
Свойства нервных центров лекция
Нервный центр (НЦ) – объединение нейронов, координирующих и регулирующих в составе функциональной системы (П. К. Анохин) активность различных физиологических эффекторов, деятельность которых обеспечивает стабильность констант внутренней среды организма (рис.1). Например, дыхательный центр – объединение нейронов ствола мозга, регулирующих работу дыхательной мускулатуры для поддержания стабильности газовой константы организма; сердечно-сосудистый центр – объединение нейронов продолговатого мозга, регулирующих работу сердца, тонуса сосудов для обеспечения стабильности константы оптимального кровяного давления; терморегуляторный центр – нейронный комплекс гипоталамической области, координирующий и регулирующий работу множества эффекторов, активность которых поддерживает постоянство температурной константы организма.
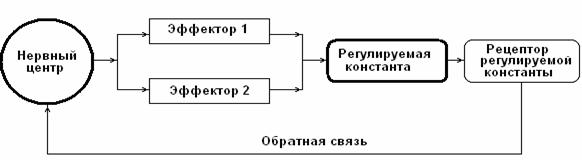
Рис. 1. Принципиальная схема функциональной системы (ФС) организма. В состав ФС входит нервный центр (НЦ), координирующий работу периферических эффекторов, деятельность которых определяет статус жизненно-важной константы. Качество координирующих и регулирующих влияний НЦ на эффекторы зависит от активности каналов обратной связи
Элементарной единицей НЦ является нейронный модуль – совокупность входных и вставочных нейронов, замыкающихся на один выходной (моторный), или конечный нейрон. НЦ обычно состоят из большого числа модулей, функционально связанных друг с другом и работающих параллельно друг с другом. На рис. 2 представлена схема простейшего НЦ, в состав которого входят 2 модуля. Каждый отдельно взятый нейронный модуль демонстрирует активность, характерную для всего НЦ.
Свойства нервных центров. Нервные центры (НЦ) обладают рядом характерных свойств, присущих всем нервным центрам. От чего зависят эти свойства?
1. От свойств одиночных нейронов, входящих в состав НЦ. 2. От свойств химических синапсов, обеспечивающих передачу влияний одних нейронов на другие. 3. От особенностей связей между отдельными нейронами в составе нейронного модуля. В простейшем варианте связи могут быть линейными, однако чаще в составе НЦ нейроны формируют сети сложной конфигурации.
Частные свойства нервных центров (НЦ).
1. НЦ проводит возбуждение только в одну сторону - от входных нейронов к выходным. Данное свойство НЦ определяется как односторонняя проводимость – от входных нейронов НЦ к выходным. Односторонняя проводимость НЦ объясняется тем, что химические синапсы (синапс 1, синапс 2 - рис. 3) проводят возбуждение только в одну сторону – от пресинаптической мембраны к мембране постсинаптической. Одностороннюю проводимость НЦ можно доказать в простом эксперименте с раздражением задних и передних корешков спинного мозга. Если раздражать электрическим током задние корешки, от передних корешков всегда можно зарегистрировать потенциалы действия. Это свидетельствует о проведении возбуждения через НЦ от входных нейронов к выходным. Если раздражать передние корешки, от задних потенциалы действия никогда не регистрируются. Это свидетельствует о том, что от выходных нейронов к входным возбуждение в НЦ не проводится.
2. Возбуждение в нервных центрах (НЦ) распространяется с медленной скоростью. Данное свойство определяется как замедленное распространение возбуждения в НЦ и в целом в ЦНС.
По периферическим нервным волокнам возбуждение распространяется быстро (по аксонам чувствительных нейронов возбуждение распространяется со скоростью 50-70 м/с). Однако, как только возбуждение поступает в НЦ скорость его распространения снижается. Это связано с феноменом синаптической задержки. Известно, что для проведения возбуждения через один химический синапс времени требуется примерно 2 мс (суммарная синаптическая задержка). Следовательно, если возбуждение в НЦ проходит несколько синапсов, средняя скорость проведения возбуждения в НЦ оказывается сниженной по сравнению со скоростью проведения возбуждения в нервных волокнах.

Рис. 2. Принципиальная схема простейшего нервного центра (НЦ), в состав которого входят два модуля (связи между ними – не показаны). Обращает на себя внимание тот факт, что в конечном итоге все входные возбуждения к НЦ сходятся (феномен конвергенции) на одном выходном нейроне модуля НЦ. Выходной нейрон, интегрируя (объединяя) входные потоки, формирует один выходной поток возбуждения, который поступает к периферическому эффектору.
Хорошо известно, что рефлекторные реакции, т. е. изменения активности периферических эффекторов в результате раздражения сенсорных рецепторов и проведения афферентного возбуждения через НЦ, возникают с некоторым скрытым (латентным) периодом. Латентный период – это время от момента нанесения раздражения до появления ответной реакции. Значительную часть латентного периода рефлекторных реакций составляет время проведения возбуждения через НЦ. Последнее определяется как центральное время рефлекторного акта.
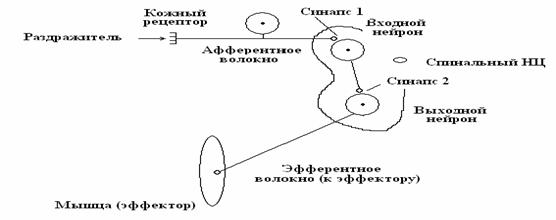
Рис. 3. Схема проведения афферентного возбуждения через спинальный НЦ к скелетной мышце – от входного нейрона к выходному, в обратном направлении возбуждение не проходит. Причина – химические синапсы (1, 2) обладают свойством односторонней проводимости.
3. Возбуждение в ЦНС может широко иррадиировать (перемещаться) от одного нервного центра (НЦ) к другим, часто расположенным всамых отдаленных зонах ЦНС.
Иррадиация возбуждения в ЦНС объясняется ветвлением аксонов выходных нейронов НЦ, связывающих соседние НЦ. При этом создаются возможности для распространения возбуждения по ЦНС (рис. 4). Беспрепятственную иррадиацию возбуждения в ЦНС ограничивают тормозные нейроны.

Рис. 4. Иррадиация возбуждения от НЦ 1 к другим НЦ в составе ЦНС. В целях упрощения нервные центры представлены лишь выходными нейронами.
Примеры макроорганизации иррадиации периферического возбуждения по коре головного мозга.
Горизонтальная иррадиация. Возбуждение от кожных болевых рецепторов поступает по спиноталамическому тракту, формируя первичный очаг в коре головного мозга. От места возникновения первичного очага возбуждение по горизонтальным ассоциативным связям может иррадиировать по коре головного мозга (рис. 5).
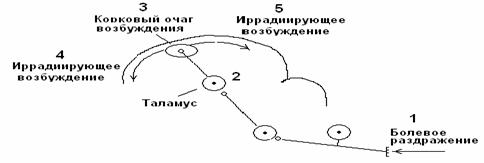
Рис. 5. Механизм горизонтальной иррадиации периферического возбуждения по коре головного мозга.
Вертикальная иррадиация. Возбуждение от кожных болевыхрецепторов по спиноталамическому тракту поступает в кору головного мозга и по стволовым коллатералям – в стволовую ретикулярную формацию. По многочисленным вертикальным ретикуло-кортикальным трактам возбуждение из ретикулярной формации вторично иррадиирует по коре головного мозга. Данный механизм иррадиации периферического возбуждения по коре головного мозга является основным (рис. 6).
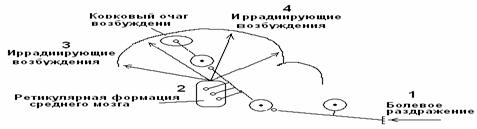
Рис. 6. Механизм вертикальной иррадиации периферического возбуждения по коре головного мозга.
Возвратная иррадиация. Первичное возбуждение в коре головного мозга по корково-ретикулярным связям может активировать нервные центры восходящей ретикулярной формации ствола мозга, при этом возбуждение из ретикулярной формации вторично перемещается в кору по множественным вертикальным ретикуло-корковым ткактам, иррадиируя по различным корковым центрам (рис. 7).

Рис. 7. Механизм возвратной иррадиации возбуждения по коре головного мозга.
На рис. 8 представлена нейронная схема НЦ, обеспечивающая режим усиления, или мультипликации (умножения) входного сигнала.
На вход НЦ поступает 2 нервных импульса, на выходе – регистрируем 6. Феномен усиления входной импульсации осуществляется в результате ее мультипликации (умножения) на нейронах НЦ, связанных в конечном итоге с нейроном выходным.

Рис. 8. Схема нейронной сети НЦ, работающей в режиме усилителя входного сигнала в результате его мультипликации (умножения) на нейронах 1, 2, 3.
На рис. 9 представлена нейронная сеть НЦ, обеспечивающая ослабление (редукцию) входного сигнала. Рассмотрим простейшую ситуацию. Линейная цепь А-В не выполняет преобразовательную работу. Добавим в нее один элемент – тормозный нейрон С, возбуждаемый выходными сигналами клетки В. Аксон тормозного нейрона соединен с выходным нейронов В по схеме возвратного торможения. Что наблюдается на выходе НЦ, если на входной нейрон А непрерывно будет поступать нервная импульсация ?

Рис. 9. Нейронная сеть нервного центра с тормозным нейроном, включенным по схеме возвратного торможения. В целом нервная сеть НЦ работает как аттенюатор – выходной поток нервных импульсов слабее входного.
Клетка А принимает поток входных импульсов, передавая их на нейрон В, который работает в паре с тормозным нейроном С. Что делает клетка В? Она отвечает на 1-й, 2-й, 3-й импульсы, поступающие к ней от нейрона А. Однако параллельно возбуждение идет к тормозному нейрону С. Нейрон С возбуждается, при этом тормозный медиатор в окончаниях аксона С тормозит активность нейрона В. Нейрон В перестает реагировать на импульсацию от нейрона А, следовательно, в структуре выходного потока клетки В появляется пауза. Однако, как только клетка В перестанет работать под влиянием нейрона С, последний сам перестает работать, т. к. к нему перестает поступать возбуждающая импульсация от нейрона В. Это немедленно восстанавливает возбудимость нейрона В, и он вновь начинает реагировать на импульсацию от нейрона А. Далее циклы повторяются, при этом выходной поток НЦ, определяемый активностью нейрона В, будет носит форму, в которой вспышки активности чередуются с тормозными паузами (рис. 9). В целом выходная импульсация НЦ будет редуцирована в сравнении с исходной.

Рис. 10. Схема нейронной сети НЦ с тормозным нейроном, включенным по схеме параллельного торможения.
Еще один пример с участием тормозного нейрона. Что будет на выходе НЦ, если тормозная клетка включается в нейронную сеть по схеме параллельного торможения?
Под влиянием начальной порции возбуждения выходной нейрон В генерирует несколько импульсов. Однако через коллатераль аксона А параллельно активируется тормозный нейрон С, который тормозит клетку В. Ее активность тормозится. В итоге – на выходе наблюдаем всего два импульса в сравнении с шестью ПД на входе (рис. 10).
5. Нервным центрам (НЦ) присуще явление последействия, под которым понимают повышенную активность нейронов НЦ на протяжении некоторого времени после прекращения поступления нервных импульсов на вход НЦ.
Как этот феномен объяснить с позиций нейрофизиологии? Линейная цепь нейронов А-В последействие не обеспечивает. Однако ситуация изменяется, когда между нейронами А и В встраивается замкнутая сеть вставочных нейронов 1, 2, 3 (рис. 11).

Рис. 11. Схема нейронной сети, обеспечивающей феномен последействия в НЦ за счет циркуляции (реверберации) нервных импульсов в цепи вставочных нейронов 1, 2, 3, замыкающихся на нейрон А. Цепь вставочных нейронов загружается импульсацией при поступлении к нейрону А входной (афферентной) импульсации (1). В дальнейшем (2) входная импульсация воспроизводится непрерывно в замкнутой цепи нейронов 1, 2, 3, далее нейрон А и выходной нейрон В.
6. Нервные центры обладают способностью суммировать возбуждения, поступающие к ним по афферентным каналам. Суммация может быть двух видов: временная и пространственная.
Временная, или частотная суммация (рис. 12). К нейрону (А) НЦ в результате воздействия на входной нерв подходят друг за другом два нервных импульса. Каждый из импульсов вызывает генерацию ВПСП, недостаточную для выведения мембранного потенциала нейрона до КУД (В). Однако если второй импульс приходит к нейрону на фоне развития ВПСП от первого, ВПСП от второго импульса, суммируясь с ВПСП от первого, смещает мембранный потенциал до КУД, нейрон НЦ при этом возбуждается. В рассмотренном примере суммация двух входных подпороговых воздействий к НЦ является производным фактора времени между входными воздействиями. При некотором оптимальном временном интервале два подпороговых воздействия могут суммироваться на нейронах НЦ.

Рис. 12. Временная (частотная) суммация входных воздействий на нейроне НЦ.
Пространственная суммация (рис. 13). К нейрону (А) по различным входным нервам поступают два нервных импульса. Каждый вызывает подпороговый ВПСП (В). Однако если нервные импульсы поступают одновременно – подпороговые ВПСП суммируются (С), при этом мембранный потенциал нейрона смещается до КУД, что приводит к возбуждению нейрона НЦ.

Рис. 13. Пространственная суммация входных воздействий на нейроне НЦ, когда два импульса подходят к нейрону одновременно по различных афферентным проводникам (феномен конвергенции).
7. Нервные центры (НЦ) обладают способностью перестраивать свою работу в зависимости от изменения активности сопряженных нервных центров, изменения качества афферентных воздействий на НЦ. Данное свойство НЦ определяется как пластичность.
Как доказать феномен пластичности НЦ? На рис. 14-1 представлена упрощенная схема моторных центров спинного мозга А и В, осуществляющих контроль за активностью периферических мышц конечностей А и В. Перережем эфферентные (моторные) нервы и соединим их по схеме – нерв А – с мышцами В, нерв В с – мышцами А (рис. 14-2). Спустя некоторое время функциональная целостность нервов восстанавливается, однако моторная активность подопытного животного после вмешательства грубо нарушается. Животное теряет способность нормально передвигаться. Тем не менее, после многочисленных попыток перемещения в среде обитания ситуация постепенно начинает улучшаться вплоть до полного восстановления в результате радикальной перестройки нейронной активности спинальных двигательных центров.


Рис. 14. Упрощенная схема моторных центров спинного мозга интактного животного (1) и животного после перекрестного сшивания эфферентных нервов (2).

Рис. 13. Кора головного мозга играет главную роль в обеспечении пластических реакций ЦНС после повреждений.
В рассмотренном примере пластические перестройки в работе спинальных нервных центров происходят преимущественно за счет пластических перестроек нейронной активности на уровне коры головного мозга. Именно нисходящие корковые возбуждения нового качества вносят решающий вклад в изменение работы спинальных нервных центров. Ведущую роль коры головного мозга в обеспечении пластических перестроек спинальных центров можно доказать в эксперименте. Перережьте нисходящие в спинной мозг корковые двигательные тракты у животного после восстановления двигательных функций – ситуация немедленно вернется к исходной.
8. Нервные центры (НЦ) обладают повышенной утомляемостью. Известно, что нервное волокно практически не утомляется (Н. Е. Введенский), напротив, НЦ обладают повышенной утомляемостью. Неслучайно в целом организме утомление раньше всего развивается в НЦ (И. М. Сеченов).
9. Нервные центры обладают повышенной чувствительностью к ядам, биологически активным веществам, лекарственным препаратам. Это связано с обилием синаптических контактов в структуре нейронных сетей НЦ, наличием множества химических механизмов передачи возбуждения через синапсы.
10. Нервные центры обладают повышенной чувствительностью к недостатку кислорода.
11. Нервные центры (НЦ) могут пребывать в трех функциональных состояниях, отражающих совокупный уровень активности всех нейронов НЦ. Исходное состояние НЦ – нормальная (обычная) активность нейронов НЦ; второе состояние – сниженная активность нейронов НЦ; третье состояние – повышенная активность нейронов НЦ. В последнем случае НЦ в составе ЦНС становится доминантным, или господствующим (А. А. Ухтомский).
1. Гормональные причины, например, избыток или недостаток в циркулирующей крови тех или иных гормонов. 2. Патологические причины, например, хроническая болевая импульсация, формирующая доминантное состояние центров, ответственных за переработку болевой импульсации (ядра ретикулярной формации, гипоталамуса, таламуса, лимбической системы). 3. Метаболические причины, например, недостаток воды в организме, формирующий доминантное состояние гипоталамического центра жажды. 4. Информационные причины (применительно к человеку), связанные с восприятием словесных сигналов, письменных сообщений и проч., формирующих доминантное состояние корковых центров, имеющих первостепенное значение для организации целенаправленных поведенческих актов.
Нервный центр – это совокупность нейронов, обеспечивающих регуляцию какого-либо конкретного физиологического процесса или функции.
Нервный центр в узком смысле – это совокупность нейронов, без которых данная конкретная функция не может регулироваться. Например, без нейронов дыхательного центра продолговатого мозга дыхание прекращается. Нервный центр в широком смысле — это совокупность нейронов, которые участвуют в регуляции конкретной физиологической функции, но не являются строго обязательными для ее осуществления! Например, в регуляции дыхания кроме нейронов продолговатого мозга участвуют нейроны пневмотаксического центра варолиевого моста, отдельные ядра гипоталамуса, кора больших полушарий и другие образования головного мозга.
Все нейроны нервного центра разделяют на 2 неравные по количеству и качеству группы.

Первая группа – нейроны центральной зоны. Это наиболее возбудимые нейроны, которые возбуждаются в ответ на поступление порогового (для нервного центра) сигнала. Таких нейронов около 15-20%, и они не обязательно располагаются в середине нервного центра, как это изображено на рис.1. Особенностью их является то, что они имеют на своем теле больше синаптических терминалей от сенсорных и вставочных нейронов.
Вторая группа – нейроны подпороговой каймы. Это менее возбудимые нейроны, которые не возбуждаются в ответ на поступление пороговых им-пульсов, но при действии более сильных раздражителей они возбуждаются и включаются в работу нервного центра, обеспечивая ее усиление. Таких нейронов большинство (80-85%), и они не обязательно располагаются на периферии нервного центра, но все имеют значительно меньше синаптических терминалей от сенсорных и вставочных нейронов по сравнению с нейронами центральной зоны.
На рис. 1 нейроны центральной зоны условно поставлены в центр внутреннего круга (А), а нейроны подпороговой каймы – в пространство между внутренним и наружным кругами (Б). Таким образом, если к нервному центру по афферентному входу (В) придет пороговый импульс, то возбудятся три нейрона центральной зоны, а на десяти нейронах подпороговой каймы потенциалы действия не возникнут, но появится местная деполяризация – возбуждающий постсинаптический потенциал (ВПСП).
От структуры нервного центра зависят его свойства, а они, в свою очередь, влияют на процесс проведения возбуждения через нервный центр, на его скорость и степень выраженности. От свойств нервных центров во многом зависит процесс распространения возбуждения по ЦНС, что имеет важное значение в интегративной деятельности организма.
Свойства нервных центров обусловлены описанной выше нейронной организацией нервного центра, а также химическим способом передачи возбуждения в синапсах. При электрическом способе передачи возбуждения нервные центры не имели бы подобных свойств.
Свойства нервных центров: 1 одностороннее проведение возбуждения; 2 задержка проведения возбуждения; 3 суммация; 4 облегчение; 5 окклюзия; 6 мультипликация; 7 трансформация; 8 последействие; 9 посттетаническая потенциация; 10 утомление; 11 тонус; 12 высокая чувствительность к изменению состояния внутренней среды организма; 13 пластичность.
3) Суммация – это возникновение возбуждения в нервном центре при поступлении к нему нескольких допороговых импульсов, каждый из которых в отдельности не может возбуждения (рис. 2). Фактически этот процесс происходит на нейронах подпороговой каймы. Различают два вида суммации: пространственную и временною.

Пространственная суммация возникает в том случае, когда к нервному центру (к его нейронам) приходят одновременно, несколько допороговых импульсов. На рисунке 2А видно, что к нейрону подпороговой каймы, имеющему пороговый потенциал 30 мВ одновременно по пяти различным афферентным входам (их аксоны обозначены сплошной линией) приходят пять импульсов, каждый из которых деполяризует мембрану нейрона на 5 мВ (то есть возникают пять отдельных ВПСП). В этом случае возбуждение нейрона не наступает, так как суммарная деполяризация мембраны нейрона составляет лишь 25 мВ (суммированный ВПСП мал для достижения КУД). Но если к нейрону придет еще один подобный импульс по шестому входу (его аксон обозначен пунктирной линией), то суммированный ВПСП будет достаточен по величине и мембрана нейрона в зоне аксонного холмика деполяризуется до критического уровня, в результате чего нейрон из состояния покоя перейдет в состояние возбуждения. На постсинаптической мембране происходит суммация ВПСП в пространстве.
Временная (последовательная) суммация возникает в том случае, когда к нейронам нервного центра по одному афферентному входу приходит не один, а серия импульсов с очень небольшими по времени межимпульсными промежутками (рис. 2Б). Два механизма временной суммации:
1) интервалы между отдельными импульсами настолько малы, что за это время медиатор, выделившийся в синаптическую щель, не успевает полностью разрушиться и вернуться в пресинаптический аппарат. В этом случае возникает постепенное накопление медиатора до критического объема, необходимого для возникновения достаточного по амплитуде ВПСП, а значит, и для возникновения ПД;

2) интервалы между отдельными импульсами настолько малы, что возникший за это время на постсинаптической мембране ВПСП не успевает исчезнуть и усиливается за счет новой порции медиатора – суммируется. На постсинаптической мембране происходит суммация ВПСП во времени.
4) Облегчение – это увеличение количества возбужденных нейронов в нервном центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному, а по двум или более афферентным входам. На рис. 3 рассмотрен случай, когда при отдельном раздражении первого афферентного входа возбуждается только три нейрона центральной зоны (А), а на пяти нейронах подпороговой каймы (Б) возникают ВПСП. Если раздражать отдельно только второй афферентный вход, то возбуждены будут пять нейронов (Г), а четыре нейрона подпороговой каймы (Д) не возбудятся. Раздражая и первый, и второй афферентные входы одновременно (!), мы ожидаем вовлечения в процесс возбуждения восьми нейронов. И они, естественно, будут возбуждаться, но кроме них (сверх ожидания!) могут возбуждаться еще некоторые нейроны подпороговой каймы. Это произойдет потому, что один или несколько нейронов подпороговой каймы являются общими как для первого, так и для второго афферентных входов (в нашем случае это два нейрона, обозначенные буквой В), и при одновременном поступлении возбуждения к этим нейронам дни возбудятся за счет возникновения пространственной суммации.

5) Окклюзия – это уменьшение количества возбужденных нейронов в нервной центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному. а по двум или более афферентным входам (рис. 4).
6) Мультипликационное возбуждение (мультипликация) заключается в том, что по разветвлениям аксона вставочного нейрона возбуждение поступает одновременно не на один, а на несколько моторных нейронов (рис. 6). В связи с этим эффект на рабочем органе усиливается в несколько раз, или в работу вовлекаются не одна, а несколько рабочих структур, Это свойство особенно ярко проявляется в ганглиях автономной (вегетативной) нервной системы.

7) Трансформация ритма возбуждения – это изменение частоты импульсов на выходе из нервного центра по сравнению с частотой импульсов на входе в нервный центр.
а) наличием дублирующих цепей вставочных нейронов, связывающих сенсорные и моторные нейроны;
б) разным количеством синапсов в каждой из этих цепей.


Например, на рис.7 представлены два варианта трансформации, которые, на первый взгляд, не отличаются друг от друга, так как в том и в другом случае показаны две дополнительные цепи вставочных нейронов (кроме прямого пути), с помощью которых возбуждение может передаваться по цепи нейронов А—Б—В. Рассмотрим эти схемы.
Вариант 1. Верхняя цепь состоит из двух дополнительных вставочных нейронов, а значит, по сравнению с прямым путем передачи возбуждения с нейрона Б на нейрон В, имеет два дополнительных синапса. Поэтому возбуждение, проходя по верхней цепи, задержится на 2 мс (время синаптической задержки в одном синапсе составляет
1 мс) и придет на нейрон В после того, как пройдет возбуждение по прямому пути. В нижней цепи три дополнительных вставочных нейрона (то есть три дополнительных синапса), значит, возбуждение будет доходить до нейрона В еще дольше, чем по верхней цепи (задержка составит 3 мс). Следовательно, по нижней цепи возбуждение на нейрон В придет после того, как пройдет возбуждение по верхней цепи. В результате на один импульс, пришедший по сенсорному нейрону А, на моторном нейроне В возникнет три потенциала действия (трансформация 1:3).
Вариант 2. В этом случае и верхняя и нижняя цепи вставочных нейронов состоят из двух дополнительных нейронов. Возбуждение по обеим цепям придет к нейрону В одновременно в виде одного потенциала действие, который появится на нейроне В только после прохождения возбуждения к нему от нейрона Б по прямому пути. В этом варианте мы тоже получим трансформацию ритма, но уже в соотношении 1:2.
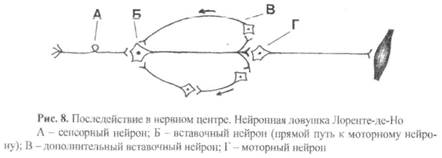
8) Последействие – это продолжение возбуждения моторного нейрона в течение некоторого времени после прекращения действия раздражителя.
Другими причинами последействия могут быть:
а) возникновение высокоамплитудного ВПСП, в результате которого возникает не один, а несколько потенциалов действия то есть ответ длится большее время;
б) длительная следовая деполяризация постсинаптической мембраны, в результате чего возникают несколько потенциалов действия, вместо одного.

9) Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей.
Если в качестве контроля вызвать одиночное раздражение афферентного нерва тестирующим раздражителем (рис. 9А), то на моторном нейроне мы получим ВПСП вполне определенной амплитуды (в нашем случае 5 мВ). Если после этого тот же афферентный нерв раздражать некоторое время серией частых импульсов (рис. 9Б), а потом вновь подействовать тестирующим раздражителем (рис. 9В), то величина ВПСП будет больше (в нашем случае 10 мВ). Причем она будет тем больше, чем более частыми импульсами мы раздражали афферентный нерв.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения: после одиночных стимулов оно выражено слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. Объясняется он тем, что при частом раздражении афферентного волокна в его пресинаптической терминали (окончании) накапливаются ионы кальция, а значит, улучшается выделение медиатора. Кроме того, показано, что частое раздражение нерва приводит к усилению синтеза медиатора, мобилизации пузырьков медиатора, к усилению синтеза белков-рецепторов на постсинаптической мембране и увеличению их чувтствительности. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах.
10) Утомление нервного центра (посттетаническая депрессия, синаптическая депрессия) – это уменьшение или прекращение импульсной активности нервного центра в результате длительной стимуляции его афферентными импульсами (или произвольного вовлечения его в процесс возбуждения посредством импульсов, идущих из коры больших полушарий). Причинами утомления нервного центра могут быть:
• истощение запасов медиатора в афферентном или вставочном нейроне;
• снижение возбудимости постсинаптической мембраны (то есть мембраны моторного или вставочного нейрона) из-за накопления, например, продуктов метаболизма.
Утомляемость нервных центров продемонстрировал Н.Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. tibialis и п. peroneus. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. Синаптическая депрессия при длительной активации центра выражается в снижении постсинаптических потенциалов.
11) Тонус нервного центра – это длительное, умеренное возбуждение нервного центра без видимо утомления Причинами тонуса могут быть:
• потоки афферентных импульсов, постоянно поступающие с неадаптирующихся рецепторов;
• гуморальные факторы, постоянно присутствующие в плазме крови;
• спонтанная биоэлектрическая активность нейронов (автоматия);
• циркуляция (реверберация) импульсов в ЦНС.
12) Нервный центр состоит из нейронов, а они очень чувствительны к изменению состава внутренней среды организма, что и отражается на свойствах нервных центров. Наиболее важными факторами, влияющими на работу нервных центров, являются: гипоксия; недостаток питательных веществ (например, глюкозы); изменение температуры; воздействие продуктов метаболизма; воздействие различных токсических и фармакологических препаратов.
Разные нервные центры имеют неодинаковую чувствительность к воздействию названных факторов. Так, нейроны коры больших полушарий наиболее чувствительны к гипоксии, недостатку глюкозы, продуктам метаболизма; клетки гипоталамуса – к изменению температуры, содержанию глюкозы, аминокислот, жирных кислот и др.; различные участки ретикулярной формации выключаются разными фармакологическими препаратами, различные нервные центры избирательно активируются или тормозятся разными медиаторами.
13) Пластичность нервного центра означает его способность изменять при определенных обстоятельствах свои функциональные свойства. В основе этого явления лежит поливалентность нейронов нервных центров. Особенно ярко проявляется это свойство при всевозможных повреждениях ЦНС, когда организм компенсирует утраченные функции за счет сохранившихся нервных центров. Особенно хорошо свойство пластичности выражено в коре больших полушарий. Например, центральные параличи, связанные с патологией двигательных центров коры, иногда полностью компенсируются, и ранее утраченные двигательные функции восстанавливаются.
Читайте также:
