
Рефлексы спинного мозга нервные центры этих рефлексов
В СМ представлен ряд жизненно важных нервных центров: регуляции большинства внутренних органов и скелетной мускулатуры; симпатической нервной системы: центр зрачкового рефлекса (С8—Th2), регуляции деятельности сердца (Т1—Т5), слюноотделения (Т2—Т4), регуляции функций почек (Т5—L3); сегментарпо расположены центры, регулирующие функции потовых желез и сосудов, гладких мышц внутренних органов. Парасимпатическая иннервация из СМ (S2—S4) регулирует деятельность всех органов малого таза, мочевого пузыря, части толстой кишки ниже ее левого изгиба, половые органы. У мужчин парасимпатическая иннервация обеспечивает рефлекторный компонент эрекции, у женщин — сосудистые реакции клитора и влагалища. Центры управления скелетной мускулатурой находятся во всех отделах СМ и иннервируют по сегментарному принципу: скелетную мускулатуру шеи (Cl—С4), диафрагмы (СЗ—С5), верхних конечностей (С5—Т2), туловища (ТЗ—L1), нижних конечностей (L2—S5). Повреждение определенного сегмента СМ или его проводящих путей вызывает специфические двигательные или чувствительные нарушения.
СМ выполняет рефлекторную и проводящую функции. Рефлексы СМ делятся на соматические, висцеральные и вегетативные. Соматические рефлексы подразделяются в зависимости от рецепторов, раздражение которых вызывает рефлекс, на проприоцептивные, висцерорецептивные, кожные
(защитные); в зависимости от органов — эффекторов рефлекса — на рефлексы конечностей, брюшные, органов тела. Рефлексы конечностей подразделяются но характеру ответной реакции: сгибательные, разгибательные, ритмические, позно-тонические; количеству переключений: моносинаптические, полисинантические.
Моносинаптические рефлексы растяжения — простейшие рефлексы СМ (рис. 2.8). Вызываются путем быстрого растяжения мышцы, например при ударе по ее сухожилию. Растяжение мышцы приводит к растяжению рецепторов — мышечных веретен. Сенсорные волокна этих рецепторов входят в СМ в составе задних корешков и заканчиваются в передних рогах, где образуют синаптические контакты на двигательных нейронах, иннервирующих растягиваемую мышцу. В центральной части (в ЦНС) дуги рефлексов растяжения имеют один синапс — между чувствительными и двигательными нейронами. Особенность рефлекса заключается в том, что начало и конец рефлекторной дуги связаны с мышцей. Рефлексы наиболее выражены в мышцах-разгибателях. Биологическое значение этих рефлексов заключается в их участии в сохранении статики и положения тела.

Рис. 2.8. Рефлексы спинного мозга:
а — коленный; б — ахиллов; в — подошвенный (рефлекс Бабинского); г — сгибательный предплечья; д — разгибательный предплечья; е — брюшной
В полисинаптической дуге на пути возбуждения от рецептора к эффектору находятся вставочные нейроны. Динамический рефлекс растяжения осуществляется при быстром удлинении мышцы, что приводит к столь же быстрому ее сокращению. После растяжения мышцы до ее новой длины следует слабый статический рефлекс растяжения. Функция статического рефлекса растяжения направлена против сил, вызывающих превышение исходной длины мышцы.
Тормозный сухожильный рефлекс проявляется следующим образом. Сильное растяжение сухожилий мышцы, например при мощном сокращении или действии внешней силы, приводит к ее расслаблению. При таком растяжении возбуждаются сухожильные органы Гольджи — окончания чувствительных волокон группы IB, которые расположены в сухожилиях и воспринимают раздражение. (К волокнам группы В относятся миелинизированные волокна средней толщины со скоростью проведения возбуждения 3—14 м/с.) Эти органы активизируются при увеличении напряжения в мышце. Волокна, следующие от органов Гольджи, входят в СМ и оканчиваются на вставочных нейронах. Последние образуют тормозные синапсы на альфа-мотонейронах, иннервирующих растянутую мышцу. При их разряде в мотонейронах возникают тормозные постсинаптические потенциалы, подавляющие активность мотонейрона, препятствуя возникновению потенциала действия. Сухожильные рефлексы защищают мышцу от повреждения при сильных сокращениях и участвуют в регуляции напряжения в ней. При достижении максимального напряжения возбуждаются сухожильные органы Гольджи, а альфа-мотонейроны, иннервирующие сокращающуюся мышцу, тормозятся, что приводит к расслаблению мышцы.
Сгибательный рефлекс возникает в ответ на сильные раздражения болевых рецепторов конечности и заключается в сокращении сгибательных мышц и одновременном расслаблении.
Висцеромоторные рефлексы возникают при возбуждении афферентных волокон внутренних органов и характеризуются появлением двигательных реакций мышц грудной и брюшной стенки, мышц-разгибателей спины. Их возникновение связано с существованием конвергенции висцеральных и соматических афферентных волокон к одним и тем же интернейронам СМ.
Вегетативные рефлексы заключаются в появлении полисинаптических разрядов в преганглионарных симпатических волокнах в ответ на возбуждение симпатических и соматических чувствительных клеток; в возникновении рефлекторных реакций парасимпатических нейронов в ответ на раздражение чувствительных путей.
В спинном мозге рефлекторные кольца скелетных мышц взаимосвязаны с рефлекторными кольцами внутренних органов. Поэтому при раздражениях рецепторов афферентные нервные импульсы вызывают также возбуждение эфферентных нейронов, иннервирующих сердце, сосуды, железы и другие органы. При этом гормоны и другие химические продукты поступают в кровь и объектом их воздействия становится и центральная нервная система.
Следовательно, в спинном мозге имеются нейроны, вызывающие соматические (двигательные) и вегетативные рефлексы.
Основные свойства нервных центров спинного мозга имеют подчиненное значение, так как ведущая роль в объединении и регулировании всей деятельности организма в его взаимодействии с окружающим миром принадлежит высшему отделу нервной системы — большим полушариям головного мозга.
Определение возбудимости центров спинного мозга
Возбудимость спинного мозга определяется по длительности латентного периода рефлекса и по величине рефлекторной реакции. Латентный период рефлекса значительно больше, чем при раздражении нервно-мышечного препарата. Он складывается из времени проведения возбуждения по афферентному пути, по центральной нервной системе и по эфферентному пути. При детальных исследованиях учитывается также латентный период рецептора и эффектора.
Время прохождения возбуждения по центральной нервной системе или необходимое для перехода возбуждения с афферентных нейронов на эфферентные, называется центральным временем рефлекса, или редуцированным временем рефлекса. Его продолжительность зависит от количества нейронов и синапсов, участвующих в осуществлении данного рефлекса. Оно тем продолжительнее, чем больше нейронов и синапсов имеется на пути проведения возбуждения по центральной нервной системе, т. е. в рефлекторном кольце, так как проведение возбуждения задерживается в каждом нейроне и особенно в каждом синапсе.
Время сухожильных рефлексов самое короткое; например, коленного — 5,3 мс, из которых на проведение возбуждения по периферическим путям приходится 3,1 мс, а на центральное время — 2,2 мс. При повторении раздражения центральное время может уменьшиться до 1 и даже до 0,3 мс. Продолжительность центрального времени рефлекса наибольшая при слабых раздражениях. При сложных соматических рефлексах центральное время рефлекса увеличивается до 10-20 и даже до 500 мс, а при вегетативных рефлексах — до десятков секунд.
Изменения возбудимости. Сразу же после проведения каждого нервного импульса повторные раздражения вызывают в нейронах спинного мозга абсолютную и относительную рефрактерную фазы, а затем наступает экзальтационная фаза. Эти фазы более продолжительны, чем в нервных волокнах.
После экзальтационной фазы возбудимость понижается. Ритм и сила рефлекторной реакции зависят от изменений возбудимости и лабильности нейронов. В связи с возникновением в одних нейронах спинного мозга возбуждения возбудимость их увеличивается, а в связи с возникновением в других нейронах нервной системы торможения возбудимость этих нейронов уменьшается. Следовательно, возбудимость нейронов спинного мозга постоянно изменяется в связи со сменой возбуждения и торможения и поэтому не существует постоянного порога рефлекторного раздражения.
Возбудимость изменяется в зависимости от притока импульсов, поступающих из рецепторов. Синапсы в течение многих дней могут находиться в состоянии резко пониженной возбудимости или, наоборот, в состоянии повышенной возбудимости, что в первом случае сопровождается отсутствием моторных рефлексов, а во втором — усилением рефлекторных движений.
Координирующие механизмы спинного мозга
Согласованность, соподчиненность сокращений отдельных мышц или их групп, основанная на взаимодействии возбуждения и торможения в нервной системе, обозначается как координация.
Побуждение группы нейронов сопровождается изменением функционального состояния соседних нейронов, их торможением. По ному каждый рефлекс — результат взаимодействия возбуждении и торможения в центральной нервной системе, согласованной и соподчиненной деятельности многих нейронов. Именно благодаря координации нейронов, которая возникла в процессе фило- и онтогенетического развития, могут осуществляться тонкие движении, изолированные двигательные рефлексы вместо общих массовых рефлекторных движений, присущих организмам, у которые нервная система находится на низших ступенях фило- и онтогенеза. Координация характеризует не только сложные, но и самые простые рефлексы.
Особенности иррадиации возбуждения в спинном мозге
Возбуждение, поступившее в спинной мозг по афферентному нейрону, распространяется на выше и ниже расположенные сегменты. Этот процесс распространения возбуждения называется иррадиацией.
Иррадиация возбуждения особенно ясно наступает вследствие увеличения силы раздражения. При слабом раздражении рецепторов наблюдается незначительный рефлекс, так как рефлекторное движение ограничивается небольшой группой мышц. При усилении раздражения рецепторов может наступить общая рефлекторная двигательная реакция. Величина рефлекса зависит от количеств двигательных нейронов, находящихся в состоянии возбуждения. При слабом раздражении рецепторов количество возбужденных эфферентных моторных нейронов невелико, а при усилении раздражении рецепторов, благодаря иррадиации возбуждения, и поэтому возбужденных нейронов все более возрастает, и поэтому начинают сокращаться все большие массы мускулатуры.
Иррадиация возбуждении по нервным центрам увеличивается также при возрастании продолжительности раздражения рецепторов, что приводит к возбуждению все большего количества нейронов. Торможение ограничивает иррадиацию возбуждения, пропуская возбуждение по определенным нервным путям.
Различаются две группы мышц, участвующих в координированном движении: 1) синергисты и 2) антагонисты.
Одновременное совместное сокращение синергистов обеспечивает выполнение данного движения, например короткая, длинная и большая мышцы, сокращаясь одновременно, приводят бедро.
Совместное сокращение антагонистов вызывает движения противоположного характера и поэтому делает невозможным выполнение данного движения, например сгибатели и разгибатели, приводящие и отводящие мышцы. При выполнении других движений антагонистические мышцы могут стать синергистами при отведении и приведении или разгибатели обеих ног становятся синергистами во время стояния.
Впервые Л. А. Спиро (1876) обнаружил на спинномозговой лягушке, что раздражение кожи на задней лапке вызывает ее сгибание и торможение сгибания или разгибания на противоположной стороне. Н. А. Миславский (1887) открыл, что кора больших полушарий одновременно возбуждает нервный центр расширения зрачка и тормозит тонус нервного центра, суживающего зрачок. Ч. Шеррингтон (1894) доказал, что возбуждение центров III и IV черепно-мозговых нервов тормозит центр VI нерва. Он назвал этот факт реципрокной иннервацией. В школе Н. Е. Введенского реципрокная иннервация обозначается как сопряженная.
Н. Е. Введенский (1896) установил, каким образом осуществляется взаимодействие двух очагов возбуждения в моторных нервных центрах коры больших полушарий головного мозга. Если раздражать на одном полушарии точку для сгибателей, то возбуждение нервного центра сгибателей на раздражаемой стороне вызывает на противоположной стороне сопряженное торможение нервного центра сгибателей и возбуждение нервного центра разгибателей.
Взаимодействие возбуждения и торможения — общее свойство всей нервной системы — обнаруживается и на спинном мозге. В зависимости от частоты и силы раздражения рецепторов и лабильности нервных центров в них в каждый момент получается возбуждение или торможение, а в соседних центрах в это время возникает торможение или возбуждение, что обеспечивает рефлекторную координацию движений. В результате взаимного сочетания возбуждения и торможения в соседних нервных центрах одни и те же мышцы в каждый момент могут быть антагонистами или синергистами. Один и тот же моторный нейрон при различных условиях раздражения и лабильности может находиться или в состоянии возбуждения или в состоянии торможения.
Господствующий очаг возбуждения вызывает в соседних нервных центрах торможение вследствие одновременной индукции. Чем больше возбуждение в очаге доминанты, тем больше заторможены соседние нервные центры. При осуществлении двух синергических рефлексов, имеющих общий конечный путь, рефлекс усиливается. При раздражении двух различных рецептивных полей вызываются два разных рефлекса, из которых один может вызвать торможение другого. В этом случае в нервной системе одновременно возникают два очага возбуждения, один из которых доминирует над другим.
По теории доминанты, главное значение имеет не чисто анатомический принцип общего конечного пути (Ч. Шеррингтон), а динамика одновременных или последовательных возбуждений в нервных центрах. Чем выше возбудимость и чем больше стойкость возбуждения очага доминанты, тем больше его способность суммировать приходящие к нему импульсы и тем сильнее возбуждение. Каждое слабое подпороговое раздражение создает в очаге доминанты повышение возбудимости и тем самым изменяет результат действия последующих посторонних импульсов. Посторонние для доминирующего очага раздражители, вызывающие другие рефлексы, не только не мешают доминанте, но, наоборот, подкрепляют, усиливают доминанту и тем самым увеличивают торможение в других сопряженных рефлекторных кольцах.
Следовательно, очаг доминанты характеризуется: 1) повышенной возбудимостью; 2) способностью к суммированию и накоплению возбуждения при притоке к нему импульсов, вызывающих другие рефлексы; 3) по мере развития в нем возбуждения способностью тормозить текущие рефлексы, встречаемые на общем конечном пути; 4) стойкостью возбуждения. Длительное возбуждение очага доминанты поддерживается слабыми раздражителями и не исчезает сразу после прекращения их действия (инерция доминанты). Доминанта обладает значительной инерцией, пока не будет выполнен соответствующий доминирующий рефлекс. Когда доминирующий рефлекс осуществляется длительно, то его инерция преодолевается лишь активным торможением с другой достаточно устойчивой доминанты. И пятое свойство — способность к самовозобновлению.
Существует также предел возбудимости, при достижении которого даже очень слабые или редкие раздражения рецептивных полей других рефлексов могут оказаться чрезмерно сильными и частыми и вызвать переход возбуждения очага доминанты в его торможение.
Таким образом, возбуждение и торможение, развивающиеся в нервной системе, рассматриваются как единое целое, а не изолированно. Существенное значение имеет исходное физиологическое состояние нервной системы. Начавшийся рефлекс может либо усиливать, либо тормозить другие рефлексы в зависимости от условий раздражения нервной системы и ее текущей лабильности.
Теория доминанты и рефлекторная теория И. П. Павлова объясняют механизм образования условного рефлекса притягиванием возбуждения из более слабого очага при действии условного раздражителя в более сильный его очаг, который образуется при действии безусловного раздражителя.
Обе теории имеют значение и для психологии.
Теория И. П. Павлова о главенствующем в данный момент сильном очаге возбуждения в коре больших полушарий, который притягивает возбуждение из более слабых очагов возбуждения и по правилу индукции тормозит окружающие участки коры, и теория доминанты раскрывают физиологические механизмы психического акта внимания. Смена одной доминанты другой проявляется в переключении внимания. Психологические исследования подтверждают теорию доминанты и показывают, что разнообразные слабые раздражения усиливают концентрацию внимания и что умственная работа протекает в обычной обстановке лучше, чем в абсолютной тишине. Для педагогов очень важно то, что посторонние раздражения небольшой силы, нарушающие монотонность, увеличивают общий тонус организма и способствуют концентрации внимания.
Нервная система - сложнейшая и интереснейшая во всем организме. Головной, спинной мозг, а также нервные волокна обеспечивают целостность нашего организма и поддерживают его функционирование. Одна из основных функций нервной системы - защита организма от внешних раздражителей. Это возможно благодаря наличию спинальных рефлексов.

Что такое рефлекс?
Рефлекс - это автоматический ответ организма на внешний раздражитель. Исторически он является одной из наиболее древних реакций нервной системы. Рефлекторный акт - непроизвольный, то есть его невозможно контролировать сознанием.
Последовательность нейронов и их отростков, которые обеспечивают определенный рефлекс, называются рефлекторными дугами. Она необходима для проведения импульса от чувствительного рецептора до нервного окончания в рабочем органе.
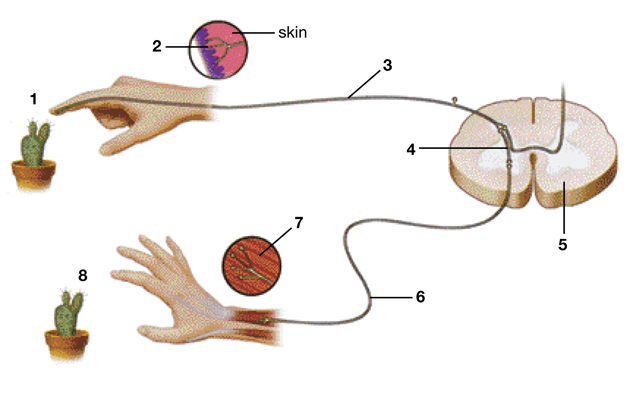
Строение рефлекторной дуги
Рефлекторную дугу двигательного рефлекса называют простейшей, так как она состоит всего лишь из двух нервных клеток или нейронов. Поэтому она также носит название двухнейронной. Проведение импульса обеспечивают следующие отделы рефлекторной дуги:
- Первый нейрон является чувствительным, своим дендритом (коротким отростком) он тянется на периферические ткани, заканчиваясь рецептором. А его длинный отросток (аксон) тянется в другую сторону - к спинному мозгу, заходит в задние рога спинного мозга, а после в передние, образуя соединение (синапс) со следующим нейроном.
- Второй нейрон называется двигательным, его аксон тянется из спинного мозга к скелетным мышцам, обеспечивая их сокращение в ответ на раздражитель. Соединение между нервом и мышечным волокном носит название нервно-мышечного синапса.
Именно благодаря передаче нервного импульса по рефлекторной дуге возможно существование спинальных двигательных рефлексов.
Виды рефлексов
В общем, все рефлексы подразделяют на простые и сложные. Спинальные рефлексы, о которых идет речь в этой статье, относятся к категории простых. Это означает, что для их осуществления достаточно лишь нейронов и спинного мозга. Структуры головного мозга не принимают участия в формировании рефлекса.
Классификация спинальных рефлексов основана на том, какой стимул вызывает данную реакцию, а также в зависимости от функции организма, выполняемой при помощи этого рефлекса. Кроме того, при классификации учитывается, какая часть нервной системы принимает участие в рефлекторном ответе.
Выделяют следующие виды спинальных рефлексов:
- вегетативные - мочеиспускание, потоотделение, сужение и расширение сосудов, дефекация;
- двигательные - сгибательные, разгибательные;
- проприоцептивные - обеспечение ходьбы и поддержание тонуса мышц, возникают при стимуляции мышечных рецепторов.
Двигательные рефлексы: подвиды
В свою очередь, двигательные рефлексы подразделяются на еще два вида:
- Фазные рефлексы обеспечиваются однократным сгибанием или разгибанием мышц.
- Тонические рефлексы возникают при многочисленном последовательном сгибании и разгибании. Они необходимы для поддержания определенной позы.
В неврологии чаще всего применяется другая классификация видов рефлексов. Согласно этому разделению, рефлексы бывают:
- глубокие или проприоцептивные - сухожильные, надкостничные, суставные;
- поверхностные - кожные (проверяются наиболее часто), рефлексы слизистых оболочек.

Методы определения рефлексов
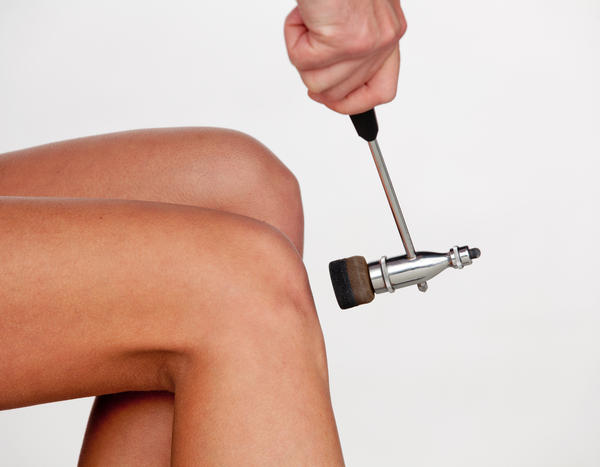
Состояние рефлекса может многое сказать о работе нервной системы. Проверка рефлексов при помощи молоточка - важная часть неврологического осмотра.
Глубокие (проприоцептивные) рефлексы можно определить при помощи легкого постукивания молоточком по сухожилию. В норме должно наблюдаться сокращение соответствующих мышц. Визуально это проявляется разгибанием или сгибанием определенной части конечности.
Кожные рефлексы вызываются быстрым проведением рукояткой неврологического молоточка по специфическим участкам кожи пациента. Эти рефлексы исторически более новые, чем глубокие. Так как они позже образовались, то и при патологии нервной системы именно этот вид рефлексов исчезает первым.
Глубокие рефлексы
Выделяют следующие виды спинальных рефлексов, которые берут свое начало в рецепторе сухожилий:
- Бицепс-рефлекс - возникает при легком ударе по сухожилию двуглавой мышцы плеча, его дуга проходит через IV-VI шейные сегменты спинного мозга (СМ), нормальная реакция - сгибание предплечья.
- Трицепс-рефлекс - происходит при ударе по сухожилию трицепса (трехглавой мышцы), его дуга проходит через VI-VII шейные сегменты СМ, нормальная реакция - разгибание предплечья.
- Пястно-лучевой - вызывается ударом по шиловидному отростку лучевой кости и характеризуется сгибанием кисти, дуга проходит через V-VIII шейные сегменты СМ.
- Коленный - вызывается ударом по сухожилию под надколенником и характеризуется разгибанием ноги. Дуга проходит через II-IV поясничные сегменты спинного мозга.
- Ахиллов - возникает при ударе молоточком по Ахиллову сухожилию, его дуга проходит через I-II крестцовые сегменты спинного мозга, нормальная рефлекторная реакция - подошвенное сгибание стопы.

Кожные рефлексы
Поверхностные, или кожные, рефлексы также важны в неврологической практике. Их механизм подобен глубоким рефлексам: сокращение мышцы, которое возникает при раздражении рецепторных окончаний. Только в данном случае раздражение происходит не при помощи удара молоточка, а штриховым движением рукоятки.
Выделяют следующие виды кожных спинальных рефлексов:
- Брюшные, которые, в свою очередь, подразделяются на верхний, средний и нижний рефлексы. Верхний брюшной рефлекс возникает при раздражении рецепторов участка кожи под реберной дугой, средний - около пупка, нижний - под пупком. Дуги этих рефлексов замыкаются на уровне VIII-IX, X-XI, XI-XII грудных сегментов СМ соответственно.
- Кремастерный - представляет собой подтягивание яичек вверх из-за сокращения его мышц в ответ на раздражение кожного участка внутренней стороны бедра. Дуга рефлекса проходит на уровне I-II поясничных сегментов СМ.
- Подошвенный - сгибание пальцев нижних конечностей при штриховом раздражении кожи подошвы, уровень рефлекса - от V поясничного сегмента до I крестцового.
- Анальный - находится на уровне IV-V крестцовых сегментов и вызывается штриховыми движениями по коже околоанальной области, что приводит к сокращению сфинктера.
Наиболее широкое использование в неврологической практике получило определение брюшных и подошвенного рефлексов.

Патология спинальных рефлексов
В норме рефлексы должны быть оживленными, однофазными (то есть без колебательных движений конечности), умеренной силы. Состояние, когда рефлексы повышенной силы или активности называются гиперрефлексией. Когда же рефлексы, наоборот, снижены, говорят о наличии гипорефлексии. Полное их отсутствие носит название арефлексии.
Гиперрефлексия возникает при повреждении центральной нервной системы. Чаще всего этот патологический симптом возникает при следующих заболеваниях:
- инсульты (ишемический и геморрагический);
- инфекционное воспаление центральной нервной системы (энцефалит, энцефаломиелит);
- церебральный паралич;
- травмы головного и спинного мозга;
- новообразования.
Гипорефлексия, в свою очередь, является одним из проявлений нарушения работы периферической нервной системы. Данное состояние вызывают такие заболевания, как:
- полиомиелит;
- периферические нейропатии (алкогольная, диабетическая).
Однако снижение рефлекторной деятельности нервной системы может возникать и при повреждении центральной нервной системы. Это происходит, когда патологический процесс возникает в том сегменте спинного мозга, где проходит дуга рефлекса. Например, при поражении V шейного сегмента СМ, бицепс-рефлекс будет снижен, в то время как другие глубокие рефлексы, замыкающиеся на более низких сегментах, будут повышены.
Вегетативные рефлексы
Наверное, вегетативные рефлексы являются наиболее сложной разновидностью спинальных рефлексов. Их функцию нельзя определить с помощью обычного неврологического молоточка, однако, именно они обеспечивают жизненно важные функции нашего организма. Их возникновение возможно благодаря функции специфического образования в головном мозге - ретикулярной формации, в которой находятся следующие центры регуляции:
- сосудодвигательный, обеспечивающий деятельность сердца и сосудов;
- дыхательный, который регулирует глубину и частоту дыхания через центры, иннервирующие дыхательные мышцы;
- пищевой, благодаря которому повышается моторная и секреторная функции желудка и кишечника;
- защитные центры, при раздражении которых человек кашляет, чихает, испытывает тошноту и рвоту.
Исследование рефлекторной деятельности нервной системы - важная часть неврологического осмотра пациента, которая позволяет установить локализацию повреждения, что способствует своевременной диагностике.
Все рефлексы спинного мозга делятся на соматические (двигательные) и вегетативные. Соматические рефлексы делятся на сухожильные (миотатические) и кожные. Сухожильные рефлексы возникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению рецепторов растяжения, затем сигналы от альфа-мотонейронов спинного мозга поступают к мышцам, последние сокращаются. Характерны в первую очередь для мышц-разгибателей. В клинике определяют коленный, ахиллов, локтевой, кистевой и др. рефлексы. Коленный рефлекс имеет моносинаптический характер, т.е. в его центральной части имеется один синапс. Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной. Спинальные нервные центры находятся под контролем вышележащих НЦ. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшается.
Вегетативные рефлексы спинного мозга делятся на симпатические и парасимпатические. Те и другие проявляются реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны спинного мозга образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эйякуляции и т.д.
Функции продолговатого мозга
Основными функциями продолговатого мозга являются проводниковая, рефлекторная и ассоциативная. Первая осуществляется проводящими путями, проходящими через него. Вторая – нервными центрами. В ромбовидной ямке продолговатого мозга находятся ядра 10, 11, 12 пар черепномозговых нервов, а также ретикулярная формация. Рефлекторные функции делятся на соматические и вегетативные. Соматическими являются статические рефлексы продолговатого мозга, относящиеся к познотоническим или рефлексам позы. Эти рефлексы осуществляются ядром Дейтерса из группы вестибулярных ядер.От него к мотонейронам разгибателей спинного мозга идут нисходящие вестибулоспинальные тракты. Рефлексы возникают тогда, когда возбуждаются вестибулярные рецепторы или проприорецепторы мышц шеи. Коррекция положения тела происходит за счет изменения тонуса мышц. Например, при запрокидывании головы животного назад повышается тонус разгибателей передних конечностей и снижается тонус разгибателей задних. При наклоне головы вперед возникает обратная реакция.Приповороте головы в сторону, повышается тонус разгибателей конечности на этой стороне и сгибателей противоположной конечности.
В продолговатом мозге находятся жизненно важные центры. К ним относятся дыхательный, сосудодвигательный центры и центр регуляции сердечной деятельности. Первый обеспечивает смену фаз дыхания, второй – тонус периферических сосудов, третий – регуляцию частоты и силы сердечных сокращений.
В области ядер блуждающего нерва находятся центры слюноотделения, секреции желудочных, кишечных желез, поджелудочной железы и печени. Здесь же расположены центры регуляции моторики пищеварительного канала.
Важной функцией продолговатого мозга является формирование защитных рефлексов. В нем находятся рвотный центр, центры кашля, чихания, смыкания век и слезотечения при раздражении роговицы.
Здесь расположены бульбарные отделы центров, участвующих в организации пищевых рефлексов – сосания, жевания, глотания.
В продолговатом мозге происходит первичный анализ ряда сенсорных сигналов. В частности, в нем расположены ядра слухового нерва, верхнее вестибулярное ядро, а к ядрам языкоглоточного нерва поступают сигналы от вкусовых рецепторов. От рецепторов кожи лица они идут к ядрам тройничного нерва.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также:
