается сегментарностью, выражающейся в том, что каждая пара нервных узлов обслуживает прилегающий к ней сегмент тела. Вместе с тем уже у червей начинает выделяться головной узел, приобретающий господствующее, доминирующее значение. У животных, обладающих узловой нервной системой, впервые появляется реакция, имеющая характер рефлекса.
Цепочечная нервная система дождевого червя
У членистоногих (у пчёл, муравьёв) — на высших ступенях развития беспозвоночных — головной мозг приобретает уже очень сложное строение; в нём дифференцируются отдельные части (грибовидные тельца), в которых происходят довольно сложные процессы переключений. В соответствии с этой относительно сложной организацией нервной системы у членистоногих, в частности у пчёл, у муравьёв, наблюдаются и довольно сложные формы поведения и психической деятельности. Эта деятельность имеет, однако, по преимуществу инстинктивный характер.
Уже у беспозвоночных сказываются основные тенденции развития нервной системы, имеющие существенное значение и для развития её психических функций. Эти тенденции заключаются в прогрессирующей централизации, цефализации и иерархизации нервной системы. Централизация нервной системы проявляется в сосредоточении нервных элементов в определённых местах, в образовании (в вполне выраженной форме у червей) ганглиев, в которых скопляется, централизуется множество ганглиозных нервных клеток; цефализация нервной системы заключается в преимущественном сосредоточении в особо высокой дифференцировке нервной системы на головном конце тела; иерархизация нервной системы выражается в подчинении одних участков или частей нервной системы другим, которые приобретают господствующее, или доминирующее, значение.
Узловая нервная система пчелы
В связанной с этим развитием нервной системы эволюции её функций проявляется существенная закономерность, заключающаяся в прогрессирующей специализации реакций. Вначале внешнее раздражение вызывает в ответ диффузную реакцию, как бы массовое действие (mass action — по Г. Э. Когхиллу), затем происходит специализация реакций, т. е. выделение местных специализированных реакций отдельных частей тела. Захватывая в какой-то мере всю нервную систему, возбуждение в результате внутрицентральных взаимодействий направляется более избирательно по некоторому числу нервных путей. В результате возникают более специализированные реакции, лучше приспособленные для достижения определённого эффекта.
Эти тенденции в развитии нервной системы приобретают ещё более глубокое и специфическое значение на другой из двух раздваивающихся линий, которая от первичных форм (турбелларий) с нерасчленённым головным ганглием диффузного нервного строения ведёт к трубчатой нервной системе позвоночных.
У позвоночных совершается всё более отчётливая дифференциация нервной системы на периферическую и центральную.Прогресс в развитии позвоночных осуществляется главным образом за счёт развития центральной нервной системы. Центральная нервная система дифференцируется на спинной и головной мозг. Наиболее существенным в развитии центральной нервной системы является эволюция строения и функций головного мозга. В головном мозге дифференцируется мозговой ствол и большие полушария. Большие полушария развиваются в филогенезе из конечного мозга. На ранних стадиях филогенетического развития конечный мозг является органом обонятельных рецепций; вторичные обонятельные центры конечного мозга образуют так называемую древнюю кору,от которой отличают ещё старую кору,состоящую из совокупности образующихся у птиц и главным образом у рептилий третичных обонятельных центров. У последних начинается образование новой коры, неокортекса.
Развитие конечного мозга у позвоночных (по Эддингеру)
I — мозг человека; II — кролика; III — ящерицы; IV — акулы. Чёрным обозначена новая кора, пунктиром — старая обонятельная часть её
В то время как у птиц эволюция конечного мозга идёт в сторону развития центральных ганглиев, у млекопитающих она идёт в сторону развития коры. Значительное развитие коры — неокортекса — является наиболее характерной чертой в развитии мозга млекопитающих; у высших из них, у приматов и особенно у человека, она занимает господствующее положение.
Энцефализация, как основной принцип прогрессивного развития центральной нервной системы, заключается в том, что в ходе эволюции совершается переход функционального управления из спинного мозга через все уровни центральной нервной системы от низших к высшим её этажам или отделам. При этом переходе функций вверх первоначальные центры сводятся на роль лишь передаточных инстанций.
I — полушария головного мозга; II — промежуточный мозг; III — средний мозг; IV — задний мозг; V — продолговатый мозг
Развитие дистантных рецепторов, увеличивая возможности отображения действительности, создаёт предпосылку для развития более совершенно организованных форм поведения. Будучи предпосылкой развития более совершенных форм поведения, в которых психические компоненты начинают играть всё более существенную роль, развитие нервной системы и в частности её рецепторного аппарата является вместе с тем и результатом развития этих форм поведения. Развитие нервной системы и психических функций у животных совершается в процессе эволюции форм их поведения.
Образ жизни и психика
У высших беспозвоночных и у членистоногих, особенно у насекомых — у пчёл и у муравьев, развитие нервной системы достигает значительной степени централизации и цефализации; головные узлы играют доминирующую роль; строение головного мозга в достаточной мере сложно; в нём дифференцируются отдельные части (грибовидные тельца), которые выполняют более или менее сложные функции переключения. Значительной сложностью и относительно высокой организованностью отличаются и психические функции и поведение — особенно у пчёл и муравьёв.
В отличие от Хесса, Фриш пришёл к выводу, что пчёлы различают цвета, а не только яркость. По данным Кюна, спектр у насекомых по сравнению с человеком смещён несколько вправо; в пределах доступного им спектра пчёлы, по данным Кюна, различают 4 цветовых качества.
У насекомых имеется ассоциативная память. Как показали в частности опыты Фриша, пчёл можно приучить к тому, чтобы они прилетали за мёдом на площадки определённого цвета. Таким образом, и в поведении высших беспозвоночных, у которых фиксированные инстинктивные формы поведения особенно сильно выражены, имеются и элементы пластичности, некоторые, хотя и ограниченные, возможности выработки индивидуально-изменчивых форм поведения. Однако у насекомых, у пчёл и муравьёв, господствующими являются инстинктивные формы поведения, достигающие у них, как о том свидетельствует жизнь пчёл и муравьёв, большой сложности.
В филогенезе низших позвоночных конечный мозг служит первоначально высшим органом обонятельных рецепций и их координации с нижележащими отделами центральной нервной системы; вторичные обонятельные центры образуют старую кору. Обоняние является главным органом дифференциации внешнего мира и ориентировки в нём. Лишь у рептилий появляется новая кора (неокортекс), не являющаяся уже непосредственно аппаратом обоняния, однако и у них, как и у всех нижестоящих позвоночных, обонятельные функции ещё преобладают. Дальнейшее развитие кора получает у млекопитающих, превращаясь в орган всё более высокой корреляции различных восприятий всё более сложного поведения.
В развитии позвоночных снова выступает принцип непрямолинейного развития по расходящимся линиям. Из развивающегося в процессе онтогенеза позвоночных конечного мозга — коры и центральных ганглиев у одних преобладающее развитие получает кора, у других — центральные ганглии. Эволюция конечного мозга в сторону преобладания центральных ганглиев наблюдается у птиц, в сторону развития и всё большего преобладания коры — у млекопитающих. Эта последняя линия, ведущая к приматам и затем к человеку, оказывается более прогрессивной. По этой линии преимущественно развиваются высшие формы индивидуально-изменчивого поведения; по другой линии — у птиц — снова особо значительную роль приобретают структурно фиксированные, инстинктивные формы поведения.
Со слабым развитием коры и преобладанием центральных ганглиев в строении центральной нервной системы у птиц сочетается значительное развитие полушарий большого мозга, знаменующее большой шаг вперёд по сравнению с рептилиями. В полушариях заметное развитие получают зрительные доли и малое — обонятельные, в области чувствительности — значительное развитие зрения и слабое развитие обоняния. Так же слабо развито у птиц осязание, хорошо, как правило, развит слух.
Центральный факт, определяющий и строение птиц и их психику, заключается в их приспособленности к полёту, к лётной жизни в воздухе. Для лётной жизни нужно хорошее развитие зрения (особенно изощрено оно, как известно, у хищных птиц, которые с большой высоты стремглав бросаются на свою жертву). Но воздух вместе с тем значительно более однообразная среда, чем почва, жизнь на которой приводит млекопитающих в соприкосновение с многообразными предметами. В соответствии с этим и деятельность птиц, включая лётные движения, отличается значительным однообразием, шаблонностью, относительно малой вариативностью. [См. В. М. Боровский, Психическая деятельность животных, М. 1936.] Некоторые птицы, несомненно, обнаруживают довольно хорошую обучаемость, но в общем у птиц очень выступает роль инстинктивных форм поведения. Наиболее характерным для птиц является сочетание относительно весьма шаблонных действий с мало вариативными двигательными возможностями и очень развитого восприятия (в частности зрительного). Благодаря последнему некоторые инстинктивные действия птиц производят впечатление действий, находящихся на грани инстинкта и интеллекта, — как, например, поведение вороны в вышеприведённом опыте с орехом и горшочком.
Инстинкты птиц — это уже не те инстинкты, что у пчёл или муравьёв, вообще у беспозвоночных. Сам инстинкт, таким образом, изменяется — на разных ступенях развития он иной; вместе с тем изменяется и соотношение инстинктивных и индивидуально-изменчивых форм поведения: у птиц — особенно у некоторых — научаемость достигает уже значительного уровня.
У млекопитающих, развитие которых ведёт к приматам и затем к человеку, значительное развитие получает новая кора — неокортекс. В поведении млекопитающих господствующее значение получают индивидуально приобретаемые, изменчивые формы поведения.
Ярким проявлением непрямолинейного хода развития, совершающегося по расходящимся линиям, служит при этом тот факт, что ни у одного из млекопитающих вплоть до приматов острота зрения на расстоянии не достигает того уровня, что у птиц. У низших млекопитающих существенную роль в поведении при ориентировке в окружающем играет ещё обоняние, в частности у крыс, а также у собак. Несомненно отчасти поэтому собаки хуже справляются с задачами, требующими зрительного охвата ситуации.
Высшего своего развития психические функции достигают у приматов. Центральный факт, которым определяются и строение мозга и психические функции обезьян, заключается в древесном образе жизни обезьян (а не в будто бы самодовлеющем развитии психических способностей или таком же самодовлеющем развитии в строении мозга). Жизнь на деревьях расширяет поле зрения; значение обоняния уменьшается, роль зрения возрастает.
Опыты Н. Н. Ладыгиной-Котс
Обусловленный этим образом жизни на деревьях способ передвижения обезьян привёл к тому, что обезьяны стали переходить к прямой походке; рука начала выполнять у них иные функции, чем нога; она стала служить для хватания; в ней выделяется большой палец, приспособленный для хватания веток, и она становится пригодной для схватывания и держания различных предметов и манипулирования ими. Развитие у обезьян руки и зрения, способности манипулировать предметами под контролем зрения, позволяющего подмечать те изменения в окружающем, которые вносит в него собственное действие, создаёт у обезьян основные биологические предпосылки для развития интеллекта.
К манипулированию над предметами и к зоркому их рассматриванию стимулирует обезьян и то, что они питаются орехами, внутренним содержанием плодов, сердцевиной стеблей, так что пищу им приходится извлекать, производя так сказать практический анализ вещей. Образ жизни обезьян определяет доступный им образ познания. Умение собрать различные части, составить из различных предметов новое целое, приложить один предмет к другому в качестве орудия, т. е. склонность и способность к практическому синтезу, по данным Н. Ю. Войтониса, у низших обезьян ещё не развита.
Специальные наблюдения и экспериментальные исследования показали, что уже для низших обезьян характерна способность зорко подмечать каждую деталь окружающих их предметов и склонность, манипулируя ими, выделять эти детали; при этом их привлекает сама новизна предметов.
Изучению психики приматов, особенно же антропоидов, человекоподобных обезьян, посвящено множество исследований. Из работ советских авторов нужно отметить прежде всего посвящённые высшим обезьянам исследования Н. Н. Ладыгиной-Котс. Изучение поведения обезьян ведётся также в Колтушах в лаборатории акад. Л. А. Орбели. Из работ зарубежных авторов особенное значение имеют работы Р. М. Иеркса, В. Келера, П. Гильома (Guillaume) и Э. Мейерсона и ряд других.
Из этих последних работ мы специально остановимся на пользующихся особенно широкой известностью исследованиях В. Келера.
Для правильной оценки исследований Келера существенно отделить объективное содержание его экспериментальных данных от той гештальтистской теории, из которой он исходит.
Данные новейших исследований, в частности советских (Н. Ю. Войтонис, Г. С. Рогинский), а также зарубежных (Л. Верлен) свидетельствуют, во-первых, о том, что В. Келер в ходе своих экспериментов, очевидно, недооценивал обезьян. Оказалось, что даже низшие обезьяны способны при надлежащих условиях разрешать некоторые задачи, которые у Келера представлялись недоступными антропоидам. Так, в частности в опытах Рогинского даже низшие обезьяны, несколько приучившись к тесёмкам и верёвкам, выбирали из многих верёвок и тесёмок только те, которые были привязаны к приманке, независимо от того, каково было их расположение. Эти опыты не удавались обезьянам только при очень возбуждённом их состоянии. По-видимому, Келер в опытах над антропоидами сделал окончательные выводы об их возможностях на основании данных, полученных в той стадии, когда обезьяны ещё не освоились с верёвками или находились в особенно возбуждённом состоянии.
Схема расположения нитей в опытах В. Келера
Таким образом, если обезьянам, как мы видели выше, доступны действия, по своей внешней эффективности превосходящие очерченные В. Келером возможности, то по своей внутренней психологической природе их поведение более примитивно, чем утверждал Келер. Однако этот вопрос об интеллекте антропоидов требует ещё дальнейших пристальных исследований. Необходимо при этом учитывать, что, судя по всем данным, индивидуальные различия между антропоидами чрезвычайно велики, поэтому сделать общие выводы на основании наблюдений за одной или двумя обезьянами едва ли возможно.
Структурный принцип гештальтистов внёс в проблематику сравнительной психологии ряд противоречивых тенденций. Развивая в полемике против теории трёх ступеней К. Бюлера гештальтистскую концепцию психологического развития, К. Коффка с полной определённостью формулирует её следующим образом:
Принцип, выдвинутый В. Келером для объяснения интеллекта в его специфическом отличии от других низших форм, объявляется общим для всех форм поведения. Этот результат заложен в самом принципе, из которого исходило гештальтистское понимание интеллекта у Келера. Принцип целостности структуры действительно не в состоянии отдифференцировать интеллект, разумное поведение от низших форм поведения, в частности от инстинкта. Только что установленные грани опять стираются в результате того, что за попыткой продвинуть низшую границу вверх последовала попытка так же неправомерно сдвинуть верхнюю границу вниз.
В. Келер выявил осмысленное поведение обезьян как новый специфический тип поведения, в отличие от случайного, неосмысленного поведения по методу проб и ошибок торндайковских животных. Но как только это было сделано, сейчас же обнаружилась тенденция превратить только что установленный новый вид поведения в такую же универсальную форму. Наряду с этой тенденцией выявилась и другая, для которой также исследование Келера послужило отправной точкой. Поскольку Келер совершенно ошибочно признал у своих обезьян интеллект того же вида и рода, что и у человека, создалась чрезвычайно благоприятная ситуация для того, чтобы в менее примитивных, более утончённых и потому опасных формах провести отожествление психики животных и человека. Эта возможность, заложенная в признании интеллекта у обезьян, была реализована отчасти самим Келером, перенесшим свои опыты над обезьянами на детей, и затем его продолжателями, исследовавшими практический интеллект у человека (см. главу о мышлении).
Читайте также:
