
Скорость проведения в миелиновых нервных волокон
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
Состоят из отростка нервной клетки, покрытого оболочкой, которая формируется олигодендроцитами. Отросток нервной клетки (аксон или дендрит) в составе нервного волокна называется осевым цилиндром.
- безмиелиновое (безмякотное) нервное волокно,
- миелиновое (мякотное) нервное волокно.
Безмиелиновые нервные волокна
Находятся преимущественно в составе вегетативной нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи, в которых на определенном расстоянии друг от друга видны овальные ядра. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько (10—20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в смежное. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж неиролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне
29. Миелиновые нервные волокна
1) внутренний, более толстый, — миелиновый слой,
2) наружный, тонкий, состоящий из цитоплазмы, ядер нейролеммоцитов и нейролеммы.
Миелиновый слой содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии — насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы видны участки волокна, лишенные миелинового слоя, — узловатые перехваты, или перехваты Ранвье, т.е. границы между соседними леммоцитами.
Отрезок волокна между смежными перехватами называется межузловым сегментом.
В процессе развития аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита — мезаксон. Мезаксон удлиняется, концентрически наслаивается на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. Цитоплазма с ядрами отодвигается на периферию – образуется наружная оболочка или светлая Шванновская оболочка (при окраске осмиевой кислотой).
Осевой цилиндр состоит из нейроплазмы, продольных параллельных нейрофиламентов, митохондрий. С поверхности покрыт мембраной – аксолеммой, обеспечивающей проведение нервного импульса. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Нервный импульс в миелиновом нервном волокне проводится как волна деполяризации цитолеммы осевого цилиндра, "прыгающая" (сальтирующая) от перехвата к следующему перехвату со скоростью до 120 м/сек.
В случае повреждения только отростка нейроцита регенерация возможна и протекает успешно при наличии определенных для этого условий. При этом, дистальнее места повреждения осевой цилиндр нервного волокна подвергается деструкции и рассасывается, но леммоциты при этом остаются жизнеспособными. Свободный конец осевого цилиндра выше места повреждения утолщается - образуется "колба роста", и начинает расти со скоростью 1 мм/день вдоль оставшихся в живых леммоцитов поврежденного нервного волокна, т.е. эти леммоциты играют роль "проводника" для растущего осевого цилиндра. При благоприятных условиях растущий осевой цилиндр достигает бывшего рецепторного или эффекторного концевого аппарата и формирует новый концевой аппарат.
30. шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
Каждое периферическое нервное волокно одето тонким цитоплазматическим слоем — невролеммой или шванновской оболочкой. Волокно является миелинизированным, если между ним и цитоплазмой шванновской клетки имеется значительный слой миелина. Если волокна лишены миелина, то они называются немиелинизированными безмякотными. Шванновские клетки могут осуществлять волнообразные движения, что, вероятно, способствует транспортировке различных веществ по отросткам нервных клеток.
С нарушением работы шванновских клеток связаны такие нервные заболевания, как синдром Гийена — Барре, болезнь Шарко-Мари, шванноматозис и хроническая воспалительная демиелинизирующая полинейропатия. Демиелинизация в основном происходит из-за ослабления двигательных функций шванновских клеток, в результате чего они оказываются не способны образовывать миелиновую оболочку.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Однако низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. Поэтому природой был выработан еще один дополнительный механизм распространения возбуждения.
32. Механизмы проведения возбуждения в миелиновых волокнах. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированномволокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением.
При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа + , которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности.
Особенности распространения возбуждения по миелиновым волокнам:
2. Возбуждение распространяется с большой скоростью.
3. Возбуждение распространяется без декримента.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
Сальтаторное проведение
(лат. saltatorius, от salto — скачу, прыгаю)
Сальтаторное распространение возбуждения в миелинизированном нервном волокне от перехвата к перехвату [стрелками показано направление тока, возникающего между возбуждённым (А) и соседним покоящимся (Б) перехватом].
34. Проведение нервного импульса, передача сигнала в виде волны возбуждения в пределах одного нейрона и от одной клетки к другой. П. н. и. по нервным проводникам происходит с помощью электротонических потенциалов и потенциалов действия, которые распространяются вдоль волокна в обоих направлениях, не переходя на соседние волокна (см. Биоэлектрические потенциалы, Импульс нервный). Передача межклеточных сигналов осуществляется через синапсы чаще всего с помощью медиаторов, вызывающих появление потенциалов постсинаптических. Нервные проводники можно рассматривать как кабели, обладающие относительно низким осевым сопротивлением (сопротивление аксоплазмы — ri) и более высоким сопротивлением оболочки (сопротивление мембраны — rm). Нервный импульс распространяетсявдоль нервного проводника посредством прохождения тока между покоящимися и активными участками нерва (локальные токи). В проводнике по мере увеличения расстояния от места возникновения возбуждения происходит постепенное, а в случае однородной структуры проводника экспоненциальное затухание импульса, который в 2,7 раза уменьшается на расстоянии l = (константа длины). Так как rm и riнаходятся в обратном отношении к диаметру проводника, то затухание нервного импульса в тонких волокнах происходит раньше, чем в толстых. Несовершенство кабельных свойств нервных проводников восполняется тем, что они обладают возбудимостью. Основное условие возбуждения — наличие у нервов потенциала покоя. Если локальный ток через покоящийся участок вызовет деполяризацию мембраны, достигающую критического уровня (порога), это приведёт к возникновению распространяющегося потенциала действия (ПД). Соотношение уровня пороговой деполяризации и амплитуды ПД, обычно составляющее не менее 1: 5, обеспечивает высокую надёжность проведения: участки проводника, обладающие способностью генерировать ПД, могут отстоять друг от друга на таком расстоянии, преодолевая которое нервный импульс снижает свою амплитуду почти в 5 раз. Этот ослабленный сигнал будет снова усилен до стандартного уровня (амплитуда ПД) и сможет продолжить свой путь по нерву.
Потенциал действия нервной клетки - Проведение возбуждения по нервным волокнам
- Потенциал действия нервной клетки
- Ионные механизмы потенциала действия
- Проведение возбуждения по нервным волокнам
- Законы проведения возбуждения
- Типы нервных волокон и их функции
- Все страницы
Нервные волокна — аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских [2] клеток в периферических нервах. Нервные волокна подразделяют на 2 типа — безмиелиновые и миелиновые. Основная функция нервных волокон — проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна (рис. 5–8) и существенно зависит от диаметра нервных волокон.
Рис. 5–8. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра [4]. Скорость проведения пропорциональна диаметру нервного волокна и в миелиновых волокнах выше, чем в безмиелиновых.
Безмиелиновые нервные волокна (рис. 5–9А). В покое мембрана аксона (осевого цилиндра) поляризована — положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется, и наружная поверхность мембраны приобретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны.
Рис. 5–9. Проведение возбуждения в нервных волокнах [7]. А — безмиелиновое волокно (электротоническое проведение), Б — миелиновое волокно (скачкообразное проведение). Миелин, полностью окружая аксон в межузловых промежутках, выступает в роли электрического изолятора, а межклеточная жидкость в перехватах Ранвье [3] — проводник.
Появление так называемых рефрактерных каналов (рефрактерное состояние мембраны после прохождения ПД) предупреждает распространение возбуждения в обратном направлении.
Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5–2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения ПД (см. рис. 5–8).
Миелиновое нервное волокно (рис. 5–9Б) состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания собственной плазматической мембраны. Миелин прерывается через регулярные промежутки (от 0,2 до 2 мм) концентрической щелью шириной около 1 мкм, это узлы, или перехваты Ранвье. Таким образом, межузловые сегменты аксона, расположенные между соседними перехватами Ранвье, содержат миелин — электрический изолятор, не позволяющий проходить через него локальным токам, поэтому ПД возникают только в перехватах Ранвье. Другими словами, ПД перемещается вдоль нервного волокна скачками, от одного перехвата Ранвье к другому перехвату (скачкообразное проведение).
Плотность потенциалозависимых Na+?каналов аксолеммы в перехватах Ранвье — до 2000 на 1 мкм2 (в перикарионе — 50–70, в начальном сегменте аксона — 2000, в межузловых сегментах Na+?каналы практически отсутствуют). В силу высокой плотности Na+?каналов перехваты Ранвье характеризуются высокой возбудимостью, а локальные токи достаточно велики для возбуждения соседнего перехвата.
Локальные токи текут от перехвата к перехвату (через внеклеточную жидкость кнаружи от миелина и через аксоплазму внутри аксона) с минимальными потерями.
Энергозатраты нервного волокна на проведение ПД относительно невелики, поскольку возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% общей поверхности мембраны аксона. Поэтому даже после длительных ритмических пачек ПД трансмембранный градиент концентраций ионов практически не изменяется.
В физиологических условиях ПД движутся в одном направлении от места раздражения (ортодромное проведение). ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения. Проведение в противоположном направлении (антидромное проведение) возможно при травматическом поражении нервных волокон и в редких случаях (аксон–рефлекс).
Нарушение миелинизации нервных волокон приводит к нарушениям проводимости (демиелинизирующие заболевания). При разрушении миелиновой оболочки происходит резкое снижение скорости и надёжности проведения возбуждения по нервам. Наиболее распространённым среди демиелинизирующих заболеваний является множественный склероз, проявляющийся различными параличами и потерей чувствительности.
Нервная система человека и позвоночных животных имеет единый план строения и представлена центральной частью – головным и спинным мозгом, а также периферическим отделом – отходящими от центральных органов нервами, представляющими собой отростки нервных клеток – нейронов.
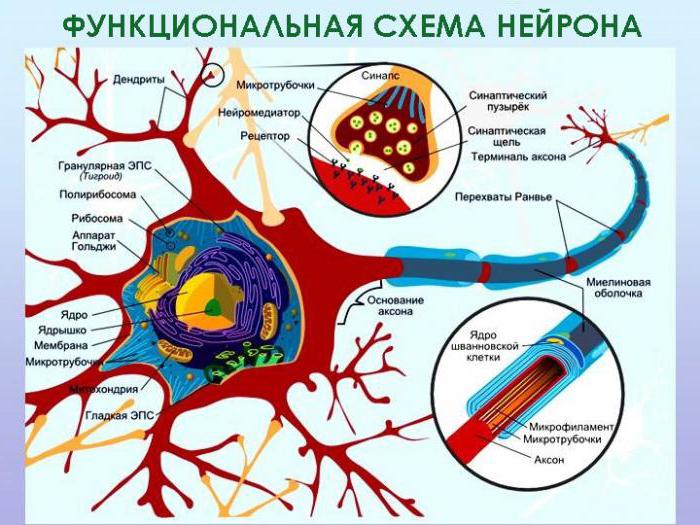
Их совокупность образует нервную ткань, главными функциями которой является возбудимость и проводимость. Эти её свойства объясняются прежде всего особенностями строения оболочек нейронов и их отростков, состоящих из вещества, названного миелином. В данной статье мы рассмотрим строение и функции этого соединения, а также выясним возможные способы его восстановления.
Почему нейроциты и их отростки покрыты миелином
Совсем не случайно дендриты и аксоны имеют защитный слой, состоящий из белково-липидных комплексов. Дело в том, что возбуждение является биофизическим процессом, в основе которого лежат слабые электрические импульсы. Если электрический ток идёт по проводу, то последний должен быть покрыт изоляционным материалом, чтобы уменьшить рассеивание электрических импульсов и не допустить снижение силы тока. Такие же функции в нервном волокне выполняет миелиновая оболочка. Кроме того, она является опорой, а также обеспечивает питание волокна.
Химический состав миелина
Как и большинство клеточных мембран, он имеет липопротеидную природу. Причём содержание жиров здесь очень высокое – до 75%, а белков – до 25%. Миелин в незначительном количестве содержит также гликолипиды и гликопротеиды. Химический состав его различается в спинномозговых и в черепно-мозговых нервах.
В первых наблюдается высокое содержание фосфолипидов – до 45%, а остальная часть приходится на холестерин и цереброзиды. Демиелинизация (то есть замена миелина на другие вещества в нервных отростках) приводит к таким тяжёлым аутоиммунным заболеваниям, как, например, рассеянный склероз.
С химической точки зрения, этот процесс будет выглядеть так: миелиновая оболочка нервных волокон меняет свою структуру, что проявляется прежде всего в уменьшении процентного содержания липидов по отношению к белкам. Далее снижается количество холестерина и возрастает содержание воды. А всё это приводит к постепенной замене миелина, содержащего олигодендроциты или шванновские клетки на макрофаги, астроциты и межклеточную жидкость.
Результатом таких биохимических изменений будет резкое снижение способности аксонов проводить возбуждение вплоть до полной блокировки прохождения нервных импульсов.
Особенности нейроглиальных клеток
Как мы уже говорили, миелиновая оболочка дендритов и аксонов образована специальными структурами, характеризующимися низкой степенью проницаемости для ионов натрия и кальция, а потому имеющих только потенциалы покоя (они не могут проводить нервные импульсы и выполняют электроизоляционные функции).
Данные структуры называются глиальными клетками. К ним относятся:
- олигодендроциты;
- волокнистые астроциты;
- клетки эпендимы;
- плазматические астроциты.
Все они формируются из наружного слоя зародыша – эктодермы и имеют общее название – макроглия. Глия симпатических, парасимпатических и соматических нервов представлена шванновскими клетками (нейролеммоцитами).
Строение и функции олигодендроцитов
Они входят в состав центральной нервной системы и являются клетками макроглии. Так как миелин – это белково-липидная структура, она способствует увеличению скорости проведения возбуждения. Сами клетки образуют электроизолирующий слой нервных окончаний в головном и спинном мозге, формируясь уже в период внутриутробного развития. Их отростки обворачивают в складки своей наружной плазмалеммы нейроны, а также дендриты и аксоны. Получается, что миелин – это основной электроизолирующий материал, разграничивающий нервные отростки смешанных нервов.
Шванновские клетки и их особенности
Миелиновая оболочка нервов периферической системы образована нейролеммоцитами (шванновскими клетками). Их отличительная особенность состоит в том, что они способны образовывать защитную оболочку только одного аксона, и не могут формировать отростки, как это присуще олигодендроцитам.
Между шванновскими клетками на расстоянии 1-2 мм располагаются участки, лишённые миелина, так называемые перехваты Ранвье. По ним скачкообразно происходит проведение электрических импульсов в пределах аксона.
Леммоциты способны к репарации нервных волокон, а также выполняют трофическую функцию. В результате генетических аббераций клетки оболочки леммоцитов начинают неконтролируемое митотическое деление и рост, вследствие чего в различных отделах нервной системы развиваются опухоли – шванномы (невриномы).
Роль микроглии в разрушении миелиновой структуры
Микроглия представляет собой макрофаги, способные к фагоцитозу и умеющие распознавать различные патогенные частицы – антигены. Благодаря мембранным рецепторам эти глиальные клетки вырабатывают ферменты – протеазы, а также цитокины, например, интерлейкин 1. Он является медиатором воспалительного процесса и иммунитета.
Более того, цитокины, активируя рецепторы, провоцируют избыточный транспорт ионов кальция в тело нейрона. Протеазы и фосфолипазы начинают расщеплять органеллы и отростки нервных клеток, что приводит к апоптозу – гибели данной структуры.
Она разрушается, распадаясь на частицы, которые и пожирают макрофаги. Это явление называется эксайтотоксичностью. Оно вызывает дегенерацию нейронов и их окончаний, приводя к таким заболеваниям, как болезнь Альцгеймера и болезнь Паркинсона.
Мякотные нервные волокна
Если отростки нейронов – дендриты и аксоны, покрывает миелиновая оболочка, то они называются мякотными и иннервируют скелетную мускулатуру, входя в соматический отдел периферической нервной системы. Немиелинизированные волокна образуют вегетативную нервную систему и иннервируют внутренние органы.
Мякотные отростки имеют больший диаметр, чем безмякотные, и формируются следующим образом: аксоны прогибают плазматическую мембрану клеток глии и формируют линейные мезаксоны. Затем они удлиняются и шванновские клетки многократно обворачиваются вокруг аксона, образуя концентрические слои. Цитоплазма и ядро леммоцита перемещаются в область наружного слоя, который называется неврилеммой или шванновской оболочкой.
Внутренний слой леммоцита состоит из слоистого мезоксона и называется миелиновой оболочкой. Толщина её в различных участках нерва неодинакова.
Как восстановить миелиновую оболочку
Рассматривая роль микроглии в процессе демиелинизации нервов, мы установили, что под действием макрофагов и нейромедиаторов (например, интерлейкинов) происходит разрушение миелина, что в свою очередь приводит к ухудшению питания нейронов и нарушению передачи нервных импульсов по аксонам.
Данная патология провоцирует возникновение нейродегенеративных явлений: ухудшение когнитивных процессов, прежде всего памяти и мышления, появление нарушения координации движений тела и тонкой моторики.
В итоге возможна полная инвалидизация больного, которая возникает в результате аутоиммунных заболеваний. Поэтому вопрос о том, как восстановить миелин, в настоящее время стоит особенно остро. К таким способам относится прежде всего сбалансированная белково-липидная диета, правильный образ жизни, отсутствие вредных привычек. В тяжелых случаях заболеваний применяют медикаментозное лечение, восстанавливающее количество зрелых глиальных клеток – олигодендроцитов.
Развитие аксона сопровождается его погружением в шванновскую клетку и образованием миелиновой оболочки (рис. 4.20). При этом аксон никогда не контактирует с цитоплазмой шванновской клетки, а погружается в углубление ее мембраны. Края этой мембраны смыкаются над аксоном, образуя удвоенную мембрану, которая несколько раз наматывается вокруг аксона в виде спирали. На более поздних стадиях спираль закручивается более плотно и образуется компактная миелиновая оболочка. Ее толщина в крупных нервах может достигать 2—3 мкм.
Миелиновая оболочка образуется в нескольких микронах от тела клетки, сразу за аксонным холмиком, и покрывает все нервное волокно. Отсутствие такой оболочки ограничивает функциональные возможности нервного волокна: снижается скорость проведения возбуждения по нему.
Раньше других начинают миелинизировагься периферические нервы, затем аксоны в спинном мозге, стволовой части головного мозга, мозжечке и позже — в больших иолуша- риях головного мозга.

Рис. 4.20. Образование миелиновой оболочки нервного волокна в периферической нервной системе (а) и в ЦНС (б)
Миелинизация спинномозговых и черепно-мозговых нервов начинается на четвертом месяце внутриутробного развития. Двигательные волокна покрываются миелином к моменту рождения ребенка, а большинство смешанных и чувствительных нервов — к трем месяцам после рождения. Многие черепно-мозговые нервы миелинизируются к полутора-двум годам. К двум годам миелинизируются слуховые нервы. Полная миелинизация зрительного и языкоглоточного нервов отмечается только у трех-четырехлетних детей, у новорожденных они еще не миелинизированы. Ветви лицевого нерва, иннервирующие область губ, миелинизируются с 21-й до 24-й недели внутриутробного периода, другие его ветви приобретают миелиновую оболочку значительно позже. Этот факт свидетельствует о раннем формировании морфологических структур, при участии которых осуществляется сосательный рефлекс, хорошо выраженный к моменту рождения ребенка.
Проводящие пути спинного мозга хорошо развиты к моменту рождения и почти все миелинизированы, за исключением пирамидных путей (они миелинизируются к третьему — шестому месяцам жизни ребенка). В спинном мозге раньше других миелинизируются моторные пути. Еще во внутриутробный период они оказываются сформированными, что проявляется в спонтанных движениях плода.
Миелинизация нервных волокон в головном мозге начинается во внутриутробном периоде развития и закапчивается после рождения (рис. 4.21). В отличие от спинного мозга, здесь раньше других миелинизируются афферентные пути и сенсорные области, а двигательные — через пять-шесть месяцев, а некоторые и значительно позже после рождения. К трем годам миелинизация нервных волокон в основном заканчивается, но рост нервов в длину продолжается и после трехлетнего возраста.
В процессе развития мозга в формировании упорядоченных связей между миллиардами нервных клеток решающая роль принадлежит активности самих нейронов, а также влиянию внешних факторов.
Хотя человек рождается с полным набором нейронов, которые образуются в эмбриональный период, мозг новорожденного по массе составляет 1/10 часть мозга взрослого. Увеличение массы мозга происходит за счет увеличения размеров нейронов, а также числа и длины их отростков.
Процесс развития нервных сетей можно разделить на три этапа. Первый этап включает образование незрелых нейронов (нейробластов) путем деления в соответствии с генетической программой. Незрелый нейрон, еще не имеющий аксона и дендритов, обычно мигрирует из места своего образования в соответствующий участок нервной системы. Нейроны могут мигрировать на большие расстояния. Способ их перемещения напоминает движение амебы. Миграцию направляют глиальные клетки (рис. 4.22, а). Незрелые мигрирующие нейроны тесно примыкают к глиальным клеткам и как бы ползут по ним. Достигнув своего постоянного места расположения, клетка образует контакты с другими нейрона-

Рис. 4.21. Сроки мислинизации основных путей мозга в раннем онтогенезе человека (по Л. О. Бадаляну)

Рис. 4.22. Миграция нейронов в развивающемся мозге:
а — незрелые нервные клетки, мигрирующие вдоль отростков радиальных глиальных клеток; 6 — постепенное утолщение стенки нервной трубки и установление ориентации пирамидных нейронов будущей коры больших
ми. Сразу же устанавливается ориентация клеток: например, пирамидные нейроны выстраиваются в ряды так, что их ден- дриты направлены к поверхности коры, а аксоны — в подлежащее белое вещество (рис. 4.22, б).
Определенные группы нейронов выделяют специфические метки, которые узнаются другими нейронами, благодаря этому возможно установление высокоизбирательных нервных связей. Кроме того, имеются специфические биологически активные вещества, ускоряющие рост нейронов. Например, фактор роста нервов влияет на рост и созревание нейронов спинальных и симпатических ганглиев.
Важными моментами в процессе развития нейрона считают появление способности к генерации и проведению нервных импульсов, а также формирование синаптических контактов.
В процессе онтогенеза в нейронах происходят и другие изменения. Так, после рождения увеличиваются длина и диаметр аксонов (рис. 4.23) и продолжается их миелинизация. Эти процессы заканчиваются в основном к 9—10 годам. При этом существенно повышается скорость проведения возбуждения по нервным волокнам: у новорожденных она составляет только 5% уровня взрослых. Другая причина увеличения

Рис. 4.23. Развитие нервных клеток человека в процессе онтогенеза
скорости проведения импульсов — возрастание числа ионных каналов в нейронах, повышение мембранного потенциала и амплитуды ПД. Эффекты положительного влияния стимуляции на развитие мозга ограничены чувствительным периодом. Ослабление стимуляции в этот период не лучшим образом сказывается на морфофункциональном формировании мозга.
Поступление достаточного объема многосторонней информации в развивающийся мозг способствует появлению нейронов, специфически реагирующих на сложные комбинации сигналов. Этот механизм, по-видимому, лежит в основе способности человека отражать реально существующие феномены внешнего мира на основе индивидуального (субъективного) опыта.
Связи между центральными нейронами наиболее активно формируются в период от рождения до 3 лет (рис. 4.24; 4.25). От того, как нейроны соединяются друг с другом на начальных этапах формирования мозга, во многом зависят его индивидуальные особенности. Информация, поступающая в мозг,

Рис. 4.24. Развитие нейронных сетей коры больших полушарий головного мозга в первые 2 года жизни ребенка
обеспечивает создание все новых сочетаний соединений и увеличение числа контактов между нейронами за счет роста их дендритов. Интенсивная нагрузка мозга до самого преклонного возраста защищает его от преждевременной деградации. Известно, что у образованных людей, постоянно пополняющих свои знания, число связей между нейронами возрастает, причем высокий уровень образования даже снижает опасность заболеваний, связанных с нарушением этих связей.
Известно, что у человека после рождения каждый нейрон па протяжении жизни сохраняет способность к росту, обра-

Рис. 4.25. Развитие двигательных умений у ребенка от рождения до 15 месяцев
зованию отростков и новых синаптических связей, особенно при наличии интенсивной сенсорной информации. Под ее влиянием синаптические связи могут также перестраиваться и менять медиатор. Это свойство лежит в основе процессов научения, памяти, адаптации к постоянно меняющимся условиям внешней среды, восстановительных процессов в период реабилитации после различных заболеваний и перенесенных травм.
Читайте также:
