
Скорость проведения нервных импульсов по миелиновым нервным волокнам
Нервы специализируются на проведении возбуждения и связывают нервные центры с исполнительными органами. Нервные волокна формируют нерв или нервный ствол, состоящий из нервных волокон, заключенных в общую соединительнотканную оболочку. Поверхность осевого цилиндра нервного волокна покрыта плазматической мембраной, которая выполняет главную роль в возникновении и проведении возбуждения.
Классификация нервных волокон.
1. По строению нервы делятся на:
Ø миелинизированные (мякотные)
Ø немиелинизированные.
Миелиновые волокна имеют прерывистую оболочку, которая образована сегментами миелина длиной 1—2 мм. Участок между двумя сегментами называется перехватом Ранвье. На поверхности миелин покрыт Швановскими клетками. Миелин обладает высоким сопротивлением и поэтому выполняет изоляционную функцию и кроме того он принимает участие в обмене веществ нервного волокна. Возбуждение по миелиновому волокну распространяется только через перехваты Ранвье так как там много натриевых каналов.
Безмиелиновые волокна обладают такой же структурой, но не имеют миелина. их поверхность покрыта Швановской клеткой.
Если перерезать нервное волокно, то его периферический конец через некоторое время теряет способность проводить возбуждение и дегенерирует. Миелин претерпевает жировое перерождение и превращается в жировые капли. Центральный конец нервного волокна способен регенерировать. На нем образуется колба роста, которая растет к периферии (в сутки от 0,4 до 4,5 мм) и достигает соответствующего органа или ткани. Таким образом, восстанавливается их иннервация. Так, первые признаки восстановления иннервации мышц могут появляться через 5—6 недель.
2. По направлению передачи информации (центр - периферия) нервы подразделяются на:
Ø афферентные
Ø эфферентные.
Эфферентные по физиологическому эффекту делятся на:
Ø Двигательные (иннервируют мышцы).
Ø Сосудодвигательные (иннервируют сосуды).
Ø Секреторные (иннервируют железы).
3. По характеру влияния на эффекторный орган нейроны делятся на:
Ø пусковые (переводят ткань из состояния физиологического покоя в состояние активности)
Ø корригирующие (изменяют активность функционирующего органа).
4. По функциональным свойствам (строению, диаметру волокна, электровозбудимости, скорости развития потенциала действия, длительности различных фаз потенциала действия, по скорости проведения возбуждения) нервные волокна разделили на группы А, В и С.
Группа А неоднородна. Волокна этого типа в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта.
А-альфа – самые толстые волокна покрыты миелиновой оболочкой. Их диаметр от 12 до 22 мкм. Скорость распространения возбуждения по такому волокну составляет от 70 до 120 мс, ПД самый короткий. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от проприорецепторов мышц к соответствующим нервным центрам.
Три другие группы волокон типа А (бета, гамма, дельта) имеют меньший диаметр от 8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна этих групп относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам.
А-бета – диаметр от 8 до 13 мкм. Скорость распространения возбуждения от 40 до 70 мс.
А-гамма – диаметр от 4 до 8 мкм. Скорость распространения возбуждения от 15 до 40 мс.
А-дельта – диаметр от 1 до 4 мкм. Скорость распространения возбуждения от 5 до 15 мс.
К волокнам типа В относятся миелинизированные преганглионарные волокна вегетативной нервной системы. Их диаметр — 1- 3 мкм, а скорость проведения возбуждения — 3-18 м/с, ПД длиться 1—2 мс. У эти волокон отсутствует период супернормальности.
К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра — 0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более от 0,5 до 3 м/с, ПД длиться 2 мс. Большинство волокон типа С — это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
Таким образом, скорость проведения возбуждения по нерву пропорциональна его диаметру.
Нервы практически не утомляются. Они способны очень длительное время проводить импульсы.
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
Состоят из отростка нервной клетки, покрытого оболочкой, которая формируется олигодендроцитами. Отросток нервной клетки (аксон или дендрит) в составе нервного волокна называется осевым цилиндром.
- безмиелиновое (безмякотное) нервное волокно,
- миелиновое (мякотное) нервное волокно.
Безмиелиновые нервные волокна
Находятся преимущественно в составе вегетативной нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи, в которых на определенном расстоянии друг от друга видны овальные ядра. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько (10—20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в смежное. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж неиролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне
29. Миелиновые нервные волокна
1) внутренний, более толстый, — миелиновый слой,
2) наружный, тонкий, состоящий из цитоплазмы, ядер нейролеммоцитов и нейролеммы.
Миелиновый слой содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии — насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы видны участки волокна, лишенные миелинового слоя, — узловатые перехваты, или перехваты Ранвье, т.е. границы между соседними леммоцитами.
Отрезок волокна между смежными перехватами называется межузловым сегментом.
В процессе развития аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита — мезаксон. Мезаксон удлиняется, концентрически наслаивается на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. Цитоплазма с ядрами отодвигается на периферию – образуется наружная оболочка или светлая Шванновская оболочка (при окраске осмиевой кислотой).
Осевой цилиндр состоит из нейроплазмы, продольных параллельных нейрофиламентов, митохондрий. С поверхности покрыт мембраной – аксолеммой, обеспечивающей проведение нервного импульса. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Нервный импульс в миелиновом нервном волокне проводится как волна деполяризации цитолеммы осевого цилиндра, "прыгающая" (сальтирующая) от перехвата к следующему перехвату со скоростью до 120 м/сек.
В случае повреждения только отростка нейроцита регенерация возможна и протекает успешно при наличии определенных для этого условий. При этом, дистальнее места повреждения осевой цилиндр нервного волокна подвергается деструкции и рассасывается, но леммоциты при этом остаются жизнеспособными. Свободный конец осевого цилиндра выше места повреждения утолщается - образуется "колба роста", и начинает расти со скоростью 1 мм/день вдоль оставшихся в живых леммоцитов поврежденного нервного волокна, т.е. эти леммоциты играют роль "проводника" для растущего осевого цилиндра. При благоприятных условиях растущий осевой цилиндр достигает бывшего рецепторного или эффекторного концевого аппарата и формирует новый концевой аппарат.
30. шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
Каждое периферическое нервное волокно одето тонким цитоплазматическим слоем — невролеммой или шванновской оболочкой. Волокно является миелинизированным, если между ним и цитоплазмой шванновской клетки имеется значительный слой миелина. Если волокна лишены миелина, то они называются немиелинизированными безмякотными. Шванновские клетки могут осуществлять волнообразные движения, что, вероятно, способствует транспортировке различных веществ по отросткам нервных клеток.
С нарушением работы шванновских клеток связаны такие нервные заболевания, как синдром Гийена — Барре, болезнь Шарко-Мари, шванноматозис и хроническая воспалительная демиелинизирующая полинейропатия. Демиелинизация в основном происходит из-за ослабления двигательных функций шванновских клеток, в результате чего они оказываются не способны образовывать миелиновую оболочку.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Однако низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. Поэтому природой был выработан еще один дополнительный механизм распространения возбуждения.
32. Механизмы проведения возбуждения в миелиновых волокнах. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированномволокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением.
При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа + , которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности.
Особенности распространения возбуждения по миелиновым волокнам:
2. Возбуждение распространяется с большой скоростью.
3. Возбуждение распространяется без декримента.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
Сальтаторное проведение
(лат. saltatorius, от salto — скачу, прыгаю)
Сальтаторное распространение возбуждения в миелинизированном нервном волокне от перехвата к перехвату [стрелками показано направление тока, возникающего между возбуждённым (А) и соседним покоящимся (Б) перехватом].
34. Проведение нервного импульса, передача сигнала в виде волны возбуждения в пределах одного нейрона и от одной клетки к другой. П. н. и. по нервным проводникам происходит с помощью электротонических потенциалов и потенциалов действия, которые распространяются вдоль волокна в обоих направлениях, не переходя на соседние волокна (см. Биоэлектрические потенциалы, Импульс нервный). Передача межклеточных сигналов осуществляется через синапсы чаще всего с помощью медиаторов, вызывающих появление потенциалов постсинаптических. Нервные проводники можно рассматривать как кабели, обладающие относительно низким осевым сопротивлением (сопротивление аксоплазмы — ri) и более высоким сопротивлением оболочки (сопротивление мембраны — rm). Нервный импульс распространяетсявдоль нервного проводника посредством прохождения тока между покоящимися и активными участками нерва (локальные токи). В проводнике по мере увеличения расстояния от места возникновения возбуждения происходит постепенное, а в случае однородной структуры проводника экспоненциальное затухание импульса, который в 2,7 раза уменьшается на расстоянии l = (константа длины). Так как rm и riнаходятся в обратном отношении к диаметру проводника, то затухание нервного импульса в тонких волокнах происходит раньше, чем в толстых. Несовершенство кабельных свойств нервных проводников восполняется тем, что они обладают возбудимостью. Основное условие возбуждения — наличие у нервов потенциала покоя. Если локальный ток через покоящийся участок вызовет деполяризацию мембраны, достигающую критического уровня (порога), это приведёт к возникновению распространяющегося потенциала действия (ПД). Соотношение уровня пороговой деполяризации и амплитуды ПД, обычно составляющее не менее 1: 5, обеспечивает высокую надёжность проведения: участки проводника, обладающие способностью генерировать ПД, могут отстоять друг от друга на таком расстоянии, преодолевая которое нервный импульс снижает свою амплитуду почти в 5 раз. Этот ослабленный сигнал будет снова усилен до стандартного уровня (амплитуда ПД) и сможет продолжить свой путь по нерву.
Безмиелиновое волокно Миелиновое волокно
Несколько осевых цилиндров 1 осевой цилиндр
Осевые цилиндры — аксоны Осевыми цилиндрами могут быть те, и др . Осевые цилиндры толще, чем в безмиелиновых волокнах
Ядро олигодендроцита – в центре Ядро и цитоплазма олигодендроцита – на периферии волокна
Мезаксоны короткие Мезаксон многократно закручен вокруг осевого циландра, образуется миелиновая оболочка
Na- каналы по всей длине осевого цилиндра Na- каналы только в перехватах Ранвье
Строение периферического нерва.
Нерв состоит из миелиновых и безмиелиновых волокон, сгруппированных в пучки. Он содержит и афферентные, и эфферентные волокна.
Механизмы проведения нервного импульса.
Синапсы — это специальные межклеточные соединения, используемые для перехода сигнала из одной клетки в другую.
Контактирующие участки нейронов очень тесно прилегают друг к другу. Но все же между ними зачастую остается разделяющая их синаптическая щель. Ширина синаптической щели составляет порядка нескольких десятков нанометров.
Чтобы нейтроны успешно функционировали, необходимо обеспечить их обособленность друг от друга, а взаимодействие между ними обеспечивают синапсы.
Синапсы выполняют функцию усилителей нервных сигналов на пути их следования. Эффект достигается тем, что один относительно маломощный электрический импульс освобождает сотни тысяч молекул медиатора, заключенных до того во многих синаптических пузырьках. Залп молекул медиатора синхронно действует на небольшой участок управляемого нейрона, где сосредоточены постсинаптические рецепторы — специализированные белки, которые преобразуют сигнал теперь уже из химической формы в электрическую.
В настоящее время хорошо известны основные этапы процесса освобождения медиатора. Нервный импульс, т. е. электрический сигнал, возникает в нейроне, распространяется по его отросткам и достигает нервных окончаний. Его преобразование в химическую форму начинается с открывания в пресинаптической мембране кальциевых ионных каналов, состояние которых управляется электрическим полем мембраны. Теперь роль носителей сигнала берут на себя ионы кальция. Они входят через открывшиеся каналы внутрь нервного окончания. Резко возросшая на короткое время примембранная концентрация ионов кальция активизирует молекулярную машину освобождения медиатора: синаптические пузырьки направляются к местам их последующего слияния с наружной мембраной и, наконец, выбрасывают свое содержимое в пространство синаптической щели.
Синаптическая передача осуществляется последовательностью двух пространственно разобщенных процессов: пресинаптического по одну сторону синаптической щели и постсинаптического по другую (рис. 3). Окончания отростков управляющего нейрона, повинуясь пришедшим в них электрическим сигналам, высвобождают в пространство синаптической щели специальное вещество-посредник (медиатор). Молекулы медиатора достаточно быстро диффундируют через синаптическую щель и возбуждают в управляемой клетке (другом нейроне, мышечном волокне, некоторых клетках внутренних органов) ответный электрический сигнал. В роли медиатора выступает около десятка различных низкомолекулярных веществ:
ацетилхолин (эфир аминоспирта холина и уксусной кислоты);глутамат (анион глутаминовой кислоты);ГАМК (гамма-аминомасляная кислота);серотонин (производное аминокислоты триптофана);аденозин и др.
Они предварительно синтезируются пресинаптическим нейроном из доступного и относительно дешевого сырья и хранятся вплоть до использования в синаптических пузырьках, где, словно в контейнерах, заключены одинаковые порции медиатора (по несколько тысяч молекул в одном пузырьке).
Схема синапса
Вверху — участок нервного окончания, ограниченный пресинаптической мембраной, в которую встроены пресинаптические рецепторы; синаптические пузырьки внутри нервного окончания наполнены медиатором и находятся в разной степени готовности к его освобождению; мембраны пузырьков и пресинаптическая мембрана содержат пресинаптические белки. Внизу — участок управляемой клетки, в постсинаптическую мембрану которой встроены постсинаптические рецепторы
Синапсы — удобный объект регулирования потоков информации. Уровень усиления сигнала при его передаче через синапс можно легко увеличить или уменьшить, изменяя количество освобождаемого медиатора, вплоть до полного запрета на передачу информации. Теоретически это можно осуществить путем направленного воздействия на любой из этапов высвобождения медиатора.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Взаимодействия между глиальными и нервными клетками отчетливо проявляются в процессах развития и структурной организации нервных волокон. Нервным волокном называется отросток нервной клетки, окруженный глиальной оболочкой.
Непосредственно сам отросток называют еще осевым цилиндром, а клетки глиальной оболочки — нейролеммоцитами. Различают миелиновые (мякотные) и безмиелиновые (безмякотные) нервные волокна.
В безмиелиновых нервных волокнах отростки нервных клеток погружены в углубления на поверхности нейролеммоцитов, имеющих вид желоба. Погруженный в тело глиальной клетки нервный отросток ограничен как собственной плазмолеммой, так и внешней мембраной нейролеммоцита. Он как бы подвешен на двухлистковой ее складке. Эти складки мембран (своеобразные ультраструктурные "брыжейки") называют мезаксонами. Безмиелиновые волокна могут включать несколько осевых цилиндров.
Миелиновое нервное волокно состоит из нервного отростка и нейролеммоцитов (шванновских клеток). Осевой цилиндр не просто погружен в цитоплазму нейролеммоцита, а окружен спиральной слоистой оболочкой (миелином), образованной наматыванием мезаксонов нейролеммоцитов при их вращении вокруг отростка нервной клетки. В миелиновой оболочке обнаружены липиды, щелочной белок миелина, маркерный белок S100 и др.
Высокое содержание липидов (почти 2/3 массы миелина) выявляется при обработке препаратов четырехокисью осмия, окрашивающей миелиновую оболочку в темно-коричневый цвет. По ходу миелинового волокна имеются сужения — узловые перехваты (перехваты Ранвье). Они соответствуют границе смежных нейролеммоцитов. Каждый межузловой сегмент оболочки волокна представлен одним нейролеммоцптом. Миелиновые волокна толще безмиелиновых. Скорость проведения нервного импульса по ним составляет 5-120 м/с, тогда как по безмиелиновым волокнам импульс проводится со скоростью 1-2 м/с.
Сложные взаимоотношения между нервными и глиальными клетками складываются при формировании чувствительных нервных окончаний (рецепторов) и двигательных нервных окончаний (эффекторов).
Нервные окончания — концевой аппарат нервных волокон, формирует межнейрональные контакты, или синапсы, рецепторные (чувствительные) окончания и двигательные (эффекторные) окончания.
Синапс (от synapsis — соединение) — специализированный для передачи нервных импульсов контакт между двумя нейронами или между нейроном и эффектором. Процессы возбуждения нейронов, возникновение импульсов и распространение их по отросткам связаны с изменениями в плазмолемме. Она является структурной основой возникновения и передачи потенциалов действия. Плазмолемма имеет существенные особенности строения и функции в участках, входящих в состав синапсов.
Межнейрональные синапсы бывают нескольких видов: аксосоматические (между аксоном одного нейрона и телом другого нейрона); аксодендритические (между аксоном одного нейрона и дендритом другого нейрона); аксоаксональные (между аксонами двух нейронов). Описаны также синапсы соматосоматические, дендродендритические и др.
Все синапсы по механизму передачи импульсов между нервными клетками подразделяются на 3 типа: синапсы с химической передачей, электротонические и смешанные синапсы. Типичный синапс с химической передачей состоит из пресинаптической и постсинаптической частей, а также синаптической щели. Пресинаптическая часть включает концевое расширение аксона, ограниченное пресинаптической мембраной. Специфическими структурами этой части являются синоптические пузырьки, содержащие нейромедиаторы. Пузырьки бывают со светлым и электронно-плотным содержимым и называются в связи с этим агранулярными и гранулярными.
По форме они подразделяются на круглые и уплощенные. На внутренней поверхности пресинаптической мембраны расположены конусовидные электронно-плотные образования — пресинаптические уплотнения. В цитоплазме пресинаптической части имеются митохондрии. Синаптическая щель размером 20-30 нм содержит филаменты, связывающие наружные слои плазмолеммы контактирующих нейронов.
Постсинаптическая часть в составе плазмолеммы второго нейрона имеет рецепторы к медиатору, который выделяется в синаптическую щель при деполяризации мембраны первого нейрона. Внутренняя поверхность постсинаптической мембраны характеризуется наличием электронно-плотного слоя цитоплазмы — постсинаптические уплотнения.
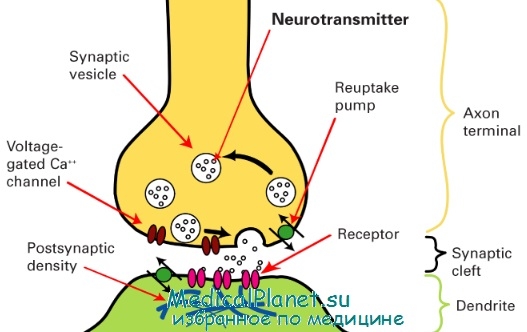
Схема строения синапса
Механизм распространения возбуждения у различных нервных волокон неодинаков.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Однако низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. Поэтому природой был выработан еще один дополнительный механизм распространения возбуждения.
Механизмы проведения возбуждения в миелиновых волокнах. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированном волокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением.
При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа + , которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности.
Особенности распространения возбуждения по миелиновым волокнам:
2. Возбуждение распространяется с большой скоростью.
3. Возбуждение распространяется без декримента.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
3. Законы проведения возбуждения по нервам.
Проведение возбуждения по нервным волокнам осуществляется по определенным законам.
Закон двустороннего проведения возбуждения по нервному волокну. Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от места его возникновения, т.е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения.Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу.
Закон анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т.е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому, основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно.
Явление парабиоза открыто русским физиологом Н.Е.Введенским в 1901 г. при изучении возбудимости нервно-мышечного препарата. Состояние парабиоза могут вызвать различные воздействия – сверхчастые, сверхсильные стимулы, яды, лекарства и другие воздействия как в норме, так и при патологии. Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т.е. воздействию повреждающего агента), то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности и было названо парабиозом. Состояние парабиоза возбудимой ткани возникает под влиянием сильных раздражителей и характеризуется фазными нарушениями проводимости и возбудимости.
Выделяют 3 фазы: первичную, фазу наибольшей активности (оптимум) и фазу сниженной активности (пессимум). Третья фаза объединяет 3 последовательно сменяющие друг друга стадии: уравнительную (провизорная), парадоксальную и тормозную.
Первая фаза (примум) характеризуется снижением возбудимости и повышением лабильности. Во вторую фазу (оптимум) возбудимость достигает максимума, лабильность начинает снижаться. В третью фазу (пессимум) возбудимость и лабильность снижаются параллельно и развивается 3 стадии парабиоза.
Первая стадия - уравнительная - характеризуется выравниванием ответов на сильные, частые и умеренные раздражения. В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители—больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т.е. 25 импульсов из 100), то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) — происходит уравнивание ответной реакции.
Вторая стадия характеризуется извращенным реагированием – сильные раздражения вызывают меньший ответ, чем умеренные. В эту - парадоксальную фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс — на редкие раздражители ответная реакция больше, чем на частые.
В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т.е. не восстанавливается ее исходное состояние. Ни сильные, ни умеренные раздражения не вызывают видимой реакции, в ткани развивается торможение.
Парабиоз — явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности. Однако, при действии сильных раздражителей за тормозной стадией может наступить полная потеря возбудимости и проводимости, а в дальнейшем – гибель ткани.
Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ вязано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
Читайте также:
