
Слуховой анализатор нервный центр
Слуховые проводящие пути и низшие слуховые центры - это проводниковая афферентная (приносящая) часть слуховой сенсорной системы, проводящая, распределяющая и преобразующая сенсорное возбуждение, порождённое слуховыми рецепторами, для формирования рефлекторных реакций эффекторов и слуховых образов в высших слуховых центрах коры.
Все слуховые центры, начиная от кохлеарных ядер и вплоть до коры головного мозга, устроены тонотопически, т.е. рецепторы кортиева органа проецируются в них на строго определенные нейроны. И, соответственно, эти нейроны обрабатывают информацию о звуках только определённой частоты, определённой высоты звучания. Чем дальше по слухового пути от улитки находится слуховой центр, тем более сложными звуковыми сигналами возбуждаются его отдельные нейроны. это говорит о том, что в слуховых центрах происходит всё более усложняющийся синтез отдельных характеристик звуковых сигналов.
Нельзя считать, что информация о звуковых сигналах обрабатывается только последовательно при переходе возбуждения от одного слухового центра к другому. Все слуховые центры связаны между собой многочисленными сложными связями, с помощью которых осуществляется не только перенос информации в одном направлении, но и её сравнительная обработка.
1 - улитка (Кортиев орган с волосковыми клетками - слуховыми рецепторами);
2 - спиральный ганглий;
3 - переднее (вентральное) улитковое (кохлеарное) ядро;
4 - заднее (дорзальное) улитковое (кохлеарное) ядро;
5 - ядро трапециевидного тела;
6 - верхняя олива;
7 - ядро латеральной петли;
8 - ядра задних холмиков четверохолмия среднего мозга;
9 - медиальные коленчатые тела метаталамуса промежуточного мозга;
10 - проекционная слуховая зона коры больших полушарий головного мозга.

Рис. 1. Схема слуховых сенсорных путей (по Сентаготаи).
1 — височная доля; 2 — средний мозг; 3 — перешеек ромбовидного мозга; 4 — продолговатый мозг; 5 — улитка; 6 — вентральное слуховое ядро; 7 — дорсальное слуховое ядро; 8 — слуховые полоски; 9 — оливо-слуховые волокна; 10 — верхняя олива: 11 — ядра трапециевидного тела; 12 — трапециевидное тело; 13 — пирамида; 14 — латеральная петля; 15 — ядро латеральной петли; 16 — треугольник латеральной петли; 17 — нижнее двухолмие; 18 — латеральное коленчатое тело; 19 — корковый центр слуха.
Схематический путь слухового возбуждения : слуховые рецепторы (волосковые клетки в Кортиевом органе улитки уха) - периферический спиральный ганглий (в улитке) - продолговатый мозг (сначала кохлеарные ядра, т.е. улитковые, после них - ядра оливы) - средний мозг (нижнее двухолмие) - промежуточный мозг (медиальные коленчатые тела, они же внутренние) - кора больших полушарий головного мозга (слуховые зоны височных долей, поля 41, 42).
Первые (I) слуховые афферентные нейроны (биполярные нейроны) находятся в спиральном ганглии, или узле (gangl. spirale), расположенном в основании полого улиткового веретена. Спиральный ганглий состоит из тел слуховых биполярных нейронов. Дендриты этих нейронов проходят по каналам костной спиральной пластинки к улитке уха, т.е. они начинаются от наружных волосковых клеток Кортиева органа. Аксоны выходят из спирального узла и собираются в слуховой нерв, вступающий в области мостомозжечкового угла в ствол мозга, где и заканчиваются синапсами на нервных клетках улитковых (кохлеарных) ядер: дорсального (nucl. cochlearis dorsalis) и вентрального (nucl. cochlearis ventralis). Эти клетки кохлеарных ядер являются вторыми слуховыми нейронами (II).
Слуховой нерв имеет следующие названия: N. vestibulocochlearis, sive n. octavus (PNA), n. acusticus (BNA), sive n. stato-acusticus - равновесно-слуховой (JNA). Это VIII пара черепно-мозговых нервов, состоящая из двух частей: улитковой (pars cochlearis) и вестибулярной, или преддверной (pars vestibularis). Улитковая часть является совокупностью аксонов I нейронов слуховой сенсорной системы (биполярных нейронов спирального ганглия), преддверная - аксоны афферентных нейронов лабиринта, обеспечивающие регулирование положения тела в пространстве (в анатомической литературе обе части также называются нервными корешками).
Вторые слуховые афферентные нейроны (II) находятся в дорсальном и вентральном кохлеарном (улитковом) ядре продолговатого мозга.
От нейронов II кохлеарных ядер начинаются два восходящих слуховых тракта. Контрлатеральный восходящий слуховой путь содержит в себе основную массу выходящих из комплекса кохлеарных ядер волокон и образует три пучка волокон: 1- вентральная слуховая полоска, или трапециевидное тело, 2 - промежуточная слуховая полоска, или полоска Хельда, 3 - задняя, или дорсальная, слуховая полоска - полоска Монакова. Основную часть волокон содержит в себе первый пучок - трапециевидное тело. Средняя, интермедиальная, полоска образована аксонами части клеток заднего отдела заднего вентрального ядра кохлеарного комплекса. Дорсальная слуховая полоска содержит в себе волокна, идущие от клеток дорсального кохлеарного ядра, а также аксоны части клеток заднего вентрального ядра. Волокна дорсальной полоски идут по дну четвертого желудочка, затем уходят в ствол мозга, пересекают среднюю линию и, минуя оливу, не оканчиваясь в ней, присоединяются к латеральной петле противоположной стороны, где поднимаются к ядрам латеральной петли. Эта полоска обходит верхнюю ножку мозжечка, затем переходит на противоположную сторону и присоединяется к трапециевидному телу.
Итак, аксоны II нейронов, отходящие от клеток дорсального ядра (слухового бугорка), образуют мозговые полоски (striae medullares ventriculi quarti), находящиеся в ромбовидной ямке на границе моста и продолговатого мозга. Большая часть мозговой полоски переходит на противоположную сторону и около средней линии погружается в вещество мозга, подключаясь к латеральной петле (lemniscus lateralis); меньшая часть мозговой полоски присоединяется к латеральной петле своей же стороны. Многочисленные волокна, выходящие из дорсального ядра, идут в составе боковой петли и оканчиваются в нижних бугорках четверохолмия среднего мозга (colliculus inferior) и во внутреннем (медиальном) коленчатом теле (corpus geniculatum mediate) таламуса, это промежуточный мозг. Часть волокон, минуя внутреннее коленчатое тело (слуховой центр), идет в наружное (латеральное) коленчатое тело таламуса, являющееся зрительным подкорковым центром промежуточного мозга, что указывает на тесную связь между слуховой сенсорной системой и зрительной.
Аксоны II нейронов от клеток вентрального ядра участвуют в образовании трапециевидного тела (corpus trapezoideum). Большая часть аксонов в составе боковой петли (lemniscus lateralis) переходит на противоположную сторону и оканчивается в верхней оливе продолговатого мозга и ядрах трапециевидного тела, а также в ретикулярных ядрах покрышки на слуховых нейронах III. Другая, меньшая, часть волокон оканчивается на своей же стороне в тех же структурах. Поэтому именно здесь, в оливах, проходит сравнение акустических сигналов, поступающих с двух сторон от двух разных ушей. Оливы обеспечивают бинауральный анализ звуков, т.е. сопоставляют звуки от разных ушей. Именно оливы обеспечивают стереозвучание и помогают точно нацелиться на источник звука.
Третьи слуховые афферентные нейроны (III) находятся в ядрах верхней оливы (1) и трапециевидного тела (2), а также в нижнем двухолмии среднего мозга (3) и в внутренних (медиальных) коленчатых телах (4) промежуточного мозга. Аксоны III нейронов участвуют в образовании латеральной петли, в которой имеются волокна II и III нейронов. Часть волокон II нейронов прерывается в ядре латеральной петли (nucl. lemnisci proprius lateralis). Таким образом, в ядре латеральной петли тоже есть III нейроны Волокна II нейронов латеральной петли переключаются на III нейроны в медиальном коленчатом теле (corpus geniculatum mediale). Волокна III нейронов латеральной петли, пройдя мимо медиального коленчатого тела, заканчиваются в нижнем двухолмии (colliculus inferior), где формируется tr. tectospinalis. Таким образом, в нижнем двухолмии среднего мозга находится низший слуховой центр, состоящий из IV нейронов.
Нервные волокна латеральной петли, которые относятся к нейронам верхней оливы, из моста проникают в верхние ножки мозжечка и затем достигают его ядер. Таким образом, ядра мозжечка получают слуховое сенсорное возбуждение из слуховых низших нервных центров оливы. Другая часть аксонов верхней оливы направляется к мотонейронам спинного мозга и далее к поперечнополосатым мышцам. Таким образом, слуховые низшие нервные центры верхней оливы управляют эффекторами и обеспечивают двигательные слуховые рефлекторные реакции.
Аксоны III нейронов, расположенных в медиальном коленчатом теле (corpus geniculatum mediate), пройдя через заднюю часть задней ножки внутренней капсулы, формируют слуховое сияние, которое заканчивается на IV нейронах в высшем нервном слуховом центре коры - поперечной извилине Гешля височной доли (поля 41, 42, 20, 21, 22). Итак, аксоны III нейронов медиальных коленчатых тел образуют центральный слуховой путь, ведущий в слуховые сенсорные первичные проекционные зоны коры больших полушарий головного мозга. Кроме восходящих афферентных волокон, в центральном слуховом пути проходят также и нисходящие эфферентные волокна - от коры к низшим подкорковым слуховым центрам.
Четвёртые слуховые афферентные нейроны (IV) находятся как в нижнем двухолмии среднего мозга, так и в височной доле коры больших полушарий головного мозга (поля 41, 42, 20, 21, 22 по Бродману).
Нижнее двухолмие является рефлекторным двигательным центром, через который подключается tr. tectospinalis. Благодаря этому при слуховом раздражении рефлекторно подключается спинной мозг для выполнения автоматических движений, чему способствует и подключение верхней оливы с мозжечком; подключается также медиальный продольный пучок (fasc. longitudinalis medialis), объединяющий функции двигательных ядер черепных нервов. Разрушение нижнего двухолмия не сопровождается потерей слуха, однако оно играет важную роль "рефлекторного" подкоркового центра, в котором формируется эфферентная часть ориентировочных слуховых рефлексов в виде движения глаз и головы.
Тела корковых нейронов IV образуют колонки слуховой коры, формирующих первичные слуховые образы. От некоторых IV нейронов идут пути через мозолистое тело на противоположную сторону, в слуховую кору контралатерального (противоположного) полушария. Это последний путь слухового сенсорного возбуждения. Он заканчивается тоже на IV нейронах. Слуховые сенсорные образы формируются в высшем нервном слуховом центре коры - поперечной извилине Гешля височной доли (поля 41, 42, 20, 21, 22). Низкие звуки воспринимаются в передних отделах верхней височной извилины, а высокие звуки — в её задних отделах. Поля 41 и 42, а также 41/42 височной области коры относятся к мелкоклеточным (пылевидным, кониокортикальным) чувствующим полям коры мозга. Они располагаются на верхней поверхности височной доли, скрытой в глубине латеральной (сильвиевой) борозды. В поле 41, наиболее мелко- и густоклеточном, заканчивается большая часть афферентных волокон слуховой сенсорной системы. Другие поля височной области (22, 21, 20 и 37) выполняют высшие слуховые функции, например, участвуют в слуховом гнозисе. Слуховой гнозис (gnosis acustica) - это узнавание предмета по характерному для него звуку.
При заболевании периферических отделов слуховой сенсорной системы в слуховом восприятии возникают шумы, звуки различного характера.
Для понижения слуха центрального происхождения характерно нарушение высшего акустического (звукового) анализа звуковых раздражений. Иногда отмечается патологическое обострение или извращение слуха (гиперакузия, паракузия).
При корковом поражении наступают сенсорная афазия и слуховая агнозия. Расстройство слуха наблюдается при многих органических заболеваниях центральной нервной системы.
Нервный импульс возникает в волосковых клетках, передается биполярным нервным клеткам, расположенным в спиральном ганглии улитки (первый нейрон). Центральные отростки клеток спирального ганглия образуют слуховой, или кохлеарный, нерв (VIII пара черепно-мозговых нервов). Кохлеарный нерв проходит в продолговатый мозг и заканчивается на клетках кохлеарных ядер (второй нейрон). Нервные волокна от кохлеарных ядер в составе боковой петли доходят до верхней оливы (третий нейрон). Одна часть волокон латеральной петли достигает среднего мозга — ядер нижних бугров четверохолмия, другая — медиального коленчатого тела зрительных бугров, где происходит переключение и находится четвертый нейрон. Далее волокна в составе слуховой радиации заканчиваются в коре верхней части височной доли большого мозга (поля 41 и 42 по Бродману), т.е. в центральной части слухового анализатора.
Функция отдельных частей проводящей системы слухового анализатора состоит в следующем. В спиральном ганглии методом разрушения и перерезок было показано пространственно раздельное представительство низких и высоких частот. Так, частичная перерезка волокон слухового нерва приводит к потере слуха на высоких частотах. При полной перерезке слухового нерва происходит потеря слуха на низких частотах.
Нижние бугры четверохолмия отвечают за ориентировочный рефлекс (поворот головы в сторону источника звука). Слуховая кора принимает участие в переработке звуковой информации в процессе дифференцировки звуков, она отвечает за бинауральный слух.
Электрические явления в улитке
При отведении электрических потенциалов от разных частей улитки различают пять электрических феноменов: 1. Мембранный потенциал волосковых клеток, равный —80 мв. Регистрируется при введении в нее микроэлектрода. 2. Эндокохлеарный потенциал — регистрируется при прохождении микроэлектрода через каналы улитки. Эндолимфа, содержащая много ионов калия, имеет положительный заряд по отношению к перилимфе верхнего и нижнего каналов, он равен +80 мв. Эндокохлеарный потенциал создается за счет функционирования сосудистой полоски и обусловлен определенным уровнем окислительно-восстановительных реакций. Он является источником энергии для процесса преобразования воздействующего раздражителя в нервный импульс. Разрушение сосудистой полоски и гипоксия приводят к исчезновению эндокохлеарного потенциала. 3. Микрофонный потенциал, или эффект, возникает в улитке при действии звука, является физическим явлением и полностью отражает форму звуковых волн. Он регистрируется при помещении электродов в барабанной лестнице вблизи от кортиева органа или на круглом окне. Этот потенциал аналогичен выходному напряжению микрофона, и если его подать на усилитель и пропустить через громкоговоритель, то получим воспроизведение речи. Происхождение микрофонного эффекта не совсем ясно, его связывают с механохимическими преобразованиями в волосковых клетках кортиева органа, повреждение которого приводит к исчезновению микрофонного эффекта. 4. Суммационный потенциал: при действии звуков большой силы и частоты происходит стойкое изменение нулевой линии на записи электрических колебаний или сдвиг исходной разности потенциалов — это суммационный потенциал, который, в отличие от микрофонного, воспроизводит не форму звуковой волны, а ее огибающую. 5. Потенциалы действия слухового нерва регистрируются при отведении от волокон слухового нерва. Их частота зависит от высоты действующего на ухо тона, но до определенных пределов. Если частота звуковых колебаний не превышает 1000 в секунду, то в слуховом нерве возникают импульсы такой же частоты. При действии на ухо высокочастотных колебаний частота импульсов в слуховом нерве ниже, чем частота звуковых колебаний. Потенциал действия слухового нерва является результатом синаптической передачи возбуждения в нервных элементах кортиева органа с участием медиатора (возможно, глутамата).
Механизм восприятия звуков различной частоты
Существуют две теории восприятия звуков. Согласно резонансной теории слуха Г.Д.Гельмгольца (1885 г.), базилярная мембрана состоит из отдельных волокон (струн резонатора), настроенных на звуки определенной частоты. Так, звуки высокой частоты воспринимаются короткими волокнами базилярной мембраны, расположенными ближе к основанию улитки, низкой частоты — длинными волокнами вершины улитки. Теория места основана на различной способности волосковых клеток, расположенных в разных местах базилярной мембраны, воспринимать звуки различной частоты. Повреждение отдельных участков базилярной мембраны с волосковыми клетками приводит к повышению порога восприятия звуков определенной частоты.
Слуховая адаптация
Понижение слуховой чувствительности, развивающееся в процессе длительного действия звука большой интенсивности или после его прекращения, называют слуховой адаптацией. Она обусловлена изменениями как в периферических, так и центральных отделах слухового анализатора. Ухо, адаптированное к тишине, обладает более низким порогом слуховой чувствительности. При длительном действии звуков большой интенсивности (громкая музыка, работа в шумных цехах) порог слуховой чувствительности повышается.
Пространственный слух
Способность человека и животного локализовать источник звука в пространстве называется пространственным слухом. Слуховая ориентация осуществляется двумя путями: определением местоположения самого звучащего объекта [первичная локализация) и с помощью эхолокации, т.е. восприятием отраженных от различных объектов звуковых волн. Эхолокация помогает ориентироваться в пространстве некоторым животным (дельфинам, летучим мышам), а также людям, потерявшим зрение. Пространственное восприятие звука возможно при наличии бинаурального слуха, т.е. способности определить местонахождение источника звука одновременно правым и левым ухом.

Слуховые пути начинаются в улитке в нейронах спирального узла (первый нейрон). Дендриты этих нейронов иннервируют кортиев орган, аксоны оканчиваются в двух ядрах моста – переднем (вентральном) и в заднем (дорзальном) улитковых ядрах. От вентрального ядра импульсы поступают к следующим ядрам (оливам) своей и чужой стороны, нейроны которых, таким образом, получают сигналы от обоих ушей. Именно здесь проходит сравнение акустических сигналов, поступающих с двух сторон от организма. От дорзальных ядер импульсы поступают через нижние холмики четверохолмия и медиальное коленчатое тело в первичную слуховую кору – задний отдел верхней височной извилины.
Схема путей слухового анализатора
2 - спиральный ганглий;
3 - переднее (вентральное) улитковое ядро;
4 - заднее (дорзальное) улитковое ядро;
5 - ядро трапециевидного тела;
6 -верхняя олива;
7 - ядро латеральной петли;
8 - ядра задних холмиков;
9 -медиальные коленчатые тела;
10 - проекционная слуховая зона.
Возбуждение нейронов периферических слуховых нейронов, подкорковых и корковых первичных клеток происходит при предъявлении слуховых раздражителей разной сложности. Чем дальше от улитки по слуховому тракту, тем более сложные звуковые характеристики требуются для активации нейронов. Первичные нейроны спирального ганглия могут возбуждаться чистыми тонами, в то время как уже в ядрах улитки одночастотный звук может вызвать торможение. Для возбуждения нейронов требуются звуки различных частот.
В нижних холмиках четверохолмия есть клетки, реагирующие на частотно-модулированные тоны со специфическим направлением. В слуховой зоне коры есть нейроны, которые отвечают только на начало звукового стимула, другие – только на его окончание. Некоторые нейроны возбуждаются при звуках определенной длительности, другие – при повторяющихся звуках. Информация, содержащаяся в звуковом стимуле, многократно перекодируется по мере прохождения через все уровни слухового тракта. Благодаря сложным процессам интерпретации происходит распознавание слуховых образов, что очень важно для понимания речи.
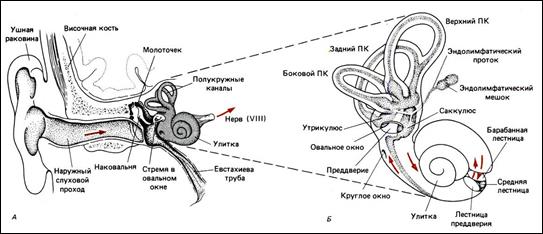
Ухо млекопитающих как орган равновесия

У позвоночных органы равновесия расположены в перепончатом лабиринте, который развивается из переднего конца системы боковой линии рыб. Они состоят из двух камер – круглого мешочка (саккулюса) и овального мешочка (маточки, утрикулюса) – и отходящих от овального мешочка трех полукружных каналов, которые лежат в трех взаимно перпендикулярных плоскостях, в полостях одноименных костных каналов. Одна из ножек каждого протока, расширяясь, образует перепончатые ампулы. Участки стенки мешочков, выстланные чувствительными рецепторными клетками, называются пятнами, аналогичные участки ампул полукружных каналов – гребешками.
Эпителий пятен содержит рецепторные волосковые клетки, на верхних поверхностях которых имеется по 60 – 80 волосков (микроворсинок), обращенных в полость лабиринта. Кроме волосков, каждая клетка снабжена одной ресничкой. Поверхность клеток покрыта студенистой мембраной, содержащей статолиты – кристаллы углекислого кальция. Мембрана поддерживается статическими волосками волосковых клеток. Рецепторные клетки пятен воспринимают изменения силы тяжести, прямолинейные движения и линейные ускорения.
Гребешки ампул полукружных каналов выстланы аналогичными волосковыми клетками и покрыты желатинообразным куполом – купулой, в который проникают реснички. Они воспринимают изменение углового ускорения. Три полукружных канала великолепно приспособлены для сигнализации о движениях головы в трехмерном пространстве.
При изменении силы тяжести, положения головы, тела, при ускорении движения и т. п. мембраны пятен и купулы гребешков смещаются. Это приводит к напряжению волосков, что вызывает изменение активности различных ферментов волосковых клеток и возбуждение мембраны. Возбуждение передается нервным окончаниям, которые разветвлены, и окружают рецепторные клетки наподобие чаш, формируя синапсы с их телами. В конечном итоге возбуждение передается ядрам мозжечка, спинному мозгу и коре теменной и височной долей больших полушарий, где находится корковый центр анализатора равновесия.
Слуховые центры можно подразделить на стволовые, подкорковые и корковые. Являясь относительно молодыми в филогенетическом отношении, слуховые центры отличаются полиморфизмом нейронного строения и обладают богатыми связями с филогенетически старыми образованиями (ретикулярной формацией, другими сенсорными и моторными системами мозгового ствола). Слуховые пути состоят из нервных проводников, соединяющих рецепторы слуха со слуховыми центрами всех уровней. Наряду с афферентными они содержат эфферентные нервные волокна, значение к-рых недостаточно выяснено. Кроме вертикально направленных пучков, в составе слуховых путей имеются горизонтальные волокна, соединяющие ядра одного уровня между собой.
Анатомия
Первый нейрон афферентного слухового пути представлен биполярными нейроцитами спирального узла улитки (см. Внутреннее ухо). Их периферические отростки направляются в спиральный орган улитки (кортиев орган), где оканчиваются у наружных и внутренних волосковых сенсорных клеток (см. Кортиев орган). Центральные отростки составляют улитковый (нижний) корешок преддверно-улиткового нерва (см.). Почти все они оканчиваются в улитковых ядрах (вентральном и дорсальном), лежащих в продолговатом мозге (см.) на границе с варолиевым мостом (мост головного мозга, Т.), соответственно преддверному полю (area vestibularis) ромбовидной ямки. В этих ядрах находятся тела 2-го нейрона слухового пути; единый путь разделяется здесь на две части. Вентральное (переднее) улитковое ядро [nucleus cochlearis ventralis (ant.)] филогенетически более старое, волокна из него идут поперечно через варолиев мост, образуя трапециевидное тело (corpus trapezoi-deum). Большинство волокон трапециевидного тела оканчивается в заложенных в нем передних (вентральных) и задних (дорсальных) ядрах (nuclei ventrales et dorsales corporis trapezoidei), а также в верхнем оливном ядре [nucleus olivaris cranialis (sup.)] своей и противоположной сторон и ядрах ретикулярной формации покрышки (nuclei tegmenti), остальные волокна продолжаются в латеральную петлю. Аксоны нейроцитов ядер трапециевидного тела и верхнего оливно-го ядра (третий нейрон) направляются в латеральную петлю своей и противоположной сторон и, кроме того, подходят к ядрам лицевого и отводящего нервов, ретикулярной формации и часть их вступает в задний продольный пучок (fasciculus Jongitudinalis post.). За счет этих связей могут осуществляться рефлекторные движения при звуковых раздражениях. Дорсальное (заднее) улитковое ядро [nucleus cochlearis dorsalis (post.)], филогенетически более молодое, дает начало волокнам, к-рые выходят на поверхность ромбовидной ямки в виде мозговых полосок (striae medullares), направляющихся к срединной борозде. Там они погружаются в вещество мозга и образуют два перекреста — поверхностный (Монакова) и глубокий (Гель-да), после чего вступают в латеральную петлю (lemniscus lat.). Последняя представляет главный восходящий слуховой путь мозгового ствола, объединяющий волокна из различных ядер слуховой системы (заднего улиткового, верхнего олив-ного ядер трапециевидного тела). Латеральная петля содержит как прямые, так и перекрещенные волокна; таким образом обеспечивается двусторонняя связь органа слуха с подкорковыми и корковыми слуховыми центрами. В латеральной петле лежит собственное ядро (nucleus lemnisci lat.), в к-ром переключается часть ее проводников.
Эфферентные волокна имеются во всех звеньях слуховых путей. От коры головного мозга идут две системы нисходящих проводников; более короткие оканчиваются в медиальном коленчатом теле и нижних холмиках, более длинные прослеживаются до верхнего оливного ядра. От последнего к улитке проходит оливоулитковый путь (tractus olivocochlearis Rasmussen), который содержит прямые и перекрещенные волокна. Те и другие достигают спирального органа улитки и оканчиваются на его наружных и внутренних волосковых клетках.
Патология
При поражении С. ц., п. развиваются нейросенсорные нарушения, к-рые разделяют на кохлеарные и ретрокохлеарные. Кохлеарные нарушения связаны с поражением нейрорецепторного аппарата в улитковом лабиринте внутреннего уха, а ретрокохлеарные — с поражением слухового нерва и его корешка, проводящих путей и центров.
При кохлеарном поражении слух (см.) страдает преимущественно на высокие тона, равномерно снижается восприятие звука как при воздушной, так и при костной проводимости. При этом кривые воздушной и костной проводимости на тональной аудиограмме располагаются параллельно, близко одна от другой. При исследовании костной проводимости с теменной области камертоном С128 (см. Вебера опыт) и ультразвуковым излучателем при 98 тыс. колебаний в 1 сек. звук в обоих случаях слышен на стороне, противоположной пораженной. На стороне поражения наблюдается повышенная чувствительность к нарастанию громкости звука (феномен ускоренного нарастания громкости бывает положительным), особенно в начальной стадии болезни Меньера (см. Меньера болезнь). При кохлеарном снижении слуха обычно отсутствуют какие-либо неврол. симптомы, но могут присоединяться вестибулярные нарушения вследствие вовлечения вестибулярного лабиринта (см. Вестибулярный симптомокомплекс) либо поражаться нервы, проходящие в височной кости (вкусовые волокна для передних 2/3 языка, лицевой нерв).
При кохлеарном поражении субъективные шумы очень сильны и мучительны для больного, локализуются в ухе; при шумовой аудиометрии (см.) слух снижается, но не резко; речетональная диссоциация выражена слабо; при наличии нескольких источников звука восприятие речи снижается незначительно; гиперакузия наблюдается в одном ухе, нередко на него снижен слух (при этом нет повышения чувствительности к раздражению в других анализаторах); нарушение локализации звука соответствует одностороннему снижению слуха; слуховых галлюцинаций не бывает.
При ретрокохлеарном поражении снижение слуха четко зависит от локализации цроцесса в мозге. Наиболее часто односторонняя глухота (см.) или одностороннее резкое снижение слуха отмечается при поражении в области мостомозжечкового угла (корешковое поражение),боковых отделов варолиева моста (ядер-ное поражение). При поражении среднего мозга возникает двустороннее снижение слуха.
Ретрокохлеарное снижение слуха характеризуется целым рядом особенностей. При нем наиболее часто и значительно снижается слух на высокие частоты (4000—8000 гц) параллельно по костной и воздушной проводимости. Поэтому по характеру тональных аудиограмм нельзя отличить кохлеарное снижение слуха от ретрокохлеарного.
При ретрокохлеарной односторонней тугоухости (см.) или глухоте отсутствует латерализация звука в опыте Вебера (см. Вебера опыт), что является основным признаком для дифференциации кохлеарного и ретрокохлеарного снижения слуха. В то же время ультразвук четко латерализуется в лучше слышащее ухо при любой форме нейросенсорной односторонней тугоухости. Характерна, особенно для поражения среднего мозга, речетональная диссоциация с преимущественным нарушением разборчивости речи. При нарастании интенсивности звука до 110 дб часто отмечается парадоксальное снижение разборчивости речи. Наличие нескольких источников звука, присоединение шума, частая подача звуковых сигналов резко снижают восприятие тонов и особенно речи. При центральном поражении многие особенности нарушения слуха связаны не столько с поражением слухового анализатора, сколько с изменением в. н. д.
При ретрокохлеарном поражении субъективные шумы у больных слабо выражены, локализуются в голове; при шумовой аудиометрии сильно снижается слух; речетональная диссоциация проявляется в очень резкой форме, причем, чем выше уровень поражения слуховых путей, тем выраженней проявляется этот симптом; гиперакузпя наблюдается в обоих ушах при нормальном слухе, что сопровождается часто повышенной чувствительностью в других анализаторах (гяперосмией, гиперестезией); при нормальном слухе нарушается пространственный слух на стороне, противоположной локализаций очага поражения в полушарии головного мозга.
При снижении слуха уровень поражения слуховых путей устанавливается при сопоставлении слуховых нарушений с данными обследования вестибулярной функции, вкуса и другими неврологическими симптомами.
Различные по характеру и локализации заболевания имеют нек-рые особенности в проявлении слуховых нарушений. Наиболее часто и сильно снижается слух при поражении мостомозжечкового угла, улитковых (слуховых) ядер в боковых отделах варолиева моста и при процессах в среднем мозге. Для поражения в области мостомозжечкового угла при опухоли преддверно-улиткового нерва (см. Мостомозжечковый угол, патология) характерно одностороннее снижение слуха, а нередко полная односторонняя глухота при длительной полной сохранности слуха на другое ухо. Арахноидит (см.) мостомозжечкового угла обычно сопровождается двусторонним снижением слуха преимущественно в диапазоне высоких частот (4000—8000 гц). При поражении в области мостомозжечкового угла обычно одновременно снижается или выпадает вкус на передних 2/3 языка, нарушается функция тройничного, отводящего и лицевого нервов, изменяется вестибулярная возбудимость, нередко имеется горизонтальный спонтанный нистагм (см.).
Поражение улитковых ядер при односторонних опухолях или латеральных инфарктах варолиева моста (см. Мост головного мозга) сопровождается односторонним резким снижением слуха или односторонней глухотой, сочетающейся с парезами и параличами взора в сторону опухоли, альтернирующими синдромами (см.), выраженным спонтанным нистагмом. Срединные опухоли варолиева моста обычно не вызывают снижения слуха.
Поражение среднего мозга (см.) протекает часто с резким двусторонним снижением слуха (иногда до полной глухоты), что может сочетаться с конвергирующим спонтанным нистагмом, выраженным повышением калорического нистагма, ослаблением или выпадением оптокинетического нистагма, нарушением зрачковых реакций (см. Зрачковые рефлексы), экстрапирамидными симптомами (см. Экстрапирамидная система).
При одностороннем поражении внутренней капсулы и височной доли головного мозга (см.) слух не снижается, т. к. слуховые пути расположены в полушариях мозга далеко друг от друга, и каждый слуховой путь в этих отделах имеет прямые и перекрещенные пути. В тех случаях, когда патол. очаг располагается в височной доле, возникают слуховые галлюцинации (см.), нарушается восприятие коротких звуковых сигналов, особенно снижается восприятие искаженной и ускоренной речи с выключением высоких тонов и речи с подачей различных слов в правое и левое ухо (дихотический слух); изменяется музыкальный слух. Патол. очаги в височно-теменных отделах мозга и нижней теменной дольке вызывают нарушения пространственного восприятия слуха на противоположной стороне (при нормальном слухе на оба уха). При больших опухолях височной доли головного мозга, вторично воздействующих на средний мозг, может отмечаться снижение слуха.
Наиболее часто снижение слуха наблюдается вследствие неврита преддверно-улиткового нерва, развивающегося после гриппа, острых респираторных заболеваний, эпидемического паротита, арахноидита с преимущественной локализацией в мостомозжечковом углу, цереброспинального менингита, применения антибиотиков, обладающих ототоксическим действием (неомицина, канамицина, мономицина, гентамицина, стрептомицина), а также фуросемида, при интоксикации свинцом, мышьяком, фосфором, ртутью, при длительном воздействии шума (у ткачей, молотобойцев и др.), при опухолях слухового нерва (улитковой части преддверно-улиткового нерва, Т.), переломах пирамиды височной кости, у больных с сосудистым, воспалительным или опухолевым поражением латеральных отделов варолиева моста.
В острой стадии неврита преддверно-улиткового нерва лечение включает внутривенное введение 40% р-ра гексаметилентетрамина (уротропина) с глюкозой, применение антибиотиков (за исключением ототоксических), прозерина, дибазола, компламина, стугерона, но-шпы или других сосудорасширяющих средств, витамина B1, 0,1% р-ра стрихнина нитрата в возрастающих дозах (от 0,2 до 1 мл), всего 20—30 инъекций, иглоукалывание, ингаляции карбогена, инъекции АТФ. Благоприятные результаты дает лечение, начатое в первые 3—5 дней от начала заболевания; лечение, начатое спустя 3 мес. от начала заболевания, малорезультативно. Лечение неврита преддверно-улиткового нерва, обусловленного применением ототоксических антибиотиков, малоэффективно; с целью профилактики неврита необходимо ограничивать их применение (только по строгим показаниям), не назначать одновременно и последовательно два разных ототоксических антибиотика, ограничивать их назначение детям и лицам пожилого возраста.
Лечение опухолей преддверно-улиткового нерва оперативное (см. Преддверно-улитковый нерв).
Восстановление слуха при энцефалитах, опухолевых и сосудистых поражениях головного мозга зависит от эффективности лечения основного заболевания.
Библиография: Благовещенская Н. С. Клиническая отоневрологии при поражениях головного мозга, М., 1976; она же, Отоневрологические симптомы и синдромы, М., 1981; Блинков С. М. и Глезер И. И. Мозг человека в цифрах и таблицах, Л., 1964, библиогр.; Богословская Л. С. и Солнцева Г. Н. Слуховая система млекопитающих, М., 1979; Гринштейн А. М. Пути и центры нервной системы, М., 1946; Зворыкин В. П. Проблема ведущей афферентации и количественная перестройка стволовых формаций слухового и зрительного анализаторов у хищных и приматов, включая чело-' века, Арх. анат.. гистол. и эмбриол., т. 60, № 3, с. 13, 1971, библиогр.; Понтов А. С. и др. Очерки по морфологии связей центральной нервной системы, Л., 1972; Склют И. А. и Слатвинская Р. Ф. Принципы ранней аудиологической диагностики неврином слухового нерва, Журн. ушн., нос. и горл, бол., ЛЬ 2, с. 15, 1979; Солдатов И. Б., Сущева Г. и Храппо Н. С. Вестибулярная дисфункция, М., 1980. библиогр.; Тугоухость, под ред. Н. А. Преображенского, М., 1978; Хечинашвили С. Н. Вопросы аудиологии, Тбилиси, 1978; Эделмен Д ж. и Маунткасл В. Разумный мозг, пер. с англ., М., 1981; С 1 а-г а М. Das Nervensystem des Menschen, Lpz., 1959; Johnson E. W. Auditory test results in 500 cases of acoustic neuroma, Arch. Otolaryng., v. 103, p. 152, 1977; Spillmann T. u. Fisch U. Die Friihdiagnose des Akustikusneurinomes, Akt. Neurol., Bd 6, S. 39, 1979.
H. С. Благовещенская; В. С. Сперанский (ан.).
Читайте также:
