
Сила тока в нервах
При определенных условиях в нервах и мышцах возникают электрические токи. Различают: 1) токи, или потенциалы, покоя и повреждения и 2) токи, или потенциал, действия – акционные точки.
Токи покоя
При отсутствии раздражения и, следовательно, возбуждения, существует разность потенциалов между наружной поверхностью цитоплазмы живой клетки (мембраной) и её внутренним содержимым.
Поверхностный слой цитоплазмы живой клетки, находиться в состоянии покоя, имеет положительный электрический заряд, а цитоплазма внутри клетки заряжена электроотрицательно. Поэтому, введя в клетку микроэлектрод и приложив другой электрод к поверхности клетки, можно отвести ток к регистрирующему прибору и измерить потенциал покоя, мембранный или клеточный потенциал.
Токи покоя, или клеточные потенциалы, обнаруживаются в неповрежденных живых клетках. Современными методами отведения электрического тока и его регистрации установлено, что токи покоя отдельных клеток животных, например гигантских нервных волокон кальмара, — 50-60 мв (милливольт), миелиновых нервных волокон амфибий – 70 мв, теплокровных – 90 мв, большинства тел нейронов – 80-100 мв, поперечнополосатых мышц амфибий – 80 мв, эпителия амфибий – 20 мв, волокон сердечной мышцы животных с постоянной температурой тела – 95 мв.
Токи повреждения
При повреждении живой ткани открывается доступ к внутреннему содержимому клеток, которое заряжено электроотрицательно. Поэтому если приложить электрод к неповрежденному участку ткани, находящемуся в состоянии покоя и, следовательно, заряженному электроположительно, а другой электрод приложить к поврежденному или перерожденному участку, заряженному электроотрицательно, то при соединении обоих электродов проводником тока с регистрирующим прибором можно измерить разность потенциалов.
Токи повреждения можно обнаружить на нервно-мышечном препарате. Если разрезать мышцу или любым способом повредить ее в одном участке (прижечь, раздавить и т. д.) и затем, приложив неполяризующиеся электроды, соединить срез мышцы или участок повреждения с целой поверхностью мышцы, не подвергающейся раздражению, то от целой поверхности к участку разреза или повреждения направится электрический ток. Сильно поврежденный участок всегда электроотрицателен по отношению к электроположительному неповрежденному участку. Следовательно, биоэлектрический ток повреждения, или альтерационный ток, возникает на границе целой и поврежденной частей ткани. Он обнаруживается и на некотором расстоянии от места повреждения, уменьшаясь с увеличением расстояния от места повреждения. Поэтому по месту происхождения его называют также демаркационным током. Однако ток повреждения появляется только тогда, когда отрицательно заряженный поврежденный участок соединяется проводником тока с положительно заряженным целым участком.
Величина тока повреждения меньше, чем клеточного потенциала, так как межклеточная и внеклеточная жидкости производят шунтирование (укорочение) при его измерении.
Разность потенциалов между целым и поврежденным участками скелетных мышц лягушки сразу же после повреждения равна 25-50 мв, наибольшая разность доходит до 80. В скелетных мышцах кролика токи повреждения равны 40 мв. В гладких мышцах токи повреждения меньше и быстрее уменьшаются до нуля. Разность потенциалов между целым и поврежденным участками седалищного нерва лягушки достигает 20-30 мв, седалищного нерва лошади — 6-16 мв, а седалищного нерва человека и обезьяны — 6-7 мв. Следовательно, токи повреждения нервов меньше, чем токи повреждения мышц. Особенно малы токи повреждения нервов человека и высших животных. Постепенно разность потенциалов уменьшается.
Когда нерв второго нервно-мышечного препарата набрасывается на первый нервно-мышечный препарат таким образом, чтобы он одновременно коснулся неповрежденного поврежденного участков мышцы, электрическая цепь замыкается. При этом мышца второго препарата сокращается в результате раздражения нерва током, возникающим в момент его замыкания между положительно заряженным нормальным участком мышцы первого препарата и отрицательно заряженным поврежденным ее участком.
Токи действия
Акционные токи, или токи (потенциалы) действия, возникают в раздражаемых тканях. Их появление характеризует возбуждение.
Для обнаружения тока действия нужно приложить один электрод к неповрежденному участку живой ткани, который находится в состоящих покоя и поэтому заряжен электроположительно, а другой — к раздражаемому участку и присоединить оба электрода к регистрирующему прибору. Раздражаемый участок при приложении к нему раздражителя немедленно или через несколько сотых миллисекунды становится электроотрицательным. Это падение потенциала’ доходит до максимума, а затем данный участок снова становится электроположительным. Различают однофазные и двухфазные токи действия.
При прикладывании одного из неполяризующихся электродов к неповрежденному участку нерва или мышцы, а другого — к поврежденному участку возникает электрический ток. Этот ток регистрируется при включении в цепь гальванометра или другого более чувствительною прибора.
Стрелка гальванометра отклонится в одну сторону вследствие электроотрицательного заряда поврежденного участка и электроположительного заряда нормального участка. Но если нанести раздражение по соседству с нормальным участком, то волна возбуждения, которая возникнет в месте раздражения, на некоторое время превратит заряд под нормальным участком в электроотрицательный. В этот момент тока не будет, так как под обоими электродами заряд станет электроотрицательным и стрелка гальванометра станет на нулевую линию. Когда волна возбуждения продвинется дальше и будет находиться между электродами, нормальный участок снова станет электроположительным и стрелка гальванометра снова отклонится в ту же сторону.
Когда волна возбуждения дойдет до поврежденного участка, положение стрелки гальванометра не изменится, так как под вторым электродом в месте повреждения уже имеется электроотрицательный заряд. Таким образом, стрелка гальванометра движется только в одну сторону, а запись этого движения регистрирует однофазный, или монофазный ток действия.
Это явление Дюбуа-Реймон назвал отрицательным колебанием тока покоя, которое возникает каждый раз, когда волна возбуждения проходит под электродом, расположенным на неповрежденном участке ткани.
Однофазный ток действия можно зарегистрировать также при раздражении ткани во время отведения клеточного потенциала. При внутриклеточном введении одного электрода и приложении другого электрода к поверхности клетки установлено, что клеточный потенциал не только исчезает в течение тысячных долей секунды, но что в течение этого промежутка времени амплитуда тока действия на 30-50 мв больше клеточного потенциала. Это объясняется тем, что в пункте прохождения волны возбуждения наружная поверхность мембраны становится электроотрицательной, а внутренняя — электроположительной. Продолжительность тока действия в нервных волокнах и клетках скелетных мышц составляет 0,1-5 мс.
Однофазный ток действия представляет собой местный электроотрицательный потенциал, мгновенно возникающий при очень слабых, подпороговых одиночных раздражениях и ограниченный несколькими миллиметрами от пункта раздражения. Величина местного низковольтного электроотрицательного заряда убывает по мере удаления от пункта раздражения. Чем больше подпороговое раздражение, тем больше возникающее возбуждение и амплитуда или отклонение однофазного тока действия.
Следовательно, местное возбуждение или местный сдвиг обмена веществ отличается градуальностью — возрастанием возбуждения по мере увеличения силы раздражения. Местное возбуждение наступает сразу после раздражения, т. е. без латентного периода. В участке слабого местного нераспространяющегося возбуждения возбудимость сохраняется, рефрактерность отсутствует. Если бы при местном возбуждении наблюдалась рефрактерность, то оно не могло бы перейти в пороговое, распространяющееся возбуждение.
Местный однофазный ток действия предшествует распространяющейся волне возбуждения и подготавливает ее. При одиночном пороговом раздражении волна возбуждения возрастает, достигает критического уровня и превращается из местной в распространяющуюся. Поэтому распространяющееся возбуждение наступает после латентного периода, продолжительность которого равна длительности местного нераспространяющегося возбуждения. В отличие от местного возбуждения распространяющееся возбуждение не градуально и сопровождается фазами изменения возбудимости. Потенциал действия, возникший в раздражаемом участке мембраны, вызывает возбуждение соседних участков мембраны и таким образом возбуждение распространяется по нервным и мышечным волокнам.
О прохождении волны возбуждения по возбудимой ткани судят прежде всего по токам действия. Однофазное колебание тока действия распространяющейся волны возбуждения в отдельных нервных и мышечных клетках состоит из нескольких составляющих: 1) высоковольтного потенциала, пика, 2) отрицательного следового потенциала и 3) положительного следового потенциала.
По сравнению со следовыми потенциалами пик имеет наибольшую высоту и наименьшую продолжительность. Пик имеет приблизительно одинаковую высоту на всем протяжении возбудимых тканей животных с постоянной температурой тела. Это указывает на то, что по мере распространения возбуждения вольтаж потенциала не падает. Этот факт свидетельствует, что химические вещества, необходимые для возникновения возбуждения, равномерно распределены по возбудимой ткани и что распространение возбуждения происходит самостоятельно, автоматически. Раздражение при достаточной интенсивности вызывает возбуждение только в том участке, который раздражается, а дальше возбуждение движется по ткани, вовлекая в сдвиг обмена веществ вещества, имеющиеся в возбудимой ткани.
Пик играет ведущую роль в распространении возбуждения. Высота пика различна у нервных волокон разных групп, имеющих неодинаковый диаметр, и изменяется в зависимости от физиологического состояния волокна и скорости проведения возбуждения. Чем толще нервное миелиновое волокно, тем меньше длительность пика. По мышечному волокну человека возбуждение проходит за 4-5 м/с.
В волокнах скелетных мышц однофазные токи действия различаются по амплитуде и по продолжительности. В волокнах сердечной мышцы они отличаются большей продолжительностью пика, который после начального крутого снижения начинает снижаться постепенно, а затем снова круто падает.
Токи действия в гигантских нервных волокнах кальмара достигают 90-110 мв, в миелиновых нервных волокнах амфибий 110 мв, в волокнах поперечнополосатых мышц амфибий — 120 мв, а в волокнах сердечной мышцы животных с постоянной темпера турой тела — 135 мв. Чем больше частота тока действия, тем меньше его вольтаж. Гладкие мышечные волокна отличаются тем, что их токи действия не превосходят мембранных потенциалов (не больше 70-80 мв).
Условия, улучшающие обмен веществ (повышение темпера туры, отдых), уменьшают продолжительность волны возбуждения, а условия, ухудшающие обмен веществ (утомление), увеличивают ее. С понижением температуры амплитуда пика уменьшается, а продолжительность его возрастает.
При температуре тела продолжительность подъема высоковольтного потенциала нерва составляет примерно 1/3 а продолжительность снижения — 2/3 его общей продолжительности. Предполагается, что крутой подъем пика обусловлен быстрым движением ионов натрия внутрь клетки.
Отрицательный следовой потенциал соответствует остаточным сдвигам распределения ионов калия и натрия. В отличие от пика он неустойчив и изменяется от условий среды. Он достигает 0,05 потенциала пика. Положительный низковольтный следовой потенциал также связан с перемещением ионов и равен примерно 0,002 потенциала пика.
Тепло и отдых уменьшают, а охлаждение и истощение увеличивают продолжительность следовой электроотрицательности. В нерве, лишенном кислорода, при действии наркоза и солей калия следовая низковольтная электроотрицательность отсутствует, а начальная высоковольтная электроотрицательность сохраняется, и при известных концентрациях наркоза даже не наступает заметных изменений её величины. Это указывает на то, что пик может возникать и при отсутствии кислорода, а следовая электроотрицательность связана с восстановительными процессами, которые совершаются в ткани вслед за возбуждением, и для её появления необходим кислород. Нарушение обмена веществ, утомление увеличивают продолжительность следовых потенциалов. Чем больше лабильность, тем меньше выражены следовые потенциалы. В высоколабильных мякотных волокнах человека и высших животных они почти не проявляются.
Двухфазный ток действия
В естественных условиях одиночные волны возбуждения встречаются чрезвычайно редко; В организме в нервах и мышцах передаются серии волн возбуждения, взаимно влияющих друг на друга. Возбуждение, которое достигает порога или несколько превышает его, носит ритмический характер. В других случаях возбуждение в естественных условиях является градуальным, длительным, стойким, переходящим в торможение. В последнем случае при чрезмерно большой силе или частоте раздражения, превышающей меру лабильности, полностью угнетается высоковольтная электрическая активность.
Волновое распространяющееся возбуждение возникает из неволнового, градуального. Такая же эволюция возбуждения происходила в филогенезе. Представление о возникновении волнового возбуждения дает следующий простой опыт.
После приложения обоих неполяризующихся электродов к нормальным целым участкам возбудимой ткани и включения этой ткани в цепь гальванометра его стрелка останется на нуле, так как все участки нормальной ткани, находящейся в состоянии покоя, имеют одинаковый заряд. Но когда ткань раздражают у одного из электродов, то возникает возбуждение, и заряд ткани под этим электродом становится отрицательным. При этом стрелка гальванометра отклоняется в одну сторону. Передвигаясь далее, волна возбуждения оказывается между электродами, и в это время заряды под электродами вновь становятся одинаковыми и стрелка гальванометра возвращается к нулю. Но когда волна возбуждения доходит до второго электрода, то гальванометр опять отмечает разность потенциалов, так как под вторым электродом заряд становится отрицательным, а в то же самое время под первым электродом заряд положительный, так как под ним нег возбуждения. Теперь стрелка гальванометра отклоняется в противоположную сторону, а затем, после прекращения возбуждения, возвращается к нулю. Следовательно, когда по ткани проходит волна возбуждения, то стрелка гальванометра отклоняется сначала в одну сторону, потом в другую. Запись этого движения стрелки гальванометра регистрирует двухфазный ток действия.
Форма регистрации токов, или потенциалов, действия зависит от места возникновения возбуждения, от используемых приборов и от расположения электродов. В настоящее время их записывают посредством катодных осциллографов, не имеющих инерции, в отличие от гальванометров. Современные катодные осциллографы регистрируют биопотенциалы в миллионные доли вольта, продолжающиеся одну стомиллионную или даже миллиардную долю секунды. Эти токи отличаются сложностью, и их нередко расшифровывают при помощи электронных устройств.
Биопотенциалы каждой возбудимой ткани, например сердца, имеют характерный вид и изменяются в зависимости от ее функционального состояния. Записываются биопотенциалы кожи, эпителия, соединительной ткани, рецепторов, сетчатки глаза, скелетных мышц, пищеварительных желез, гладкой мускулатуры пищеварительного канала и других органов, разных отделов центральной нервной системы и даже одного нейрона. Первые исследования биопотенциалов больших полушарий головного мозга провели В. Я. Данилевский, Р. Кетон (1875), продолговатого мозга — И. М. Сеченов (1882).
Н. В. Введенский (1883, 1884) соединил мышцу проводами с телефонной трубкой и установил важнейшие законы, по которым протекает в ней возбуждение. Он же впервые применил телефон для изучения явлений возбуждения в нерве.

Мышечное напряжение
Прежде всего, надо помнить, что перенапряжение бывает физическим и эмоциональным. И, когда мы говорим о первом, нельзя не учитывать и второе. Что касается физического перенапряжения, то если ресурсы тела человека позволяют его компенсировать, все в порядке. Это нормально: мы постоянно так или иначе испытываем напряжение.
Но, если оно было сильным и длительным и превысило возможности организма, это может привести к формированию нежелательных изменений в мышцах, что требует вмешательства специалиста: остеопата или массажиста. Когда изменения накапливаются и организм не справляется с ними, человек в течение длительного времени может быть вялым, ощущать отсутствие энергии, что сказывается на его качестве жизни, на продуктивности. Кроме того, в разных частях тела могут появиться боли.

Как бы устрашающе это ни звучало, но факт остается фактом: 95% детей получают родовую травму. Не устраненные в детстве последствия создают дополнительную зону риска, в частности, для перенапряжений, возникновения спазмов и т. д. И, если это происходит часто, необходимо обращаться к специалисту и устранять последствия травм.
Чем чревато
О последствиях мышечных зажимов мало кто думает. При этом они могут быть довольно серьезными. Итак, в первую очередь напряжение в теле, присутствующее постоянно, становится причиной головных болей. Спазмы пережимают сосуды и нервные окончания, нарушается кровоснабжение органов и ухудшается работа нервной системы. Наиболее часто возникают спазмы мышц шеи и плеч: это характерно для сидячих видов работы. На фоне такого нарушения развиваются головные боли, мигрени и даже ухудшается зрение.
Естественно, из-за постоянного тонуса в мышцах развиваются проблемы с осанкой, ведь спазмы в мышцах нередко даже меняют геометрию скелета.
Еще одним следствием постоянного напряжения в теле становится сбой в работе внутренних органов. Ведь развиваются проблемы с лимфотоком, кровообращением, неправильно идет дыхание (оно становится укороченным), нарушается работа пищеварительной системы и т. д.
И, конечно же, жесткие напряженные мышцы не могут хорошо тянуться. Соответственно, человек становится негибким, ему сложнее выполнять некоторые простые манипуляции и т. д.

Правильные методы
Расслабляться надо уметь! Конечно, сон — это решение многих вопросов, однако не всех! Спазм, перенапряжение просто так не пройдут. Поэтому вам придется приложить некоторые усилия, выполнить определенные действия, чтобы помочь организму как следует расслабиться.
1. Тепло
Да, это работает. Причем согревать надо организм полностью, а особенно — ту часть тела, которая перенапряжена, болит, где присутствует спазм. Самый простой вариант — горячая ванна. Только принимать ее надо правильно:
- Вода должна быть горячей;
- Покинув ванну, нужно вытереть тело, надеть сухую одежду и продолжить процедуру под одеялом;
- Не попадать под сквозняк, не выходить сразу на улицу;
- Пить горячие напитки, например, чай.
Однократное принятие ванны не даст ожидаемого результата, поэтому повторяйте прогревание в течение нескольких дней.

2. Баня или сауна
3. Прогулки
Длительная ходьба — это умеренная и полезная физическая нагрузка, которая показана всем, особенно людям, чье тело перенапряжено и находится в спазме.
4. Легкие упражнения, растяжка
Забудьте на время о тяжелых упражнениях. А вот легкие надо делать постоянно и часто: это не просто безопасно, это полезно.
5. Самомассаж, легкий массаж

Вспоминаем бабушек: шерстяные носки, теплый шарф, пуховый платок, шерстяной пояс, разогретые мешочки с солью или крупой и т. д. — это отлично работает во все времена, помогая снимать напряжение.
7. Ароматерапия
Масло лаванды прекрасно помогает в борьбе с перенапряжением. Мазь, кремы с лавандой, вдыхание аромамасла лаванды — для снятия мышечных болей хороши любые средства. Можно добавить несколько капель масла лаванды в ванну.
Запомните эти простые и эффективные способы снятия перенапряжения. Не думайте, что таблетки и уколы помогут вам быстрее. Когда речь идет о спазмах и перенапряжении, у вас есть два пути: помочь себе в домашних условиях практически народными рецептами и обратиться к массажисту или к остеопату.
Законы раздражения возбудимости тканей
Закон силы
Возникновение распространяющегося возбуждения (ПД) возможно при условии, когда действующий на клетку раздражитель имеет некоторую минимальную (пороговую силу), иначе говоря, когда сила раздражителя соответствует порогу раздражения.
Порог раздражения – это та наименьшая величина раздражителя, которая действуя на клетку какое-то определенное время, способна вызвать максимальное возбуждение.
– наименьшая величина раздражителя, при действии которой потенциал покоя может сместиться до уровня критической деполяризации;
– критическая величина деполяризации клеточной мембраны, при которой активируется перенос ионов натрия внутрь клетки.

Рисунок 5. Возникновение местного потенциала при прохождении электрического тока, через участок нерва. Ток идет от анода к катоду (оба электрода – вне нерва) частично через пленку жидкости на поверхности нерва, а частично через оболочку нерва и в продольном направлении внутри волокна. Кривая внизу показывает вызываемое током изменения мембранного потенциала нервного волокна (по В. Катц)
Зависимость пороговой силы стимула от его длительности
Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Эта зависимость, открытая Гоорвегом, Вейсом, Лапиком получила название кривой “сила – длительность” или “сила – время”. Кривая “силы времени” имеет форму, близкую к равносторонней гиперболе и в первом приближении может быть описана эмпирической формулой:
где I – сила тока, Т – длительность его действия, а и b – постоянные, определяемые свойствами ткани.
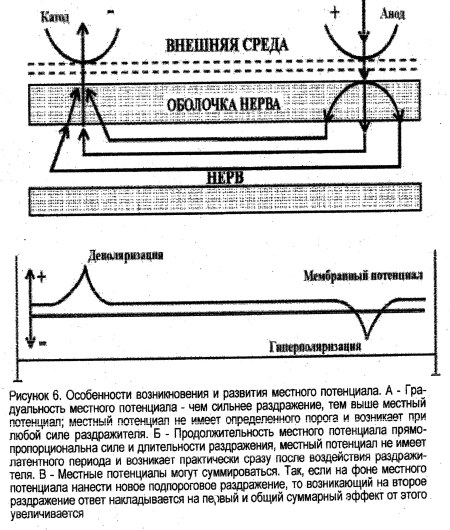
Рисунок 6. Особенности возникновения и развития местного потенциала. А – Градуальность местного потенциала – чем сильнее раздражение, тем выше местный потенциал; местный потенциал не имеет определенного порога и возникает при любой силе раздражителя. Б – Продолжительность местного потенциала прямопропорциональна силе и длительности раздражения, местный потенциал не имеет латентного периода и возникает практически сразу после воздействия раздражителя. В – Местные потенциалы могут суммироваться. Так, если на фоне местного потенциала нанести новое подпороговое раздражение, то возникающий на второе раздражение ответ накладывается на первый и общий суммарный эффект от этого увеличивается
Таким образом, го этой кривой вытекают два следствия:
1. Ток величиной ниже порога не вызывает возбуждения, как бы длительно он не действовал.
2. Какой бы сильный не был раздражитель, но если он действует очень короткое время, то возбуждение не возникает.
Реобаза – минимальная сила тока (или напряжение), способная вызвать возбуждение. Наименьшее время, в течение которого должен действовать стимул в одну реобазу, чтобы вызвать возбуждение – полезное время. Дальнейшее его увеличение не имеет значения для возникновения возбуждения.
Порог (реобаза) – величины непостоянные, зависят от функционального состояния клеток в покое, поэтому Лапик предложил определять более точный показатель – хронаксию.
Хронаксия – наименьшее время, в течение которого ток в две реобазы должен действовать на ткань, чтобы вызывать возбуждение.
Метод определения хронаксии – хронаксиметрия используется в клинике для диагностики повреждения нервных стволов и мышц.
Зависимость порога от крутизны нарастания раздражителя (аккомодация)
Порог раздражения имеет наименьшую величину при толчках электрического тока прямоугольной формы, когда сила нарастает очень быстро.
При уменьшении крутизны нарастания стимула ускоряются процессы инактивации натриевой проницаемости, приводящие к повышению порога и снижению амплитуды потенциалов действия. Чем круче должен нарастать ток, чтобы вызвать возбуждение, тем выше скорость аккомодации. Очень мала скорость аккомодации тех образований, которые склонны к автоматической деятельности (миокард, гладкие мышцы).
“Все” — на пороговые и сверхпороговые стимулы ответ максимальный и развивается потенциал действия; “ничего” – надо – пороговый стимул потенциал действия не развивается. Закон “все или ничего” установлен Боудичем в 1871 г. на мышце сердца: при подпороговой силе раздражения мышца сердца не сокращается, а при пороговой силе раздражения – сокращение максимально. При дальнейшем увеличении силы раздражения амплитуда сокращений не увеличивается.
Со временем была установлена и относительность этого закона. Оказалось, что “все” зависит от функционального состояния ткани (охлаждение, исходное растяжение мышцы и т. д.). С появлением микроэлектродной техники было установлено еще одно несоответствие: подпороговое раздражение вызывает местное, не распространяющееся возбуждение, следовательно, нельзя говорить, что допороговое раздражение не дает ничего. Процесс развития возбуждения подчиняется этому закону с уровня критической деполяризации, когда запускается лавинообразное поступление ионов калия в клетку.
Изменение возбудимости при возбуждении
Мера возбудимости – это порог раздражения. При местном, локальном, возбуждений возбудимость увеличивается. Потенциал действия сопровождается многофазными изменениями возбудимости.
1. Период повышенной возбудимости соответствует локальному ответу, когда мембранный потенциал достигает УКД, возбудимость повышена.
2. Период абсолютной рефрактерности соответствует фазе деполяризации потенциала действия, пику и началу фазы реполяризации, возбудимость снижена вплоть до полного отсутствия во время пика.
3. Период относительной рефрактерности соответствует оставшейся части фазы реполяризации, возбудимость постепенно восстанавливается к исходному уровню.
4. Супернормальный период соответствует фазе следовой деполяризации потенциала действия (отрицательный следовый потенциал), возбудимость повышена.
5. Субнормальный период соответствует фазе следовой гиперполяризации потенциала действия (положительный следовый потенциал), возбудимость снижена.

Рисунок 7. Изменения возбудимости нервного волокна в различные фазы потенциала действия и следовых изменений потенциала действия (по Б.И. Ходорову). Для наглядности длительность первых двух фаз на каждой кривой несколько увеличена. Пунктирной линией на рисунке А обозначает потенциал покоя, а на рисунке Б исходный уровень возбудимости
Закон лабильности или функциональной подвижности
Лабильность — скорость протекания физиологических процессов в возбудимой ткани. Например, можно творить о максимальной частоте раздражения, которую возбудимая ткань способна воспроизводить без трансформации ритма.
Мерой лабильности могут служить:
– длительность отдельного потенциала
– величина абсолютной рефрактерной фазы
– скорость восходящей и нисходящей фаз ПД.
Уровень лабильности характеризует скорость возникновения и компенсации возбуждения в любых клетках и уровень их функционального состояния. Можно измерять лабильность мембран, клеток, органов, причем, в системе из нескольких элементов (тканей, органов, образований) лабильность определяется по участку с наименьшей лабильностью:
Полярный закон раздражения (закон Пфлюгера)
Закон изменения мембранного потенциала при действии на возбудимые ткани постоянного электрического тока открыл Пфлюгер в 1859 г.
1. Постоянный ток проявляет свое раздражающее действие только в момент замыкания и размыкания цепи.
2. При замыкании цепи постоянного тока возбуждение возникает под катодом; при размыкании по анодом.
Изменение возбудимости под катодом.
При замыкании цепи постоянного тока под катодом (действуют допороговым, но продолжительным раздражителем) на мембране возникает стойкая длительная деполяризация, которая не связана с изменением ионной проницаемости мембраны, а обусловлена перераспределением ионов снаружи (они привносятся на электроде) и внутри – катион перемещается к катоду.
Вместе со смещением мембранного потенциала уровень критической деполяризации (УКД) смещается к нулю. При размыкании цепи постоянного тока под катодом мембранный потенциал быстро возвращается к исходному уровню, а УКД медленно, следовательно, порог увеличивается, возбудимость снижается и отмечается католическая депрессия Вериго. Таким образом, возбуждение возникает только при замыкании цепи постоянного тока под катодом.
Изменение возбудимости под анодом.
При замыкании цепи постоянного тока под анодом (действуют допороговым, но продолжительным раздражителем) на мембране развивается гиперполяризация за счет перераспределения ионов по обе стороны мембраны (без изменения ионной проницаемости мембраны) и возникающее за ней смещение уровня критической деполяризации в сторону мембранного потенциала. Следовательно, порог уменьшается, возбудимость повышается – анодическая экзальтация.
При размыкании цепи мембранный потенциал быстро восстанавливается к исходному уровню и достигает сниженного уровня критической деполяризации, генерируется потенциал действия. Таким образом, возбуждение возникает только при размыкании цепи постоянного тока под анодом. Сдвиги мембранного потенциала вблизи полюсов постоянного тока получили название электротонических. Сдвиги мембранного потенциала не связанные с изменением ионной проницаемости мембраны клетки называют пассивными.
Проведение возбуждения.
Потенциал действия – это волна возбуждения, распространяющаяся по мембранам нервных и мышечных клеток.
ПД обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам. Синоним ПД – нервный импульс или спайк. Сложная информация о действующих на организм раздражениях кодируется в виде отдельных групп потенциалов действия – рядов.
Согласно закону “все или ничего” амплитуда и длительность отдельных потенциалов действия постоянны, а частота и количество в ряду зависит от интенсивности раздражения. Такой способ кодирования информации и ее передачи является наиболее психоустойчивым.
В живых организмах информация может передаваться и гуморальным путем.
Преимущества ПД:
1. Информация более целенаправлена;
2. Передается быстро;
3. Адресат точно известен;
4. Информация может быть точнее закодирована.
ПД распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками. Из-за перезарядки мембраны во время генерации потенциала действия последний обладает способностью к самораспространению. Возникнув на одном участке, является стимулом для соседних. Наступающая после возбуждения в данном участке мембраны рефрактерностъ, обусловливает поступательное движение ПД.
Конкретные особенности распространения возбуждения связаны со строением мембраны клетки, нервных волокон. По мембранам мышечных клеток и в безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны.
В волокнах, покрытых миелиновой оболочкой, потенциал действия может распространяться только скачкообразно (сальтаторно), перепрыгивая через участки волокна, покрытые шванновскими клетками, с одного перехвата Ранвье на другой.
Перехваты Ранвье представляют собой своеобразные станции ретрансляции, постоянно усиливающие сигнал, не давая ему угаснуть.
Причины сальтаторного проведения:
1. В перехватах Ранвье, свободных от миелина, сопротивление электрическому току минимально;
2. Порог раздражения в перехватах Ранвье минимальный;
3. Амплитуда ПД в каждом перехвате в 5 – 6 раз превышает порог в соседнем перехвате;
4. Велика плотность натриевых каналов на мембране перехвата.
Следовательно, возбуждение, возникающее в одном перехвате Ранвье, вызывает смещение электронов во внешней среде данного волокна и этого смещения достаточно, чтобы вызвать возбуждение в соседнем участке. Таким образом, скорость проведения возбуждения по нервному волокну зависит от диаметра волокон и наличия перехватов Ранвье.
Различают декрементное и бездекрементное распространение волны возбуждения.
ДЕКРЕМЕНТНОЕ проведение:
1. Наблюдается в безмиелиновых волокнах;
2. Скорость проведения невелика;
3. По мере удаления от места возникновения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания;
4. Свойственно волокнам, которые иннервируют внутренние органы, обладающие низкой функциональной активностью.
БЕЗДЕКРЕМЕНТНОЕ проведение:
1. ПД проходит весь путь от места раздражения до места реализации без затухания.
2. Характерно для миелиновых и тех безмиелиновых волокон, которые передают сигналы к органам, обладающими высокой реактивностью (сердце).
Распространение одиночного потенциала действия само по себе не требует энергетических затрат. Однако, восстановление исходного состояния мембраны и поддержание ее готовности к проведению нового импульса связано с затратой энергии.
Законы проведения возбуждения в нервах
Закон анатомической и физиологической непрерывности волокна.
Любая травма волокна нарушает проводимость. При действии новокаина (дикаина, кокаина) блокируются натриевые и калиевые каналы мембраны. Возникновение возбуждения и его проведение в этом случае становится невозможным.
Закон двустороннего проведения возбуждения
В целом организме по рефлекторной дуге возбуждение всегда распространяется в одном направлении: от рецептора к эффектору.
Причины:
1. Возбуждение всегда возникает при раздражении специфических рецепторов;
2. Рефрактерность во время возбуждения обусловливает поступательное движение;
3. В рефлекторной дуге возбуждение с одной нервной клетки на другую передастся в синапсах с помощью медиатора, который может выделяться только в одном направлении.
Закон изолированного проведения возбуждения в нервных стволах.
Передача возбуждения на большие расстояния невозможна из-за значительной потери тока во внеклеточной среде.
Физиология нейронов, глиальных клеток, рецепторов и синапсов
Классическая рефлекторная дуга состоит из:
– рецептора;
– афферентного пути (афферентного нейрона, который располагается в спинальном ганглии);
– нервного центра, где возбуждение с афферентного нейрона переходит на вставочную нервную клетку.
Затем возбуждение переходит на эффекторный орган (эффектор), в роли которого может выступать мышца. Многие нервные волокна покрыты глиальными клетками (миелиновая оболочка). Между этими Шванновскими клетками есть промежутки – перехваты Ранвье. Возбуждение с одного нейрона на другой и с мотонейрона на мышцу передается в синапсах с помощью медиатора.
Нервная клетка – структурная и функциональная единица ДНС, которая окружена клетками нейроглии.
Нейроглия (глиоциты) – совокупность всех клеточных элементов нервной ткани кроме нейронов.
В мозге взрослого человека 1150 – 200 млрд. глиальных клеток, что в 10 раз больше нервных.
Нейроглия:
1. макроглия:
– астроциты;
– олигодендроциты;
– эиендимоциты.
2. микроглия: глиальные макрофаги.
Астроциты составляют 45 – 60% серого вещества мозга. Покрывают 85% поверхности капилляров мозга (сосудистые ножки астроцитов), крупные отростки астроцитов контактируют с телами нейронов. Основная функция – трофическая.
Олигодендроциты образуют миелин в нервной системе к поддерживают его целостность.
Эпендимоциты – клетки, выстилающие стенки спинномозгового канала и всех желудочков головного мозга. Это граница между спинномозговой жидкостью (ликвор) и тканью мозга.
Функции нейроглии:
1. Опорная – вместе с сосудами и мозговыми оболочками образуют строму ткани мозга.
2. Трофическая – обеспечивают метаболизм нервных клеток (связь с кровеносными сосудами). В глиоцитах сосредоточен весь гликоген ЦНС.
3. Участие в интегративной деятельности мозга:
– формирование следов воздействия (память), а значит и условного рефлекса;
– без глиоцитов (блокада антиглиальным гамма-глобулином) меняется электрическая активность нейронов.
Особенности глиальных клеток:
1. Более чувствительны к ионным изменениям среды;
2. Высокая активность калий – натриевой АТФ-азы;
3. Высокая проницаемость для ионов калия;
4. Мембранный потенциал равен 90 мВ, у нейронов 60 – 80 мВ;
5. На раздражение отвечает только медленной деполяризацией не более 10 мВ;
6. Потенциал действия в глиальных клетках не генерируется.
Читайте также:
