
Свойство нервных центров посттетаническая потенциация
Нервным центром называется совокупность нейронов в различных отделах ЦНС, обеспечивающих регуляцию какой-либо функции организма. Например, бульбарный дыхательный центр.
Для проведения возбуждения через нервные центры характерны следующие особенности:
1. Одностороннее проведение. Оно идет от афферентного, через вставочный, к эфферентному нейрону. Это обусловлено наличием межнейронных синапсов.
2. Центральная задержка проведение возбуждения, т.е. по ЦНС возбуждение идет значительно медленнее, чем по нервному волокну. Это объясняется синаптической задержкой. Так как больше всего синапсов в центральном звене рефлекторной дуги, там скорость проведения наименьшая. Исходя из этого время рефлекса – это время от начала воздействия раздражителя до появления ответной реакции. Чем длительнее центральная задержка, тем больше время рефлекса. Вместе с тем оно зависит от силы раздражителя. Чем оно больше, тем время рефлекса короче и наоборот. Это объясняется явлением суммации возбуждений в синапсах. Кроме того, оно определяется и функциональным состоянием ЦНС. Например, при утомлении нервного центра длительность рефлекторной реакции увеличивается.
3. Пространственная и временная суммация. Временная суммация возникает как в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда возбуждающего постсинаптического потенциала. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторных нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются, и в мембране нейрона генерируется распространяющийся потенциал действия.
4. Трансформация ритма возбуждения – изменение частоты нервных импульсов при прохождении через нервный центр. Частота может снижаться или повышаться. Например, повышающая трансформация – увеличение частоты обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне. Второе – генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких возбуждающих постсинаптических потенциалов и возникновением одного потенциала действия в нейроне.
5. Посттетаническая потенциация – это усиление рефлекторной реакции в результате двигательного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы, выделяется большое количество нейромедиаторов в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
6. Последействие – это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
7. Тонус нервных центров – состояние постоянной повышенной активности. Он обусловлен постоянным поступлением к нервному центру нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например, проявлением тонуса соответствующих центров является тонус определенной группы мышц.
8. Автоматия (спонтанная активность) нервных центров. Периодическая или постоянная генерация нейронами нервных импульсов, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов.
9. Пластичность нервных центров. Это их способность изменять функциональные свойства. При этом центр приобретает возможность выполнять новые функции или восстанавливать старые после повреждения. В основе пластичности нервного центра лежит пластичность синапсов и мембран нейронов, которые могут изменять свою молекулярную структуру.
10. Низкая физиологическая лабильность и быстрая утомляемость. Нервные центры могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов, истощение состава медиаторов, длительность их синтеза.
СИСТЕМЫ ОБЛУЧЕНИЯ
ИСТОЧНИКИ ОБЛУЧЕНИЯ
ПРИМЕНЕНИЕ ИСКУССТВЕННЫХ ИСТОЧНИКОВ УФ-ИЗЛУЧЕНИЯ
7 октября 1965 г. Министерство здравоохранения СССР утвердило "Указания по профилактике светового голодания у людей". Севернее 60° - с 1 октября по 1 апреля. Средние широты (50-60°) – с 1 ноября по 1 апреля. Южные широты (45-50°) – с 1 декабря по 1 марта, Применяют 3 варианта облучения людей:
а) эритемными дозами;
б) закаливающими дозами, которые по действию мягче, но постепенно возрастают по количественному значению. Организм имеет возможность хорошо компенсировать воздействие Уф-лучей;
в) витаминные дозы — небольшие по количественному выражению. Большая биологическая активность Уф-лучей выдвигает необходимость точной дозировки. Минимальное количество УФ-излучения — 1/8-1/10 эритемной дозы в сутки. Из практики известно, что оптимум лежит в пределах 1/3-1/6 эритемной дозы. Величина оптимальной дозы постепенно повышается по мере привыкания человека к Уф-лучам и достигает 1/2 -1 биодозы при длительном облучении.
1. БУВ 15 и 30 (ЛБ-30-1) максимальное излучение 254 нм.
2. ЭУВ 15 и 30 (ЛЭ-30-1) максимальное излучение 313 нм.
3. ПРК 2, 4, 7 (375,220,1000 вт) максимальное излучение в области А.
4. ДКсТ — безбалластные дуговые трубчатые ксеноновые лампы, мощностью от 2 до 100 кВт. Они могут применяться в больших спортзалах,
1. Светооблучательные установки длительного действия (ЭУВ, ДКсТ).
2. фотарии (ЭУВ и ПРК) маячного, кабинного и лабиринтного типов. Антирахитический эффект можно получить, если облучать 600 см 2 поверхности кожи 1/8-1/10 эритем-ной дозы (лицо, руки). В фотариях облучению подвергается сразу 8-16000 см2 поверхности кожи с начальной дозировкой не менее 1/2 биодозы.
Ритмическая активация синапса часто сопровождается значительным увеличением амплитуды синаптических потенциалов, что выражается в увеличении амплитуды, например, сокращения мышцы. Посттетаническая потенциация – это усиление рефлекторного ответа после тетанических раздражений. Длительность посттетанической потенциации может составлять от нескольких минут до нескольких часов. С функциональной точки зрения посттетаническая потенциация представляет собой процесс облегчения в ЦНС, связанный с приобретением опыта, т.е. процесс научения, памяти.
Механизм:
1). Ритмическая активация мембраны пресинаптического окончания аксона ведет к увеличению амплитуды ПД, что вызывает высвобождение большего количества медиатора в синаптическую щель.
2). Ритмическая активация сопровождается увеличением запаса доступного медиатора, готового к выделению.
3). Ритмическая активация приводит к вхождению и накоплению Са 2+ в пресинаптическом окончании(мы уже с вами об этом упоминали при рассмотрении работы химического синапса), что увеличивает высвобождение медиатора в синаптическую щель. Это самый важный фактор для посттетанической потенциации.
Большее количество медиатора – больше SВПСП – больше амплитуда ПД – больше амплитуда сокращения мышцы.
Нервным центром (НЦ) называется совокупность нейронов в различных отделах ЦНС, обеспечивающих регуляцию какой-либо функции организма. Например, бульбарный дыхательный центр.
Для проведения возбуждения через нервные центры характерны следующие, особенности:
1. однострочное проведение, оно идет от афферентного, через вставочный к эфферентному нейрону. Это обусловлено наличием межнейронных синапсов.
2.Центральная задержка проведения возбуждения т.е по НЦ возбуждения идет значительно медленнее, чем по нервному волокну. Это объясняется синаптической задержкой т.к больше всего синапсов в центральном звене рефлекторной дуги, там скорость проведения наименьшая. Чем длительнее центральная задержка, тем больше время рефлекса. Вместе с тем оно зависит от силы раздражителя. Чем она больше, тем время рефлекса короче и наоборот. Эго объясняется явлением суммации возбуждений в синапсах. Кроме того, оно определяется и функциональным состоянием ЦНС. Например, при утомлении НЦ длительность рефлекторной реакции увеличивается.
3. Пространственная и временная суммация. Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда ВПСП. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторов нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются 11 и мембране нейрона генерируется распространяющийся ПД.
4. Трансформация ритма возбуждения - изменение частоты нервных импульсов при прохождении через нервный центр. Частота может понижаться или повышаться.. Понижающая трансформация объясняется суммацией нескольких ВПСП и возникновением одного ПД в нейроне.
5. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения нейронов центра.
6. Последействие- это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
7. Тонус нервных центров - состояние постоянной повышенной активности. Он обусловлен постоянным поступлением к НЦ нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например, проявлением тонуса соответствующих центров является тонус определенной группы мышц.
8. Автоматия или спонтанная активность нервных центров. Периодическая или постоянная генерация нейронами нервных импульсов, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессор метаболизма в нейронах и действием на них гуморальных факторов.
9. Пластичность нервных центров. Это их способность изменять функциональные свойства. При этом центр приобретает возможность выполнять новые функции или восстанавливать старые после повреждения. В основе пластичности Н.Ц. лежит пластичность синапсов и мембран нейронов, которые могут изменять свою молекулярную структуру.
10. Низкая физиологическая лабильность и быстрая утомляемость. Н.Ц. могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов.
Центральное торможение (сеченовское, постсинаптическое, пресинапртическое, пессимальное). Механизмы центрального торможения. Физиологическое значение прцесса торможения.
Явление центрального торможения обнаружено И.М. Сеченовым в 1362 гиду. Торможение в ЦНС препятствует развитию возбуждения или ослабляет протекающее возбуждение. Примером торможения может быть прекращение рефлекторной реакции, на фоне - действия другого более сильного раздражителя. Первоначально была предложена унитарно-химическая теория торможения. Она основывалась на принципе Дейла: один нейрон - один медиатор. Согласно ей торможение обеспечивается теми же нейронами и синапсами, что и возбуждение. В последующем была доказана правильность бинарно-химической теории. В соответствии с последней, торможение обеспечивается специальными тормозными нейронами, которые являются вставочными. Это клетки Реншоу спинного мозга и нейроны Пуркинье промежуточного. Торможение в ЦНС необходимо для интеграции нейронов в единый нервный центр. В ЦНС выделяют следующие механизмы торможения:
1.| Постсинаптическое. Оно возникает в постсинаптической мембране сомы и дендритов нейронов, т.е. после передающего синапса. На этих участках образуют аксо-дендритные или аксосоматические синапсы специализированные тормозные нейроны . Эти синапсы являются глицинергическими. В результате воздействия, НЛИ на глициновые хеморецепторы постсинаптической мембраны, открываются, ее калиевые и хлорные каналы. Ионы калия и хлора входят в нейрон, развивается ТПСП. Роль ионов хлора в развитии ТПСП: небольшая. В результате возникшей гиперполяризации возбудимость нейрона падает. Проведение нервных, импульсов через него прекращается. Алкалоид стрихнин может связываться с глицериновыми рецепторами постсинаптической мембраны и выключать тормозные синапсы. Это используется для демонстрации роли торможения. После введения стрихнина у животного развиваются судороги всех мышц.
2. Пресинаптическое торможение. В этом случае тормозной нейрон образует синапс на аксоне нейрона, подходящем к передающему синапсу. Т.е. такой синапс является аксо-аксональным (рис). Медиатором этих синапсов служит ГАМК. Под действием ГАМК активируются хлорные каналы постсинаптической мембраны. Но в этом случае ионы хлора начинают выходить из аксона. Это приводит к небольшой локальной, но длительной деполяризации его мембраны. Значительная часть натриевых каналов мембраны инактивируется, что блокирует проведение нервных импульсов по аксону, а следовательно выделение нейромедиатора в передающем синапсе. Чем ближе тормозной синапс расположен к аксонному холмику, тем сильнее его тормозной эффект. Пресинаптическое торможение наиболее эффективно при обработке информации, так как проведение возбуждения блокируется не во всем нейроне, а только на его одном входе. Другие синапсы, находящиеся на нейроне продолжают функционировать.
3. Пессимальное торможение. Обнаружено Н.Е. Введенским. Возникает при очень высокой частоте нервных импульсов. Развивается стойкая длительная деполяризация всей мембраны нейрона и инактивация ее натриевых каналов. Нейрон становится невозбудимым. В нейроне одновременно могут возникать и тормозные и возбуждающие постсинаптические потенциалы. За счет этого и происходит выделение нужных сигналов.
Начнем с названия. Правильный перевод этого словосочетания может очень многое объяснить.
То есть тетаническая стимуляция — это стимуляция импульсами, подаваемыми с высокой частотой. В нашем случае тетаническая стимуляция — это высокочастотная стимуляция пресинапса, после которой в постсинаптическом нейроне возникает потенциация. Отсюда и название явления — посттетаническая потенциация.
В результате длительной активизации синапса потенциалами действия, имеющими очень высокую частоту, его пресинаптическая мембрана становится легко проницаемой для ионов кальция, которым богата окружающая синапс внеклеточная среда. Кальций начинает накапливаться в пресинапсе и, как мы помним из материалов предыдущих глав, запускает химический механизм, в результате которого медиатор начинает высвобождаться из везикул. Таким образом, явление потенциации связывают с накоплением ионов кальция в пресинапти- ческом окончании, вследствие которого каждый пресинаптический потенциал вызывает высвобождение большого числа квантов медиатора.
В случае посттетанической потенциации медиатора высвобождается так много, что одиночный нервный импульс, приходящий вслед за предшествовавшим ему ритмическим раздражением, может вызвать синаптическую реакцию нейрона, гораздо более сильную, чем та, которую он вызывал до ритмического раздражения.
В различных структурах мозга посттетаническая потенциация может длиться от нескольких минут до нескольких часов. На рисунке 26 приведен пример посттетанической потенциации моносинаптиче- ского рефлекторного разряда.
Это один из опытов Д. Ллойда, проведенных еще в 1949 году. Осциллограмма опыта очень длинная, поэтому она разбита на ряд фрагментов, обозначенных литерами от А до И. Каждый всплеск на осциллограмме — это потенциал действия нейрона, возникающий в ответ на приход импульса в пресинапс. Обратите внимание на отрезок осциллограммы под литерой А. Первые четыре потенциала действия (они отмечены цифрами 1, 2, 3,4) — это контроль. Ответ нейрона на одиночное раздражение афферентного волокна. Сразу после четвертого ответа на афферентное волокно нанесли тетаническое раздражение. Шестой всплеск на осциллограмме — это уже потенциал действия, возникший на стимуляцию афферентного волокна после его тетанизации. Он еще не очень велик, но уже отличается от контрольных значений. Эффективность синаптического действия в этом случае не так высока, вероятно, потому, что непосредственные запасы медиатора, которые клетка имела до тетанизации, в результате такого мощного воздействия несколько поиссякли. Зато уже следующие реакции на стимуляцию афферентного волокна становятся все более и более мощными.

Рис 26. Посттетаническая потенциация моносинаптического рефлекторного разряда 1
Усиление реакции является очень стойким и сохраняется несколько минут. Это хорошо видно на отрезках, обозначенных литерами Б и В. Реакции клетки, возникающие на каждое раздражение афферентного волокна, здесь в несколько раз мощнее, чем в контроле. На отрезке, обозначенном литерой Г, уже хорошо видно, что со временем реакции начинают слабеть и через несколько минут они возвращаются к контрольным значениям. Это демонстрирует отрезок осциллограммы под литерой И.
Посттетаническая потенциация возникает только в том случае, если ритмический разряд и тестирующий импульс поступают к клет Д. Ллойд, 1949.
ке по одним и тем же афферентным волокнам. Если же ритмический разряд и тестирующий стимул прошли по разным волокнам, имеющим окончания на одном и том же нейроне, поттенциация в этом нейроне не произойдет. Это свидетельствует о том, что эффект посттетаниче- ской потенциации обусловлен изменениями, произошедшими только в тех синаптических окончаниях, через которые прошли частые ритмические импульсы. В других синапсах на этой же клетке никаких изменений не происходит. То есть посттетаническая потенциация — это гомосинаптический процесс. Это очень важная характеристика, и чуть ниже мы объясним, почему.
Естественно, что такой замечательный механизм сразу же привлек к себе внимание огромного числа физиологов. В первую очередь пост- тетаническую потенциацию стали использовать для изучения механизмов замыкания временных связей, возникающих в центральной нервной системе при обучении. Однако некоторые исследователи отнеслись со здоровым скептицизмом к предположению, что посттетаническая потенциация играет значимую роль в этих процессах. И причины для появления такого скептицизма были достаточно основательными.
Согласно мнению П. Г. Костюка (1977 г.), основных причин три. Первая причина — это относительная непродолжительность посттетанической потенциации. Как мы уже отметили, она длится от нескольких минут до нескольких часов, а временные связи, возникающие при обучении, сохраняются в течение многих дней и даже месяцев. Вторая причина — это гомосинаптичность посттетанической потенциации. Для ее возникновения, как мы только что отметили, необходимо, чтобы и ритмическая стимуляция, и тестирующие стимулы проходили по одним и тем же волокнам, к одним и тем же синапсам. А для образования временной связи нужно, чтобы во времени совпали возбуждающие влияния из двух разных источников — центров безусловного и условного рефлексов. То есть влияния, приходящие по разным системам волокон. Наконец, третьей причиной является сам механизм возникновения посттетанической потенциации. В его основе лежит высокая частота импульсации, поступающей в пресинапс. Посттетаническая потенциация хорошо развивается при частоте раздражения 100—200 импульсов в секунду. В естественных условиях клетки не разряжаются с такой частотой. Ритмическая импульсация нейронов центральной нервной системы в большинстве случаев имеет более низкую частоту. Ее средние значения лежат в пределах от 5 до 40 импульсов в секунду. Поэтому в естественных условиях рефлекторной деятельности развитие посттетанической потенциации не может быть очень частым явлением. Тем не менее исследование механизмов, лежащих в основе этого процесса, продолжает оставаться одной из главных проблем нейрофизиологии, поскольку посттетаническая потенциация служит показателем пластических возможностей, как отдельных нервных клеток, так и всей нервной системы.
Еще одной формой проявления пластических свойств клеток нервной системы является пресинаптическое торможение, о котором мы упомянули в предыдущей главе. Пресинаптическое торможение по своей функциональной значимости является как бы антиподом посттетани- ческой потенциации. Если посттетаническая потенциация приводит к усилению деятельности синаптических окончаний, то пресинаптическое торможение на длительное время снижает эффективность синаптической передачи. При этом такое снижение эффективности синапсов, как и при посттетанической потенциации, зависит в первую очередь от изменений, возникающих не в пост-, а в пресинаптической мембране, и связано, как полагают исследователи, с уменьшением количества медиатора, освобождаемого каждым приходящим в пресинаптическое окончание импульсом.
Существенное различие между посттетанической потенциацией и пресинаптическим торможением состоит в том, что если первый процесс, как мы отметили, всегда гомосинаптичен, то второй может быть как гомо-, так и гетеросинаптичным. Пресинаптическое торможение может распространяться не только на те афферентные волокна, по которым прошел вызывающий его импульс, но и на соседние, по системе аксо-аксональных связей.
Нервный центр – это совокупность нейронов, обеспечивающих регуляцию какого-либо конкретного физиологического процесса или функции.
Нервный центр в узком смысле – это совокупность нейронов, без которых данная конкретная функция не может регулироваться. Например, без нейронов дыхательного центра продолговатого мозга дыхание прекращается. Нервный центр в широком смысле — это совокупность нейронов, которые участвуют в регуляции конкретной физиологической функции, но не являются строго обязательными для ее осуществления! Например, в регуляции дыхания кроме нейронов продолговатого мозга участвуют нейроны пневмотаксического центра варолиевого моста, отдельные ядра гипоталамуса, кора больших полушарий и другие образования головного мозга.
Все нейроны нервного центра разделяют на 2 неравные по количеству и качеству группы.
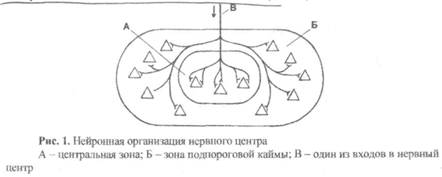
Первая группа – нейроны центральной зоны. Это наиболее возбудимые нейроны, которые возбуждаются в ответ на поступление порогового (для нервного центра) сигнала. Таких нейронов около 15-20%, и они не обязательно располагаются в середине нервного центра, как это изображено на рис.1. Особенностью их является то, что они имеют на своем теле больше синаптических терминалей от сенсорных и вставочных нейронов.
Вторая группа – нейроны подпороговой каймы. Это менее возбудимые нейроны, которые не возбуждаются в ответ на поступление пороговых им-пульсов, но при действии более сильных раздражителей они возбуждаются и включаются в работу нервного центра, обеспечивая ее усиление. Таких нейронов большинство (80-85%), и они не обязательно располагаются на периферии нервного центра, но все имеют значительно меньше синаптических терминалей от сенсорных и вставочных нейронов по сравнению с нейронами центральной зоны.
На рис. 1 нейроны центральной зоны условно поставлены в центр внутреннего круга (А), а нейроны подпороговой каймы – в пространство между внутренним и наружным кругами (Б). Таким образом, если к нервному центру по афферентному входу (В) придет пороговый импульс, то возбудятся три нейрона центральной зоны, а на десяти нейронах подпороговой каймы потенциалы действия не возникнут, но появится местная деполяризация – возбуждающий постсинаптический потенциал (ВПСП).
От структуры нервного центра зависят его свойства, а они, в свою очередь, влияют на процесс проведения возбуждения через нервный центр, на его скорость и степень выраженности. От свойств нервных центров во многом зависит процесс распространения возбуждения по ЦНС, что имеет важное значение в интегративной деятельности организма.
Свойства нервных центров обусловлены описанной выше нейронной организацией нервного центра, а также химическим способом передачи возбуждения в синапсах. При электрическом способе передачи возбуждения нервные центры не имели бы подобных свойств.
Свойства нервных центров: 1 одностороннее проведение возбуждения; 2 задержка проведения возбуждения; 3 суммация; 4 облегчение; 5 окклюзия; 6 мультипликация; 7 трансформация; 8 последействие; 9 посттетаническая потенциация; 10 утомление; 11 тонус; 12 высокая чувствительность к изменению состояния внутренней среды организма; 13 пластичность.
3) Суммация – это возникновение возбуждения в нервном центре при поступлении к нему нескольких допороговых импульсов, каждый из которых в отдельности не может возбуждения (рис. 2). Фактически этот процесс происходит на нейронах подпороговой каймы. Различают два вида суммации: пространственную и временною.
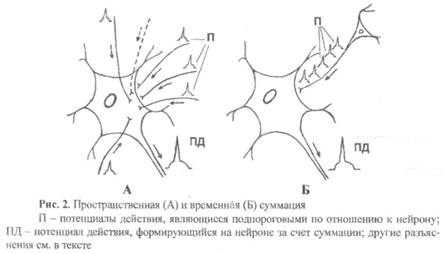
Пространственная суммация возникает в том случае, когда к нервному центру (к его нейронам) приходят одновременно, несколько допороговых импульсов. На рисунке 2А видно, что к нейрону подпороговой каймы, имеющему пороговый потенциал 30 мВ одновременно по пяти различным афферентным входам (их аксоны обозначены сплошной линией) приходят пять импульсов, каждый из которых деполяризует мембрану нейрона на 5 мВ (то есть возникают пять отдельных ВПСП). В этом случае возбуждение нейрона не наступает, так как суммарная деполяризация мембраны нейрона составляет лишь 25 мВ (суммированный ВПСП мал для достижения КУД). Но если к нейрону придет еще один подобный импульс по шестому входу (его аксон обозначен пунктирной линией), то суммированный ВПСП будет достаточен по величине и мембрана нейрона в зоне аксонного холмика деполяризуется до критического уровня, в результате чего нейрон из состояния покоя перейдет в состояние возбуждения. На постсинаптической мембране происходит суммация ВПСП в пространстве.
Временная (последовательная) суммация возникает в том случае, когда к нейронам нервного центра по одному афферентному входу приходит не один, а серия импульсов с очень небольшими по времени межимпульсными промежутками (рис. 2Б). Два механизма временной суммации:
1) интервалы между отдельными импульсами настолько малы, что за это время медиатор, выделившийся в синаптическую щель, не успевает полностью разрушиться и вернуться в пресинаптический аппарат. В этом случае возникает постепенное накопление медиатора до критического объема, необходимого для возникновения достаточного по амплитуде ВПСП, а значит, и для возникновения ПД;
2) интервалы между отдельными импульсами настолько малы, что возникший за это время на постсинаптической мембране ВПСП не успевает исчезнуть и усиливается за счет новой порции медиатора – суммируется. На постсинаптической мембране происходит суммация ВПСП во времени.
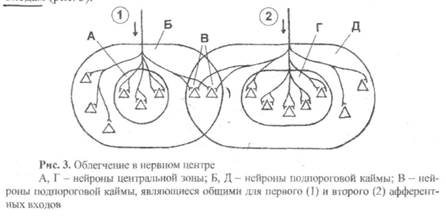
4) Облегчение – это увеличение количества возбужденных нейронов в нервном центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному, а по двум или более афферентным входам. На рис. 3 рассмотрен случай, когда при отдельном раздражении первого афферентного входа возбуждается только три нейрона центральной зоны (А), а на пяти нейронах подпороговой каймы (Б) возникают ВПСП. Если раздражать отдельно только второй афферентный вход, то возбуждены будут пять нейронов (Г), а четыре нейрона подпороговой каймы (Д) не возбудятся. Раздражая и первый, и второй афферентные входы одновременно (!), мы ожидаем вовлечения в процесс возбуждения восьми нейронов. И они, естественно, будут возбуждаться, но кроме них (сверх ожидания!) могут возбуждаться еще некоторые нейроны подпороговой каймы. Это произойдет потому, что один или несколько нейронов подпороговой каймы являются общими как для первого, так и для второго афферентных входов (в нашем случае это два нейрона, обозначенные буквой В), и при одновременном поступлении возбуждения к этим нейронам дни возбудятся за счет возникновения пространственной суммации.
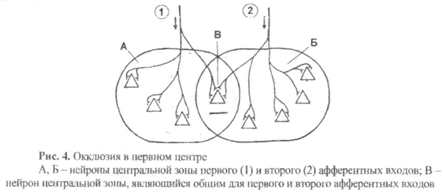
5) Окклюзия – это уменьшение количества возбужденных нейронов в нервной центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному. а по двум или более афферентным входам (рис. 4).
6) Мультипликационное возбуждение (мультипликация) заключается в том, что по разветвлениям аксона вставочного нейрона возбуждение поступает одновременно не на один, а на несколько моторных нейронов (рис. 6). В связи с этим эффект на рабочем органе усиливается в несколько раз, или в работу вовлекаются не одна, а несколько рабочих структур, Это свойство особенно ярко проявляется в ганглиях автономной (вегетативной) нервной системы.
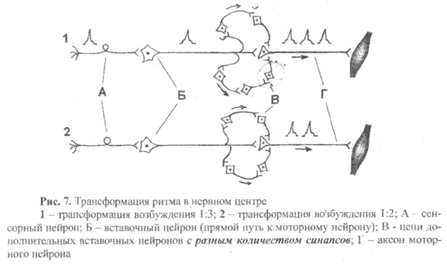
7) Трансформация ритма возбуждения – это изменение частоты импульсов на выходе из нервного центра по сравнению с частотой импульсов на входе в нервный центр.
а) наличием дублирующих цепей вставочных нейронов, связывающих сенсорные и моторные нейроны;
б) разным количеством синапсов в каждой из этих цепей.


Например, на рис.7 представлены два варианта трансформации, которые, на первый взгляд, не отличаются друг от друга, так как в том и в другом случае показаны две дополнительные цепи вставочных нейронов (кроме прямого пути), с помощью которых возбуждение может передаваться по цепи нейронов А—Б—В. Рассмотрим эти схемы.
Вариант 1. Верхняя цепь состоит из двух дополнительных вставочных нейронов, а значит, по сравнению с прямым путем передачи возбуждения с нейрона Б на нейрон В, имеет два дополнительных синапса. Поэтому возбуждение, проходя по верхней цепи, задержится на 2 мс (время синаптической задержки в одном синапсе составляет
1 мс) и придет на нейрон В после того, как пройдет возбуждение по прямому пути. В нижней цепи три дополнительных вставочных нейрона (то есть три дополнительных синапса), значит, возбуждение будет доходить до нейрона В еще дольше, чем по верхней цепи (задержка составит 3 мс). Следовательно, по нижней цепи возбуждение на нейрон В придет после того, как пройдет возбуждение по верхней цепи. В результате на один импульс, пришедший по сенсорному нейрону А, на моторном нейроне В возникнет три потенциала действия (трансформация 1:3).
Вариант 2. В этом случае и верхняя и нижняя цепи вставочных нейронов состоят из двух дополнительных нейронов. Возбуждение по обеим цепям придет к нейрону В одновременно в виде одного потенциала действие, который появится на нейроне В только после прохождения возбуждения к нему от нейрона Б по прямому пути. В этом варианте мы тоже получим трансформацию ритма, но уже в соотношении 1:2.
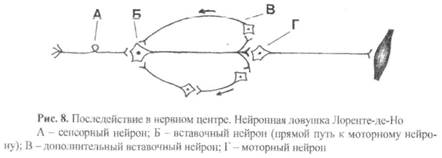
8) Последействие – это продолжение возбуждения моторного нейрона в течение некоторого времени после прекращения действия раздражителя.
Другими причинами последействия могут быть:
а) возникновение высокоамплитудного ВПСП, в результате которого возникает не один, а несколько потенциалов действия то есть ответ длится большее время;
б) длительная следовая деполяризация постсинаптической мембраны, в результате чего возникают несколько потенциалов действия, вместо одного.
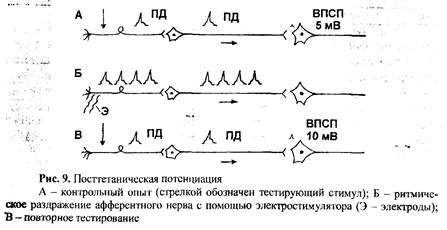
9) Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей.
Если в качестве контроля вызвать одиночное раздражение афферентного нерва тестирующим раздражителем (рис. 9А), то на моторном нейроне мы получим ВПСП вполне определенной амплитуды (в нашем случае 5 мВ). Если после этого тот же афферентный нерв раздражать некоторое время серией частых импульсов (рис. 9Б), а потом вновь подействовать тестирующим раздражителем (рис. 9В), то величина ВПСП будет больше (в нашем случае 10 мВ). Причем она будет тем больше, чем более частыми импульсами мы раздражали афферентный нерв.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения: после одиночных стимулов оно выражено слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. Объясняется он тем, что при частом раздражении афферентного волокна в его пресинаптической терминали (окончании) накапливаются ионы кальция, а значит, улучшается выделение медиатора. Кроме того, показано, что частое раздражение нерва приводит к усилению синтеза медиатора, мобилизации пузырьков медиатора, к усилению синтеза белков-рецепторов на постсинаптической мембране и увеличению их чувтствительности. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах.
10) Утомление нервного центра (посттетаническая депрессия, синаптическая депрессия) – это уменьшение или прекращение импульсной активности нервного центра в результате длительной стимуляции его афферентными импульсами (или произвольного вовлечения его в процесс возбуждения посредством импульсов, идущих из коры больших полушарий). Причинами утомления нервного центра могут быть:
• истощение запасов медиатора в афферентном или вставочном нейроне;
• снижение возбудимости постсинаптической мембраны (то есть мембраны моторного или вставочного нейрона) из-за накопления, например, продуктов метаболизма.
Утомляемость нервных центров продемонстрировал Н.Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. tibialis и п. peroneus. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. Синаптическая депрессия при длительной активации центра выражается в снижении постсинаптических потенциалов.
11) Тонус нервного центра – это длительное, умеренное возбуждение нервного центра без видимо утомления Причинами тонуса могут быть:
• потоки афферентных импульсов, постоянно поступающие с неадаптирующихся рецепторов;
• гуморальные факторы, постоянно присутствующие в плазме крови;
• спонтанная биоэлектрическая активность нейронов (автоматия);
• циркуляция (реверберация) импульсов в ЦНС.
12) Нервный центр состоит из нейронов, а они очень чувствительны к изменению состава внутренней среды организма, что и отражается на свойствах нервных центров. Наиболее важными факторами, влияющими на работу нервных центров, являются: гипоксия; недостаток питательных веществ (например, глюкозы); изменение температуры; воздействие продуктов метаболизма; воздействие различных токсических и фармакологических препаратов.
Разные нервные центры имеют неодинаковую чувствительность к воздействию названных факторов. Так, нейроны коры больших полушарий наиболее чувствительны к гипоксии, недостатку глюкозы, продуктам метаболизма; клетки гипоталамуса – к изменению температуры, содержанию глюкозы, аминокислот, жирных кислот и др.; различные участки ретикулярной формации выключаются разными фармакологическими препаратами, различные нервные центры избирательно активируются или тормозятся разными медиаторами.
13) Пластичность нервного центра означает его способность изменять при определенных обстоятельствах свои функциональные свойства. В основе этого явления лежит поливалентность нейронов нервных центров. Особенно ярко проявляется это свойство при всевозможных повреждениях ЦНС, когда организм компенсирует утраченные функции за счет сохранившихся нервных центров. Особенно хорошо свойство пластичности выражено в коре больших полушарий. Например, центральные параличи, связанные с патологией двигательных центров коры, иногда полностью компенсируются, и ранее утраченные двигательные функции восстанавливаются.
Дата добавления: 2015-01-21 ; просмотров: 21321 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Читайте также:
