
Строение синапса и основы химической передачи нервного импульса
Механизм химической передачи нервных импульсов через синапс сводится к тому, что содержимое синаптических пузырьков (медиатор) поступает в виде небольших порций (квантов) в синаптическую щель и взаимодействует затем с рецепторными белками постсинаптической мембраны. Это вызывает деполяризацию мембраны и возбуждение следующего нейрона. Ультраструктурные особенности синапса и механизм передачи импульсов определяют строгую однонаправленность передачи импульсов, что лежит в основе проведения импульсов по рефлекторным дугам.
В зависимости от того, какое вещество выполняет роль нейромедиатора, синапсы подразделяются на холинергические (медиатор — ацетилхолин), адренергические (адреналин и норадреналин), дофаминергические (дофамин), серотонинергические (серотонин), пептидергические (медиаторы — пептиды и аминокислоты, например, мет-энкефалин, гамма-аминомасляная кислота, глицин и др.).
Нейрохимические синапсы подразделяются в функциональном отношении на две противоположные по своему значению группы — возбуждающие и тормозные. Свойства этих синапсов зависят как от медиаторов, так и от ультраструктурных особенностей синапсов. Так, некоторые медиаторы (например, глутамат) характерны для возбуждающих синапсов, а в тормозных синапсах медиатором является гамма-аминомасляная кислота. Предполагают, что в возбуждающих синапсах к постсинаптической мембране прилежит электронно-плотное вещество, в связи с чем синапс приобретает асимметричное строение.
В пресинаптических частях таких соединений содержатся круглые синаптические пузырьки. Тормозные синапсы имеют симметричное строение. Синаптическая щель у них сужена и в пресинаптической части содержатся уплощенные синаптические пузырьки.
Кроме нейрохимических синапсов между нервными клетками (преимущественно между дендритами или телами нейронов) возникают электротонические синапсы. Последние у млекопитающих встречаются редко и по строению соответствуют щелевым контактам. Они проводят возбуждение благодаря формированию трансмембранных каналов — коннексонов.
Каждый нейрон на своей поверхности имеет огромное количество (до 10000) синапсов. Интеграция импульсов в пределах отдельного нейрона происходит так: от синапсов, расположенных на дендритах и теле, по его плазмолемме передается импульс к аксонному холмику (генераторному пункту нейрона), где путем суммирования всех возбуждающих и тормозных импульсов возникает результирующий потенциал действия.
Синаптические структуры обладают высокой чувствительностью к действию токсических факторов, в том числе психотропных отравляющих веществ. Их изменения играют важную роль в механизмах реактивности нервных тканей.
Рецепторные нервные окончания подразделяются на две группы: экстерорецепторы, воспринимающие раздражения из внешней среды, и интерорецепторы, служащие для восприятия раздражений из внутренней среды организма. В зависимости от специфичности раздражения, воспринимаемого рецептором, различают следующие чувствительные нервные окончания: механорецепторы, барорецепторы, хеморецепторы, терморецепторы, болевые рецепторы (ноцицепторы). Все рецепторы по особенностям их строения подразделяют на свободные и несвободные нервные окончания.
Первые из них состоят только из конечных разветвлений периферического отростка чувствительного нейрона; вторые — имеют в составе рецептора кроме нервного отростка глиальный компонент, а нередко и соединительнотканную капсулу. В последнем случае несвободные рецепторные окончания называют инкапсулированными. К числу таких рецепторов относятся пластинчатые, осязательные тельца, концевые колбы, мышечные веретена и др.
Эффекторные нервные окончания подразделяются на двигательные и секреторные. Двигательный, или моторный, эффектор передает нервные импульсы на рабочие органы и ткани. В скелетных мышцах они называются нервно-мышечными (аксомышечными) окончаниями. Терминальное разветвление аксона двигательного нейрона заканчивается булавовидным расширением. Оно соответствует пресинаптической части нервно-мышечного синапса, содержит скопление синаптических пузырьков и митохондрий, ограничено пресинаптической мембраной характерного строения.
Плазмолемма мышечного волокна в этой области образует многочисленные складки и углубления. В постсинаптической части волокна находится зернистая саркоплазма с большим количеством митохондрий и овальных ядер. Синаптическая щель имеет ширину 50-100 нм. У человека медиатором в нервно-мышечных синапсах является ацетилхолин. Моторные окончания в гладкой мышечной ткани, а также секреторные эффекторы имеют вид тонких пучков аксонов или их одиночных терминалей, прилежащих к клеткам иннервируемых тканей.
Возрастные изменения нервных тканей связаны с постепенным уменьшением запаса нервных клеток, особенно — чувствительных нейронов, а также снижением уровня метаболических процессов, что выражается в закономерном накоплении включений липофусцина ("пигмент изнашивания") в нейроплазме.
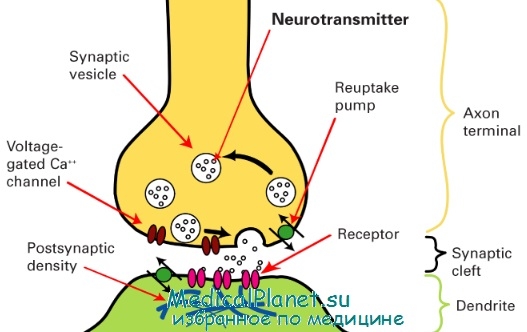
Схема строения синапса








- Физиология
![]()
- История физиологии
![]()
- Методы физиологии



Физиология и строение синапса
Возбуждение с нервного волокна на нервную, мышечную и железистую клетку передается посредством специального структурного образования — синапса.
Синапс — структура, обеспечивающая проведение сигнала от одной клетки к другой. Термин был введен английским физиологом Ч. Шеррингтоном в 1897 г.
Синапсы состоят из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 1).

Рис. 1. Строение синапса: 1 — микротрубочки; 2 — митохондрии; 3 — синаптические пузырьки с медиатором; 4 — пресинаптическая мембрана; 5 — постсинаптическая мембрана; 6 — рецепторы; 7 -синаптическая щель
Некоторые элементы синапсов могут иметь и другие названия. Например, синаптическая бляшка — это синапс между нейронами, концевая пластинка — постсинаптическая мембрана нервно-мышечного синапса, моторная бляшка — пресинаптическое окончание аксона на мышечном волокне.

Пресинаптическая мембрана покрывает расширенное нервное окончание, которое представляет собой нейросекреторный аппарат. В пресинаптической части находятся пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в гранулах (пузырьках).
Постсинаптическая мембрана - утолщенная часть мембраны клетки, с которой контактирует пресинаптическая мембрана. Она имеет ионные каналы и способна к генерации потенциала действия. Кроме того, на ней расположены специальные белковые структуры — рецепторы, воспринимающие действие медиаторов.
Синаптическая щель представляет собой пространство между пресинаптической и постсинаптической мембранами, заполненное жидкостью, близкой по составу к плазме крови.

Рис. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала.
По месту положения выделяют нервно-мышечные синапсы, нервно-железистые и нейро-нейрональные; последние, в свою очередь, делятся на аксо-аксональные, аксо-дендритические, аксо-соматические, дендро-соматические, дендро-дендротические.
По характеру действия на воспринимающую структуру синапсы могут быть возбуждающими и тормозящими.
По способу передачи сигнала синапсы делятся на электрические, химические, смешанные.
Таблица 1. Классификация и виды синапсов

Синапсы классифицируют следующим образом:
- по местоположению — периферические и центральные;
- по характеру их действия — возбуждающие и тормозящие;
- по способу передачи сигналов — химические, электрические, смешанные;
- по медиатору, с помощью которого осуществляется передача, — холинергические, адренергические, серотонинергические и т.д.
В химических синапсах возбуждение передается с помощью медиаторов (посредников).
Медиаторы — молекулы химических веществ, которые обеспечивают передачу возбуждения в синапсах. Другими словами химические вещества, участвующие в передаче возбуждения или торможения от одной возбудимой клетки к другой.
Свойства медиаторов
- Синтезируются в нейроне
- Накапливаются в окончании клетки
- Выделяются при появлении иона Са2+ в пресинаптическом окончании
- Оказывают специфическое действие на постсинаптическую мембрану
По химическому строению медиаторы можно подразделить на амины (норадреналин, дофамин, серотонин), аминокислоты (глицин, гамма-аминомасляная кислота) и полипептиды (эндорфины, энкефалины). Ацетилхолин известен в основном как возбуждающий медиатор и содержится в различных отделах ЦНС. Медиатор находится в пузырьках пресинаптического утолщения (синаптической бляшки). Медиатор синтезируется в клетках нейрона и может ресинтезироваться из метаболитов его расщепления в синаптической щели.
При возбуждении терминалей аксона происходит деполяризация мембраны синаптической бляшки, вызывающая поступление ионов кальция из внеклеточной среды внутрь нервного окончания через кальциевые каналы. Ионы кальция стимулируют перемещение синаптических пузырьков к пресинаптической мембране, их слияние с ней и последующий выход медиатора в синаптическую щель. После проникновения в щель медиатор диффундирует к постсинаптической мембране, содержащей на своей поверхности рецепторы. Взаимодействие медиатора с рецепторами вызывает открытие натриевых каналов, что способствует деполяризации постсинаптической мембраны и возникновению возбуждающего постсинаптического потенциала. В нервно-мышечном синапсе этот потенциал называется потенциалом концевой пластинки. Между деполяризованной постсинаптической мембраной и соседними с ней поляризованными участками этой же мембраны возникают местные токи, которые деполяризуют мембрану до критического уровня с последующей генерацией потенциала действия. Потенциал действия распространяется по всем мембранам, например, мышечного волокна и вызывает его сокращение.
Выделившийся в синаптическую щель медиатор связывается с рецепторами постсинаптической мембраны и подвергается расщеплению соответствующим ферментом. Так, холинэстераза разрушает медиатор ацетилхолин. После этого некоторое количество продуктов расщепления медиатора поступает в синаптическую бляшку, где из них снова ресинтезируется ацетилхолин.
В организме имеются не только возбуждающие, но и тормозные синапсы. По механизму передачи возбуждения они сходны с синапсами возбуждающего действия. В тормозных синапсах медиатор (например, гамма-аминомасляная кислота) связывается с рецепторами постсинаптической мембраны и способствует открытию в ней каналов для ионов хлора. При этом активизируется проникновение этих ионов внутрь клетки и развивается гиперполяризация постсинаптической мембраны, обусловливающая возникновение тормозного постсинаптического потенциала.
В настоящее время выяснено, что один медиатор может связываться с несколькими различными рецепторами и индуцировать различные реакции.
Химические синапсы
Синапсы с химической передачей возбуждения обладают определенными свойствами:
- возбуждение проводится в одном направлении, так как медиатор выделяется только из синаптической бляшки и взаимодействует с рецепторами на постсинаптической мембраны;
- распространение возбуждения через синапсы происходит медленнее, чем по нервному волокну (синаптическая задержка);
- передача возбуждения осуществляется с помощью специфических медиаторов;
- в синапсах изменяется ритм возбуждения;
- синапсы способны утомляться;
- синапсы обладают высокой чувствительностью к различным химическим веществам и гипоксии.
Одностороннее проведение сигнала. Сигнал передается только от пресинаптической мембраны к постсинаптической. Это вытекает из особенностей строения и свойств синаптических структур.
Замедленная передача сигнала. Обусловлена синаптической задержкой в передаче сигнала с одной клетки на другую. Задержка вызывается временными затратами на процессы выброса медиатора, его диффузии к постсинаптической мембране, связывания с рецепторами постсинаптической мембраны, деполяризации и преобразования постсинаптического потенциала в ПД (потенциал действия). Длительность синаптической задержки колеблется от 0,5 до 2 мс.
Способность к суммации эффекта от приходящих к синапсу сигналов. Такая суммация проявляется, если последующий сигнал приходит к синапсу через короткое время (1- 10 мс) после предыдущего. В таких случаях амплитуда ВПСП возрастает и на постсинаптическом нейроне может генерироваться большая частота ПД.
Трансформация ритма возбуждении. Частота нервных импульсов, приходящих к пресинаптической мембране, обычно не соответствует частоте ПД, генерируемых постсинаптическим нейроном. Исключение составляют синапсы, передающие возбуждение с нервного волокна на скелетную мышцу.
Низкая лабильность и высокая утомляемость синапсов. Синапсы могут проводить 50-100 нервных импульсов в секунду. Это в 5-10 раз меньше, чем максимальная частота ПД, которую могут воспроизводить нервные волокна при их электростимуляции. Если нервные волокна считаются практически неутомляемыми, то в синапсах утомление развивается весьма быстро. Это происходит из-за истощения запасов медиатора, энергетических ресурсов, развития стойкой деполяризации постсинаптической мембраны и т.д.
Высокая чувствительность синапсов к действию биологически активных веществ, лекарственных препаратов и ядов. Например, яд стрихнин блокирует функцию тормозных синапсов ЦНС, связываясь с рецепторами, чувствительными к медиатору глицину. Столбнячный токсин блокирует тормозные синапсы, нарушая выделение медиатора из пресинаптической терминали. В обоих случаях развиваются опасные для жизни организма явления. Примеры действия биологически активных веществ и ядов на передачу сигналов в нервно-мышечных синапсах рассмотрены выше.
Свойства облегчения и депрессии синоптической передачи. Облегчение синаптической передачи имеет место, когда нервные импульсы поступают к синапсу через короткое время (10-50 мс) друг за другом, т.е. достаточно часто. При этом в течение некоторого промежутка времени каждый последующий ПД, приходящий к пресинаптической мембране, вызывает увеличение содержания медиатора в синаптической щели, возрастание амплитуды ВПСП и увеличение эффективности синаптической передачи.
Одним из механизмов облегчения является накопление ионов Са 2 в пресинаптической терминали. Для удаления кальциевым насосом порции кальция, вошедшей в синаптическую терминаль при поступлении ПД, необходимо несколько десятков миллисекунд. Если в это время приходит новый потенциал действия, то новая порция кальция входит в терминаль и ее эффект на высвобождение нейромедиатора складывается с остаточным количеством кальция, которое кальциевый насос не успел удалить из нейроплазмы терминали.
Имеются и другие механизмы развития облегчения. Этот феномен в классических руководствах по физиологии называют также посттетанической потенциацией. Облегчение синаптической передачи имеет значение в функционировании механизмов памяти, для образования условных рефлексов и обучения. Облегчение передачи сигналов лежит в основе развития пластичности синапсов и улучшения их функций при частой активации.
Депрессия (угнетение) передачи сигналов в синапсах развивается при поступлении очень частых (для нервно-мышечного синапса более 100 Гц) нервных импульсов к пресинаптической мембране. В механизмах развития явления депрессии имеют значение истощение запасов медиатора в пресинаптической терминали, снижение чувствительности рецепторов постсинаптической мембраны к медиатору, развитие стойкой деполяризации постсинаптической мембраны, затрудняющих генерацию ПД на мембране постсинаптической клетки.
Электрические синапсы
Кроме синапсов с химической передачей возбуждения в организме есть синапсы с электрической передачей. Эти синапсы имеют очень узкую синаптическую щель и пониженное электрическое сопротивление между двумя мембранами. Благодаря наличию поперечных каналов между мембранами и низкому сопротивлению, электрический импульс легко проходит через мембраны. Электрические синапсы обычно характерны для однотипных клеток.
В результате воздействия раздражителя пресинаптический потенциал действия раздражает постсинаптическую мембрану, где возникает распространяющийся потенциал действия.
Электрические синапсы характеризуются большей скоростью проведения возбуждения по сравнению с химическими синапсами и низкой чувствительностью к воздействию химических веществ.
Электрические синапсы бывают с одно- и двусторонней передачей возбуждения.
В организме встречаются и электрические тормозные синапсы. Тормозное влияние развивается за счет действия тока, который вызывает гиперполяризацию постсинаптической мембраны.
В смешанных синапсах может происходить передача возбуждения с помощью как электрических импульсов, так и медиаторов.
Синапс – это определенная зона контакта отростков нервных клеток и остальных невозбудимых и возбудимых клеток, которые обеспечивают передачу информационного сигнала. Синапс морфологически образуется контактирующими мембранами 2-х клеток. Мембрана, относящаяся к отростку нервных клеток, зовется пресинаптической мембраной клетки, в которую поступает сигнал, второе ее название – постсинаптическая. Вместе с принадлежностью постсинаптической мембраны синапс может быть межнейрональным, нейромышечным и нейросекреторным. Слово синапс было введено в 1897 г. Чарльзом Шеррингтоном (англ. физиологом).

Что же такое синапс?
Синапс – это специальная структура, которая обеспечивает передачу от нервного волокна нервного импульса на другое нервное волокно или нервную клетку, а чтобы произошло воздействие на нервное волокно от рецепторной клетки (области соприкосновения друг с другом нервных клеток и другого нервного волокна), требуется две нервные клетки.
Синапс – это небольшой отдел в окончании нейрона. При его помощи идет передача информации от первого нейрона ко второму. Синапс находится в трех участках нервных клеток. Также синапсы находятся в том месте, где нервная клетка вступает в соединение с разными железами или мышцами организма.
Из чего состоит синапс
Строение синапса имеет простую схему. Он образуется из 3-х частей, в каждой из которых осуществляются определенные функции во время передачи информации. Тем самым такое строение синапса можно назвать подходящим для передачи нервного импульса. Непосредственно на процесс передачи информации воздействуют две главные клетки: воспринимающая и передающая. В конце аксона передающей клетки находится пресинаптическое окончание (начальная часть синапса). Оно может повлиять в клетке на запуск нейротрансмиттеров (это слово имеет несколько значений: медиаторы, посредники или нейромедиаторы) – определенные химические вещества, с помощью которых между 2-мя нейронами реализуется передача электрического сигнала.

Медиаторы синапса
Медиатор (от латинского Media - передатчик, посредник или середина). Такие медиаторы синапса очень важны в процессе передачи нервного импульса.
Морфологическое различие тормозного и возбуждающего синапса заключается в том, что они не имеют механизм освобождения медиатора. Медиатор в тормозном синапсе, мотонейроне и другом тормозном синапсе считается аминокислотой глицином. Но тормозной или возбуждающий характер синапса определяется не их медиаторами, а свойством постсинаптической мембраны. К примеру, ацетилхолин дает возбуждающее действие в нервно-мышечном синапсе терминалей (блуждающих нервов в миокарде).
Ацетилхолин служит возбуждающим медиатором в холинэргических синапсах (пресинаптическую мембрану в нем играет окончание спинного мозга мотонейрона), в синапсе на клетках Рэншоу, в пресинаптическом терминале потовых желез, мозгового вещества надпочеников, в синапсе кишечника и в ганглиях симпатической нервной системы. Ацетилхоли-нестеразу и ацетилхолин нашли также во фракции разных отделов мозга, иногда в большом количестве, но кроме холинэргического синапса на клетках Рэншоу пока не смогли идентифицировать остальные холинэргические синапсы. По словам ученых, медиаторная возбуждающая функция ацетилхолина в ЦНС весьма вероятна.

Когда завершают свою функцию медиаторы синапса, катехоламин поглощается пресинаптическим нервным окончанием, при этом включается трансмембранный транспорт. Во время поглощения медиаторов синапсы находятся под защитой от преждевременного истощения запаса на протяжении долгой и ритмичной работы.
Синапс: основные виды и функции
Лэнгли в 1892 году было предположено, что синаптическая передача у вегетативной ганглии млекопитающих не электрической природы, а химической. Через 10 лет Элиоттом было выяснено, что из надпочечников адреналин получается от того же воздействия, что и стимуляция симпатических нервов.
Синапс химический
Химический синапс принципиально отличается передачей раздражения при помощи медиатора с пресинапса на постсинапс. Поэтому и образуются различия в морфологии химического синапса. Химический синапс более распространен в позвоночной ЦНС. Теперь известно, что нейрон способен выделять и синтезировать пару медиаторов (сосуществующих медиаторов). Нейроны тоже имеют нейромедиаторную пластичность – способность изменять главный медиатор во время развития.

Нервно-мышечный синапс
Данный синапс осуществляет передачу возбуждения, однако эту связь могут разрушить различные факторы. Передача заканчивается во время блокады выбрасывания в синаптическую щель ацетилхолина, также и во время избытка его содержания в зоне постсинаптических мембран. Множество ядов и лекарственных препаратов влияют на захват, выход, который связан с холинорецепторами постсинаптической мембраны, тогда мышечный синапс блокирует передачу возбуждения. Организм гибнет во время удушья и остановки сокращения дыхательных мышц.
Синапс нервный: особенности и компоненты
Синапс – это соединение места контакта среди двух клеток. Причем каждая из них заключена в свою электрогенную мембрану. Нервный синапс состоит из трех главных компонентов: постсинаптическая мембрана, синаптическая щель и пресинаптическая мембрана. Постсинаптическая мембрана – это нервное окончание, которое проходит к мышце и опускается внутрь мышечной ткани. В пресинаптической области имеются везикулы – это замкнутые полости, имеющие медиатор. Они всегда находятся в движении.

Синаптическая щель среди пост- и пресинаптических мембран
В разных синапсах величина щели различна. Это пространство наполнено межклеточной жидкостью, в которой имеется медиатор. Постсинаптическая мембрана накрывает место контакта нервного окончания с иннервируемой клеткой в мионевральном синапсе. В определенных синапсах постсинаптическая мембрана создает складку, возрастает контактная площадь.
Дополнительные вещества, входящие в состав постсинаптической мембраны
В зоне постсинаптической мембраны присутствуют следующие вещества:
- Рецептор (холинорецептор в мионевральном синапсе).
- Липопротеин (обладает большой схожестью с ацетилхолином). У этого белка присутствует электрофильный конец и ионная головка. Головка поступает в синаптическую щель, происходит взаимодействие с катионовой головкой ацетилхолина. Из-за этого взаимодействия идет изменение постсинаптической мембраны, затем происходит деполяризация, и раскрываются потенциально зависимые Na-каналы. Деполяризация мембраны не считается самоподкрепляющим процессом;
- Градуален, его потенциал на постсинаптической мембране зависит от числа медиаторов, то есть потенциал характеризуется свойством местных возбуждений.
- Холинэстераза – считается белком, у которого имеется ферментная функция. По строению она схожа с холинорецептором и обладает похожими свойствами с ацетилхолином. Холинэстеразой разрушается ацетилхолин, вначале тот, который связан с холинорецептором. Под действием холинэстеразы холинорецептор убирает ацетилхолин, образуется реполяризация постсинаптической мембраны. Ацетилхолином расщепляется до уксусной кислоты и холина, необходимого для трофики мышечной ткани.
При помощи действующего транспорта выводится на пресинаптическую мембрану холин, он используется для синтеза нового медиатора. Под воздействием медиатора меняется проницаемость в постсинаптической мембране, а под холинэстеразой чувствительность и проницаемость возвращается к начальной величине. Хеморецепторы способны вступать во взаимодействие с новыми медиаторами.
Синапс представляет собой место функционального, а не физического контакта между нейронами; в нем происходит передача информации от одной клетки к другой. Обычно встречаются синапсы между концевыми веточками аксона одного нейрона и дендритами (аксодендритные синапсы) или телом (аксосоматические синапсы) другого нейрона. Число синапсов, как правило, очень велико, что обеспечивает большую площадь для передачи информации. Например, на дендритах и телах отдельных мотонейронов спинного мозга находится свыше 1000 синапсов. Некоторые клетки головного мозга могут иметь до 10000 синапсов (рис. 16.8).
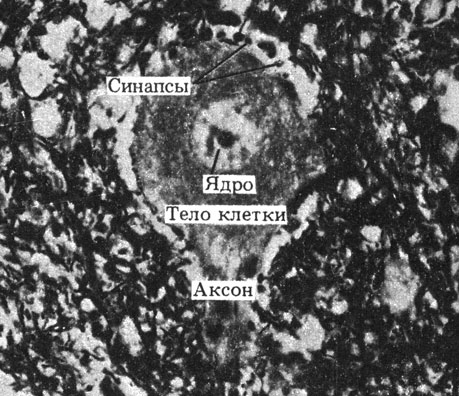
Рис. 16.8. Синапсы на двигательном нейроне (электронная микрофотография)
Существуют два типа синапсов - электрические и химические - в зависимости от природы проходящих через них сигналов. Между окончаниями двигательного нейрона и поверхностью мышечного волокна существует нервно-мышечное соединение, отличающееся по строению от межнейронных синапсов, но сходное с ними в функциональном отношении. Структурные и физиологические различия между обычным синапсом и нервно-мышечным соединением будут описаны несколько позже.
Химические синапсы - наиболее распространенный тип синапса у позвоночных. Это луковицеобразные утолщения нервных окончаний, называемые синаптическими бляшками и расположенные в непосредственной близости от окончания дендрита. Цитоплазма синаптической бляшки содержит митохондрии, гладкий эндоплазматический ретикулум, микрофиламенты и многочисленные синаптические пузырьки. Каждый пузырек имеет в диаметре около 50 нм и содержит медиатор - вещество, с помощью которого нервный сигнал передается через синапс. Мембрана синаптической бляшки в области самого синапса утолщена в результате уплотнения цитоплазмы и образует пресинаптическую мембрану. Мембрана дендрита в области синапса также утолщена и образует постсинаптическую мембрану. Эти мембраны разделены промежутком - синаптической щелью шириной около 20 нм. Пресинаптическая мембрана устроена таким образом, что к ней могут прикрепляться синаптические пузырьки и выделяться в синаптическую щель медиаторы. Постсинаптическая мембрана содержит крупные белковые молекулы, действующие как рецепторы медиаторов, и многочисленные каналы и поры (обычно закрытые), через которые в постсинаптический нейрон могут поступать ионы (см. рис. 16.10, А).
Синаптические пузырьки содержат медиатор, который образуется либо в теле нейрона (и попадает в синаптическую бляшку, пройдя через весь аксон), либо непосредственно в синаптической бляшке. В обоих случаях для синтеза медиатора нужны ферменты, образующиеся в теле клетки на рибосомах. В синаптической бляшке молекулы медиатора "упаковываются" в пузырьки, в которых они хранятся до момента высвобождения. Основные медиаторы нервной системы позвоночных - ацетилхолин и норадреналин, но существуют и другие медиаторы, которые будут рассмотрены позже.
Ацетилхолин - аммонийное производное, формула которого приведена на рис. 16.9. Это первый из известных медиаторов; в 1920 г. Отто Леви выделил его из окончаний парасимпатических нейронов блуждающего нерва в сердце лягушки (разд. 16.2). Структура норадреналина подробно рассматривается в разд. 16.6.6. Нейроны, высвобождающие ацетилхолин, называются холинэргическими, а высвобождающие норадреналин - адренэргическими.
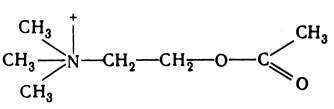
Рис. 16.9. Структурная формула ацетилхолина
Как полагают, прибытие нервного импульса в синаптическую бляшку вызывает деполяризацию пресинаптической мембраны и повышение ее проницаемости для ионов Са 2+ . Входящие в синаптическую бляшку ионы Са 2+ вызывают слияние синаптических пузырьков с пресинаптической мембраной и выход их содержимого из клетки (экзоцитоз), в результате чего оно попадает в синаптическую щель. Весь этот процесс называют электросекреторным сопряжением. После высвобождения медиатора материал пузырьков используется для образования новых пузырьков, заполняемых молекулами медиатора. Каждый пузырек содержит около 3000 молекул ацетилхолина.
Молекулы медиатора диффундируют через синаптическую щель (этот процесс занимает около 0,5 мс) и связываются с находящимися на постсинаптической мембране рецепторами, способными узнавать молекулярную структуру ацетилхолина. При связывании молекулы рецептора с медиатором ее конфигурация меняется, что приводит к открытию ионных каналов и поступлению в постсинаптическую клетку ионов, вызывающих деполяризацию или гиперполяризацию (рис. 16.4,А) ее мембраны в зависимости от природы высвобождаемого медиатора и строения молекулы рецептора. Молекулы медиатора, вызвавшие изменение проницаемости постсинаптической мембраны, сразу же удаляются из синаптической щели либо путем их реабсорбции пресинаптической мембраной, либо путем диффузии из щели или ферментативного гидролиза. В случае холинэргических синапсов находящийся в синаптической щели ацетилхолин гидролизуется ферментом ацетилхолинэстеразой, локализованным на постсинаптической мембране. В результате гидролиза образуется холин, он всасывается обратно в синаптическую бляшку и вновь превращается там в ацетилхолин, который хранится в пузырьках (рис. 16.10).
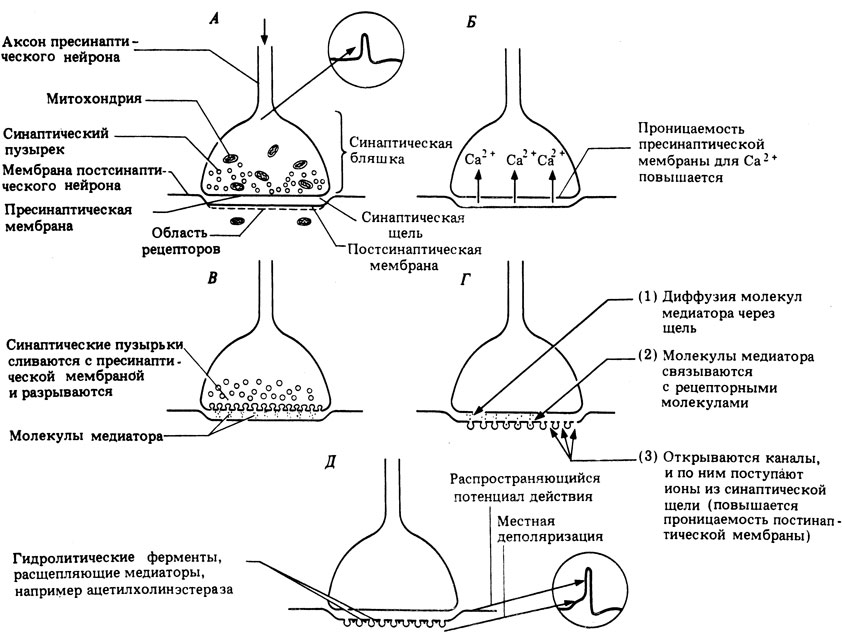
Рис. 16.10. Механизм химической передачи импульсов в нервном синапсе; от А до Д - последовательные этапы процесса
В возбуждающих синапсах под действием ацетилхолина открываются специфические натриевые и калиевые каналы, и ионы Na + входят в клетку, а ионы К + выходят из нее в соответствии с их концентрационными градиентами. В результате происходит деполяризация постсинаптической мембраны. Эту деполяризацию называют возбудительным постсинаптическим потенциалом (ВПСП). Амплитуда ВПСП обычно невелика, но продолжительность его больше, чем у потенциала действия. Амплитуда ВПСП меняется ступенчатым образом, и это позволяет предполагать, что медиатор освобождается порциями, или "квантами", а не в виде отдельных молекул. По-видимому, каждый квант соответствует освобождению медиатора из одного синаптического пузырька. Одиночный ВПСП не способен, как правило, вызвать деполяризацию пороговой величины, необходимой для возникновения потенциала действия. Но деполяризующие эффекты нескольких ВПСП складываются, и это явление носит название суммации. Два или больше ВПСП, возникших одновременно в разных синапсах одного и того же нейрона, могут сообща вызвать деполяризацию, достаточную для возбуждения потенциала действия в постсинаптическом нейроне. Это называют пространственной суммацией. Быстро повторяющееся высвобождение медиатора из пузырьков одной и той же синаптической бляшки под действием интенсивного стимула вызывает отдельные ВПСП, которые следуют так часто один за другим во времени, что их эффекты тоже суммируются и вызывают в постсинаптическом нейроне потенциал действия. Это называется временной суммацией. Таким образом, импульсы могут возникать в одиночном постсинаптическом нейроне либо как результат слабой стимуляции нескольких связанных с ним пресинаптических нейронов, либо как результат повторной стимуляции одного из его пресинаптических нейронов. В тормозных синапсах высвобождение медиатора повышает проницаемость постсинаптической мембраны за счет открытия специфических каналов для ионов К + и Сl - . Перемещаясь по концентрационным градиентам, эти ионы вызывают гиперполяризацию мембраны, называемую тормозным постсинаптическим потенциалом (ТПСП).
Медиаторы сами по себе не обладают возбуждающими или тормозящими свойствами. Например, ацетилхолин оказывает возбуждающее действие в большинстве нервно-мышечных соединений и других синапсов, но вызывает торможение в нервно-мышечных соединениях сердца и висцеральной мускулатуры. Эти противоположные эффекты обусловлены теми событиями, которые развертываются на постсинаптической мембране. От молекулярных свойств рецептора зависит, какие ионы будут входить в постсинаптический нейрон, а эти ионы в свою очередь определяют характер изменения постсинаптических потенциалов, как описано выше.
У многих животных, в том числе у кишечнополостных и позвоночных, передача импульсов через некоторые синапсы осуществляется путем прохождения электрического тока между пре- и постсинаптическими нейронами. Ширина щели между этими нейронами составляет всего лишь 2 нм, и суммарное сопротивление току со стороны мембран и жидкости, заполняющей щель, очень мало. Импульсы проходят через синапсы без задержки, и на их передачу не действуют лекарственные вещества или другие химические препараты.
Нервно-мышечное соединение представляет собой специализированный вид синапса между окончаниями двигательного нейрона (мотонейрона) и эндомизием мышечных волокон (разд. 17.4.2). Каждое мышечное волокно имеет специализированный участок - двигательную концевую пластинку, где аксон моторного нейрона (мотонейрона) разветвляется, образуя немиелинизированные веточки толщиной около 100 нм, проходящие в неглубоких желобках по поверхности мышечной мембраны. Мембрана мышечной клетки - сарколемма - образует множество глубоких складок, называемых постсинаптическими складками (рис. 16.11). Цитоплазма окончаний мотонейрона сходна с содержимым синаптической бляшки и во время стимуляции освобождает ацетилхолин с помощью того же механизма, о котором говорилось выше. Изменения конфигурации молекул - рецепторов, находящихся на поверхности сарколеммы, ведут к изменению ее проницаемости для Na + и К + , и в результате происходит местная деполяризация, называемая потенциалом концевой пластинки (ПКП). Эта деполяризация по величине вполне достаточна для возникновения потенциала действия, который распространяется по сарколемме в глубь волокна по системе поперечных трубочек (Т-системе) (разд. 17.4.7) и вызывает сокращение мышцы.

Рис. 16.11. Схема строения двигательной концевой пластинки и нервно-мышечного соединения. В верхней части рисунка (над линией Х-Х) изображено окончание аксона моторного нейрона на поверхности мышцы - двигательная концевая пластинка. В нижней части (под линией XX) показана ультраструктура нервно-мышечного соединения
Основная функция межнейронных синапсов и нервно-мышечных соединений состоит в передаче сигнала от рецепторов к эффекторам. Кроме того, строение и организация этих участков химической секреции обусловливают ряд важных особенностей проведения нервного импульса, которые можно резюмировать следующим образом:
1. Однонаправленность передачи. Высвобождение медиатора из пресинаптической мембраны и локализация рецепторов на постсинаптической мембране допускают передачу нервных сигналов по данному пути только в одном направлении, что обеспечивает надежность работы нервной системы.
2. Усиление. Каждый нервный импульс вызывает освобождение в нервно-мышечном синапсе достаточного количества ацетилхолина, чтобы вызвать распространяющийся ответ в мышечном волокне. Благодаря этому нервные импульсы, приходящие к нервно-мышечному соединению, как бы они ни были слабы, могут вызвать реакцию эффектора, и это повышает чувствительность системы.
3. Адаптация, или аккомодация. При непрерывной стимуляции количество освобождающегося в синапсе медиатора постепенно уменьшается до тех пор, пока запасы медиатора не будут истощены; тогда говорят, что синапс утомлен, и дальнейшая передача им сигналов тормозится. Адаптивное значение утомления состоит в том, что оно предотвращает повреждение эффектора вследствие перевозбуждения. Адаптация имеет место также на уровне рецепторов. (См. описание в разд. 16.4.2.)
4. Интеграция. Постсинаптический нейрон может получать сигналы от большого числа возбуждающих и тормозных пресинаптических нейронов (синаптическая конвергенция); при этом постсинаптический нейрон способен суммировать сигналы от всех пресинаптических нейронов. Благодаря пространственной суммации нейрон интегрирует сигналы, поступающие из многих источников, и выдает координированный ответ. В некоторых синапсах имеет место облегчение, состоящее в том, что после каждого стимула синапс становится более чувствительным к следующему стимулу. Поэтому следующие друг за другом слабые стимулы могут вызывать ответ, и это явление используется для повышения чувствительности определенных синапсов. Облегчение нельзя рассматривать как временную суммацию: здесь происходит химическое изменение постсинаптической мембраны, а не электрическая суммация постсинаптических мембранных потенциалов.
5. Дискриминация. Временная суммация в синапсе позволяет отфильтровывать слабые фоновые импульсы, прежде чем они достигнут мозга. Например, экстероцепторы кожи, глаз и ушей постоянно получают из окружающей среды сигналы, не имеющие особого значения для нервной системы: для нее важны лишь изменения интенсивности стимулов, приводящие к увеличению частоты импульсов, которое обеспечивает их передачу через синапс и надлежащую реакцию.
6. Торможение. Передача сигналов через синапсы и нервно-мышечные соединения может затормаживаться определенными блокирующими агентами, воздействующими на постсинаптическую мембрану (см. ниже). Возможно и пресинаптическое торможение, если на окончании аксона чуть выше данного синапса оканчивается другой аксон, образующий здесь тормозный синапс. При стимуляции такого тормозного синапса уменьшается число синаптических пузырьков, разряжающихся в первом, возбуждающем синапсе. Такое устройство позволяет изменять воздействие данного пресинаптического нейрона с помощью сигналов, приходящих от другого нейрона.
Химические вещества выполняют в нервной системе множество различных функций. Воздействия одних веществ широко распространены и хорошо изучены (как, например, возбуждающее действие ацетилхолина и адреналина), тогда как эффекты других носят локальный характер и пока еще недостаточно ясны. Некоторые вещества и их функции приведены в табл. 16.2.

Таблица 16.2. Вещества, влияющие на синапсы и нервно- мышечные соединения млекопитающих
Полагают, что некоторые лекарственные препараты, используемые при таких психических нарушениях, как тревожность и депрессия, воздействуют на химическую передачу в синапсах. Многие транквилизаторы и седативные средства (трициклический антидепрессант имипрамин, резерпин, ингибиторы моноаминоксидазы и др.) оказывают свой лечебный эффект, взаимодействуя с медиаторами, их рецепторами или отдельными ферментами. Так, например, ингибиторы моноаминоксидазы подавляют фермент, участвующий в расщеплении адреналина и норадреналина, и скорее всего оказывают свой лечебный эффект при депрессии, увеличивая продолжительность действия этих медиаторов. Галлюциногены типа диэтиламида лизерговой кислоты и мескалина, воспроизводят действие каких-то природных медиаторов мозга или же подавляют действие других медиаторов.
Проводившееся недавно изучение действия некоторых болеутоляющих веществ - опиатов героина и морфина - показало, что в мозгу млекопитающих присутствуют природные (эндогенные) вещества, вызывающие сходный эффект. Все эти вещества, взаимодействующие с опиатными рецепторами, получили общее название эндорфинов. К настоящему времени открыто много таких соединений; из них лучше всего изучена группа относительно небольших пептидов, называемых энкефалинами (мет-энкефалин, β-эндорфин и др.). Считается, что они подавляют болевые ощущения, влияют на эмоции и имеют отношение к некоторым психическим заболеваниям.
Все это открыло новые пути для изучения функций мозга и биохимических механизмов, лежащих в основе воздействия на боль и лечения с помощью таких различных методов, как внушение, гипно? и акупунктура. Предстоит выделить еще много других веществ типа эндорфинов, установить их строение и функции. С их помощью можно будет получить более полное представление о работе мозга, и это лишь вопрос времени, так как методы выделения и анализа веществ, присутствующих в столь малых количествах, непрерывно совершенствуются.
Читайте также:
