
Тела вставочных нейронов вегетативной нервной системы
Вставочный нейрон, известный так же как ассоциативный или интернейрон, присутствует только в тканях ЦНС, взаимосвязан исключительно с другими нервными клетками. Эта особенность отличает его от сенсорных или моторных аналогов. Сенсорные взаимодействуют с другими системами организма, к примеру, с кожными рецепторами и органами чувств, когда преобразуют стимулы, поступающие из внешней среды в биоэлектрические сигналы. Моторные клетки иннервируют волокна мышечной ткани и обеспечивают двигательную активность человека.
Виды и характеристики нейронов
Нервные клетки, именуемые нейронами, принимают, отправляют и проводят биоэлектрические сигналы. Различают эфферентные (двигательные) нейроны – это компоненты ЦНС, которые перенаправляют сигналы исполнительным органам, к примеру, скелетным мышцам. Афферентные (чувствительные) нейроны – это такие клетки, которые воспринимают внешние и внутренние стимулы, что обеспечивает связь организма с внешней средой и реакции на изменение функциональной активности внутренних органов.
Вставочные клетки обеспечивают взаимосвязи в рамках общей нейрональной сети. Нейроны всех типов (чувствительные, эфферентные, ассоциативные) являются функциональными единицами, поддерживающими деятельность нервной системы, они находятся во всех тканях организма, где играют роль связующих звеньев между рецепторными (воспринимающими раздражающие стимулы) и эффекторными органами, которые отвечают на раздражающие стимулы.
К эффекторным органам относят мышцы и железы, к рецепторным – органы чувств. Значение проводимых сигналов существенно различается в зависимости от вида клетки и ее роли в функционировании ЦНС. К примеру, чувствительные, воспринимающие импульсы внешней среды, передают сигналы от кожных рецепторов и органов чувств в направлении головного мозга, двигательные нейроны перенаправляют команды, сформированные в мозге, вызывающие сокращение скелетных мышц и инициирующие движение.
Несмотря на разное значение биоэлектрических импульсов, их природа одинакова и заключается в изменении показателей электрического потенциала в области плазматической мембраны нервной клетки. Механизм распространения нервных импульсов основан на способности электрического возмущения, появившегося в одном месте клетки, передаваться на другие участки. При отсутствии факторов, усиливающих сигнал, импульсы затухают по мере удаления от источника возбуждения.
Сенсорный, известный так же как чувствительный – это афферентный нейрон, который проводит импульсы от дистальных участков организма к центральным отделам ЦНС. К примеру, сенсорные образуют волокна, отходящие от светочувствительных клеток органов зрения. Сигналы отходят от сетчатки глаза, направляясь по миллионам аксонов, принадлежащих структурам базальных ганглий, в направлении участка зрительной коры.
Чувствительный нейрон в совокупности с исполнительными (двигательными) нейронами образует простую рефлекторную дугу.
К примеру, коленный рефлекс – безусловная рефлекторная реакция растяжения, возникает в результате активности подобной рефлекторной дуги. Реакция в виде неконтролируемого разгибания голени происходит при механическом воздействии на сухожилие мышцы бедра, пролегающее под надколенником. Механизм реакции:
- Механическое воздействие на нервно-мышечные веретена, пролегающие в мышце-разгибателе бедра.
- Повышение интенсивности нервных сигналов в окончаниях, оплетающих нервно-мышечные веретена, вследствие их растяжения.
- Передача импульсов чувствительным нейронам, находящимся в спинальных ганглиях, посредством дендритов, отходящих от бедренного нерва.
- Передача импульсов от чувствительных клеток альфа-мотонейронам, пролегающим в передних рогах в границах спинного мозга.
- Передача сигнала от альфа-мотонейронов способным к сокращению мышечным волокнам бедренной мышцы.
В механизме коленного рефлекса принимают участие интернейроны, которые передают тормозящие импульсы мотонейронам мышц-сгибателей, и другие вставочные нейроны, к примеру, клетки Реншоу. В механизме коленного рефлекса также задействованы гамма-мотонейроны, которые регулируют интенсивность растяжения веретен.
В спинном мозге, образованном серым веществом, расположены нейроны трех типов – моторные, вставочные, вегетативные. Причем вегетативные находятся в висцеральных (относящихся к внутренним органам) ядрах. Эти клетки взаимодействуют с афферентными (восходящие проводящие пути, которые передают импульсы от периферических рецепторов в центральные зоны ЦНС) волокнами, отвечающими за общую висцеральную чувствительность.
Висцеральные афференты проводят нервные сигналы (чаще болезненные или рефлекторные ощущения) от внутренних органов, элементов кровеносной системы, желез к соответствующим зонам ЦНС. Висцеральные афференты находятся в составе вегетативного отдела нервной системы. Рефлекторные дуги в рамках вегетативного отдела ЦНС отличаются строением от дуг соматического отдела.
Чувствительный нейрон – это такой компонент нервной системы, который передает в мозг информацию о раздражителях, воздействующих на определенный участок тела. Примером раздражителей могут служить факторы: солнечный свет, механическое воздействие (удар, касание), действие химического вещества. Чувствительные нейроны расположены в ганглиях мозга – спинного и головного.
Связь, образованная с чувствительным нейроном, может провоцировать возбуждение или торможение, которое направляется по нервным волокнам к корковым отделам мозга. По мере возрастания уровня сенсорных путей, передаваемая информация перерабатывается с идентификацией важных признаков. Чувствительные относятся к псевдоуниполярным нейронам – их аксон и дендриты отходят от тела вместе, впоследствии разделяются и находятся в спинном, головном мозге (аксон) и в периферических отделах тела (дендриты).
Вставочные нейроны передают преобразованные нервные импульсы, полученные в результате обработки сенсорной информации, поступившей из разных источников, к примеру, от органов зрения и кожных рецепторов. В результате переработанная информация становится исходными данными для формирования адекватных двигательных команд.
Двигательные нервные клетки бывают двух видов – крупные и мелкие. В первом случае речь идет об α-мотонейронах, во втором – о γ-мотонейронах. Альфа-мотонейроны присутствуют в базальных ядрах латеральной (ближе к боковой плоскости) и медиальной (ближе к срединной плоскости) локализации. Это самые крупные клетки, присутствующие нервной ткани.
Их аксоны взаимодействуют с поперечнополосатыми волокнами, содержащимися в составе скелетных мышц. В результате образуются синапсы (места передачи нервных сигналов). Аксоны альфа-мотонейронов взаимосвязаны со вставочными аналогами, известными так же как клетки Реншоу, что приводит к формированию коллатеральных путей и тормозных синапсов в спинном мозге.
Гамма-мотонейроны находятся в составе нервно-мышечного веретена, которое представляет собой сложный рецептор, состоящий из нервных окончаний (афферентных, эфферентных). Главная функция нервно-мышечных веретен заключается в регуляции силы и скорости сокращения или растяжения мускулатуры скелета.
Строение и функции
Вставочная клетка состоит из тела, от которого отходят единичный аксон и дендриты. Дендриты вставочных клеток чаще короткие. Их аксоны вариативно переходят в границах спинного мозга из задних рогов в передние (замыкают дугу на уровне отрезка спинного мозга) или распространяются в область других уровней мозговых структур – спинных, головных.
Одна из функций вставочных нейронов – торможение интенсивности некоторых сигналов. К примеру, интернейроны неокортекса (новой коры, отвечающей за высшие психические функции – сенсорное восприятие, осознанное мышление, произвольную двигательную активность, речь) избирательно понижают интенсивность части сигналов, поступающих из таламуса, чтобы предотвратить необходимость отвлекаться на посторонние, малозначащие стимулы. Если импульсация, спровоцированная внешним стимулом, недостаточно сильна, она может затухнуть, не доходя до коркового слоя головного мозга.
Область влияния вставочных клеток ограничена индивидуальными особенностями строения – длина отростков-аксонов, количество коллатеральных ответвлений. Обычно вставочные оснащены аксонами с терминалями (концевой участок, представленный синаптическим окончанием – местом контакта с другими клетками), заканчивающимися в пределах одного центра, что обуславливает интеграцию в рамках группы.
Вставочные нейроны замыкают рефлекторные дуги, они воспринимают возбуждение от афферентных нервных структур, перерабатывают данные и передают их двигательным нейронам. Ассоциативные клетки играют ведущую роль в формировании нейрональных сетей, где продлевается срок хранения поступающей и переработанной информации.
Порядок взаимодействия
Рефлекторная регуляция функций организма в интерпретированной, упрощенной форме описана в учебнике биологии для 8-го класса. Вставочные, сенсорные и двигательные нейроны взаимосвязаны. Характер взаимодействия зависит от вида функций нервной системы. Примерный порядок взаимодействия в случае функций чувствительных нейронов, которые локализованы в области кожных покровов:
- Восприятие внешнего стимула нервным рецептором, расположенным в коже.
- Передача стимула сенсорными клетками к зонам головного мозга. Обычно сигнал проходит через 2 синапса (в спинном мозге и таламусе), затем попадает в сенсорную зону коры полушарий.
- Преобразование импульса в универсальную форму.
- Передача преобразованного импульса во все корковые отделы полушарий при помощи вставочных нейронов, которые находятся только в ЦНС.
Произвольные движения мышц осуществляются благодаря активности мотонейронов, находящихся в корковой двигательной зоне. Мотонейроны инициируют движение – сигнал поступает в скелетные мышцы по эфферентным волокнам. В то время как основные сигналы, отправленные мотонейронами, поступают к мышечной ткани, возбуждение распространяется на другие участки мозга, к примеру, на область оливы и мозжечка, где происходит тонкая настройка планирующегося действия.
Вставочные клетки играют роль посредников, обеспечивающих связь между эфферентными и афферентными нервными клетками.
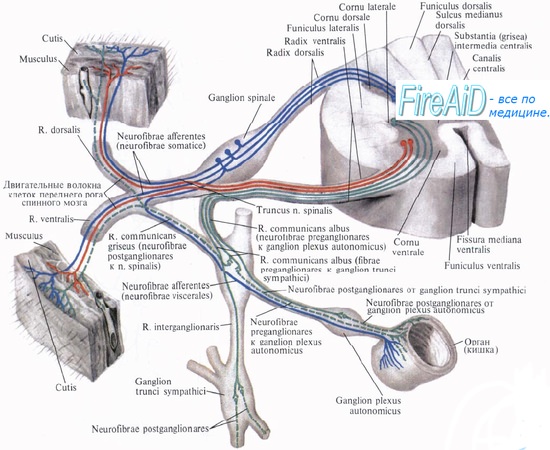
Рефлекторная дуга вегетативной нервной системы.
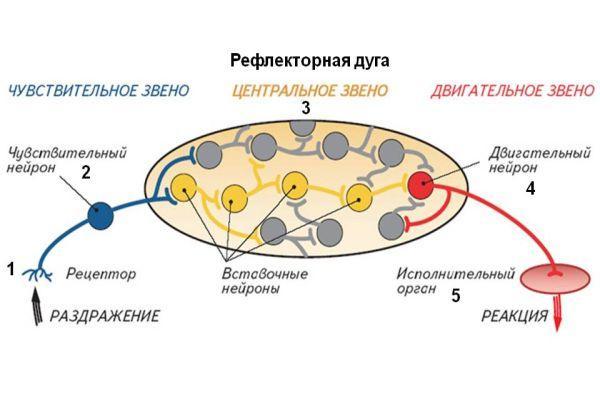
Клеточное тело воспринимающего нейрона как для анимальной, так и для вегетативной нервной системы помещается в спинномозговом узле, ganglion spinale, куда стекаются афферентные пути как от органов животной жизни, так и от органов растительной жизни и который, таким образом, является смешанным анимально-вегетативным узлом.
Клеточное тело вставочного нейрона вегетативной нервной системы в отличие от анимальной нервной системы помещается в боковых рогах спинного мозга. При этом аксон вставочного анимального нейрона, исходящий из клеток заднего рога, заканчивается в пределах спинного мозга среди клеток его передних рогов. Что же касается вставочного нейрона вегетативной нервной системы, то он в спинном мозге не заканчивается, а выходит за его пределы, к нервным узлам, расположенным на периферии.
Выйдя из спинного мозга, аксон вставочного нейрона подходит или к узлам симпатического ствола, ganglia trunci sympathici, относящимся к симпатическому отделу вегетативной нервной системы (они образуют симпатический ствол), или волокна не заканчиваются в этих узлах, а направляются к предпозвоночным узлам, расположенным более периферично, между симпатическим стволом и органом (ganglia coeliaca, ganglia mesenterica).
Кроме макроскопически видимых обособленных узлов, по ходу вегетативных нервов встречаются мигрировавшие сюда в ходе эмбрионального развития небольшие группы эффекторных нейронов — микроганглии. Все волокна, идущие до узлов первого, второго или третьего порядка и являющиеся аксонами промежуточного нейрона, называются предузловыми волокнами, rami preganglionares. Они покрыты миелином.

Третий, эффекторный, нейрон анимальной рефлекторной дуги помещается в передних рогах спинного мозга, а эффекторный нейрон вегетативной рефлекторной дуги вынесен в процессе развития из центральной нервной системы в периферическую, ближе к рабочему органу, и располагается в вегетативных нервных узлах. Из такого расположения эффекторных нейронов на периферии вытекает главный признак вегетативной нервной системы — двухнейронность эфферентного периферического пути: первый нейрон — вставочный; тело его лежит в вегетативных ядрах черепных нервов или боковых рогах спинного мозга, а нейрит идет к узлу; второй — эфферентный, тело которого лежит в узле, а нейрит достигает рабочего органа.
Эффекторные нейроны симпатических нервов начинаются в ganglia trunci sympathici (узлы первого порядка) или ganglia intermedia (узлы второго порядка), а для парасимпатических нервов — в около- или внутриорганных узлах, ganglia terminalia (третьего порядка); так как в названных узлах осуществляется связь вставочных и эфферентных нейронов, то отмеченная разница между симпатическим и парасимпатическим отделами вегетативной нервной системы связана именно с этими нейронами.
Аксоны эфферентных вегетативных нейронов почти лишены миелина — безмиелиновые (серые). Они составляют послеузловые волокна, rami postganglionics. Послеузловые волокна симпатической нервной системы, отходящие от узлов симпатического ствола, расходятся в двух направлениях. Одни волокна идут к внутренностям и составляют висцеральную часть симпатической системы. Другие волокна образуют rami communicantes grisei, соединяющие симпатический ствол с анимальными нервами.
В составе последних волокна достигают соматических органов (аппарата движения и кожи), в которых иннервируют непроизвольную мускулатуру сосудов и кожи, а также железы.
Совокупность описанных эфферентных вегетативных волокон, идущих от узлов симпатического ствола до органов сомы, составляет соматическую часть симпатического отдела. Такая структура обеспечивает функцию вегетативной нервной системы, которая регулирует обмен веществ всех частей организма применительно к непрерывно изменяющимся условиям среды и условиям функционирования (работы) тех или иных органов и тканей.
Соответственно этой наиболее универсальной своей функции, связанной не с какими-либо отдельными органами и системами, а со всеми частями, со всеми органами и тканями организма, вегетативная нервная система и морфологически характеризуется универсальным, повсеместным распространением в организме.
Следовательно, симпатический отдел иннервирует не только внутренности, но и сому, обеспечивая в ней обменные и трофические процессы.
В результате каждый орган, по И. П. Павлову, находится под тройным нервным контролем, в связи с чем он различает три вида нервов:
1) функциональные, осуществляющие функцию данного органа;
2) сосудодвигательные, обеспечивающие доставку крови к органу, и
3) трофические, регулирующие усвоение из доставленной крови питательных веществ.
Висцеральная часть симпатического отдела содержит все эти три вида нервов для внутренностей, а соматическая часть — только сосудодвигательные и трофические. Что же касается функциональных нервов для органов сомы (скелетная мускулатура и др.), то они идут в составе соматической, ани-мальной, нервной системы.
Таким образом, основное отличие эфферентной части вегетативной нервной системы от эфферентной части анимальной заключается в том, что анимальные, соматические, нервные волокна, выйдя из центральной нервной системы, идут до рабочего органа, нигде не прерываясь, тогда как вегетативные волокна на своем пути от мозга до рабочего органа прерываются в одном из узлов первого, второго или третьего порядка. Вследствие этого эфферентный путь вегетативной нервной системы разбивается на две части, из которых он и состоит: предузловые миелиновые волокна, rami preganglionares, и послеузловые, лишенные миелина (безмиелиновые) волокна, rami postganglionares.
Наличие узлов в эфферентной части рефлекторной дуги составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной.
14.1. Компоненты нервной системы
14.1.1. Подразделение нервной системы
на центральную и периферическую
2. Последняя включает:
1. а) Нервные узлы (или ганглии) - это скопления нервных клеток (точнее, их тел) вне центральной нервной системы.
б) Скопления же нейронов в головном или спинном мозгу называются ядрами .
2. а) А нервные стволы - это совокупность идущих параллельно нервных волокон.
б) Среди последних в нерве могут одновременно присутствовать
а) два типа образований периферической нервной системы -
б) а также один из двух компонентов центральной нервной системы -
б) Они различаются в строении и локализации нервных ядер, узлов и стволов.
14.1.2. Соматическая нервная система
и её рефлекторная дуга
14.1.2.1. Введение
2. Её рефлекторная дуга обычно включает три части :

14.1.2.2. Чувствительные нейроны
б) Узлы же располагаются по ходу
в) А. В первом случае узел называется спинномозговым (1) .
а) В одних случаях они (как показано на схеме)
б) В других случаях аксоны чувствительных нейронов
14.1.2.3. Ассоциативные нейроны
б) Тела последующих ассоциативных нейронов (если они вовлечены в дугу) могут находиться также в других отделах ЦНС: в т.ч. в
б) Аксоны же могут
14.1.2.4. Двигательные нейроны
Б. Импульсы от ассоциативных нейронов обычно поступают по многочисленным аксо соматическим синапсам (п. 13.3.4).
б) Тела (8.А) двигательных нейронов находятся
б) А б лагодаря восходящим и нисходящим связям спинного мозга с корой больших полушарий (создающим более сложные дуги), человек может
14.1.3. Вегетативная нервная система
14.1.3.1. Функция
I. Общие сведения
а) В свою очередь, вегетативная нервная система подразделяется на два отдела, или две системы:
II. Действия симпатической и парасимпатической систем
процессы восстановления:
спазма сосудов и
усиления сердцебиения,
14.1.3.2. Общие особенности вегетативной нервной системы
Вегетативная нервная система имеет ряд особенностей.-
б) По отношению к этим ганглиям, различают 2 вида вегетативных нервных волокон:
б) Медиаторы же в окончаниях пост ганглионарных волокон различны:
14.1.3.3. Рефлекторные дуги
симпатической нервной системы
В случае симпатической нервной системы вышеперечисленные особенности проявляются следующим образом.
а) Локализация остальных частей данных нейронов обычна:

б) Постганглионарные волокна (10) , отходящие от экстрамуральных узлов, во-первых,
в) Кроме того, от узлов симпатического ствола часть постганглионарных волокон (в составе т.н. серых соединительных ветвей)
14.1.3.4. Рефлекторные дуги
парасимпатической нервной системы
б) Аксоны этих нейронов образуют преганглионарные парасимпатические волокна и идут в составе
б) Поэтому постганглионарные волокна парасимпатической системы (образованные аксонами эффекторных нейронов) -
14.1.3.5. Периферические вегетативные рефлекторные дуги
но также других нейронов:
б) Тогда последовательность прохождения сигнала и ответа такова.-
Теперь рассмотрим подробней морфологию упоминавшихся выше компонентов нервной системы -
14.2. Нервные стволы и нервные узлы
14.2.1. Нервные стволы
14.2.1.1. Соединительнотканные элементы
а) Малое увеличение

прослойки соединительной ткани между пучками называются периневрием (5 ).
б) Большое увеличение

14.2.1.2. Нервные волокна
б ) В данном препарате нерв состоит, главным образом, из миелиновых нервных волокон (п. 12.4.3.1) :
б) Большое увеличение
14.2.2. Нервные узлы
Здесь будет рассмотрено строение трёх типов узлов -
14.2.2.1. Чувствительные узлы
а) В качестве пример а обратимся к спинномозговы м узл ам .
б) Строение чувствительных узлов, лежащих по ходу некоторых черепномозговых нервов, аналогично.
I. Корешки спинного мозга и местоположение спинномозговых узлов

II. Строение узла
б ) А. В центре узла ( между группами нейронов ) проходят нервные волокна ( 5 ).
б) В соответствии с п. 12.2.2.2 , эти нейроны имеют

14.2.2.2. Симпатические узлы
Согласно п. 14.1.3.4, такие узлы находятся
I. Общий вид

II. Эффекторные нейроны

б) Видимо, с одним нейроном контактируют сразу несколько преганглионарных волокон (чем и объясняется наличие нескольких дендритов).
2. Аксоны эффекторных нейронов , покидая узел и подходя к органу, образуют , как мы уже говорили (п. 14.1.3. 2) ,
III. МИФ-клетки
2. Их на данном препарате различить не удаётся.
3. а) Считают, что, возбуждаясь преганглионарными волокнами, МИФ-клетки частично
14.2.2.3. Интрамуральные ганглии
I. Общий вид
1. а) Интрамуральный ганглий (1) выявляется как скопление нервных клеток в толще органа.
б) В данном случае видна окружающая мышечная ткань (2) стенки мочевого пузыря.
а) 
б) Вокруг них, как обычно, находятся глиальные клетки-сателлиты (4) и соединительнотканные элементы.
б) (Другое поле зрения) .jpg)
II. Типы нейронов
14.3. Спинной мозг
2. Как видно, схема устанавливает связь между
14.3.2. План строения
14.3.2.1. Оболочки мозга
2. Первые две образованы рыхлой волокнистой соединительной тканью,
4. а) Паутинная оболочка не заходит в углубления мозга.
б) Поэтому между ней и мягкой оболочкой образуется
В самом спинном мозгу можно различить
14.3.2.2. Серое вещество: общие сведения

14.3.2.3. Белое вещество: общие сведения
б) Между передними канатиками - глубокая срединная вырезка (1 0 ), просвет которой на препарате не всегда виден.
Теперь дадим более детальную характеристику серого и белого вещества.
14.3.3. Серое вещество спинного мозга
14.3.3.1. Функциональные структуры серого вещества
А. ЗАДНИЕ РОГА
Как мы знаем, в задних рогах содержатся вставочные (ассоциативные) нейроны, которые получают сигналы от чувствительных нейронов спинномозговых узлов.
Нейроны задних рогов образуют следующие структуры.
Аксоны этих нейронов идут к мотонейронам передних рогов того же сегмента спинного мозга -
Б. ПРОМЕЖУТОЧНАЯ ЗОНА И БОКОВЫЕ РОГА
Здесь нейроны сгруппированы в два или одно ядро (в зависимости от уровня спинного мозга).
В. ПЕРЕДНИЕ РОГА
Г. НЕЙРОНЫ, СОДЕРЖАЩИЕСЯ ВО ВСЕХ РОГАХ
14.3.3.2. Классификация нейронов спинного мозга
I. Классификация всех нейронов по местонахождению их аксонов
II. Классификация клеток передних рогов
2. Поэтому данные клетки, иннервируя в скелетных мышцах экстрафузальные мышечные волокна, участвуют
2. Таким образом, они стимулируют цепочку событий. -
14.3.3.3. Просмотр препарата
Рассмотрим отдельные фрагменты предыдущего препарата.
Мы видим глиальную строму, в которой располагаются нейроны.

а) Нейроны (1), лежащие ближе к центральному каналу,

а) Видны крупные мотонейроны (1) с телами неправильной формы и отходящими от них отростками.

14.3.4. Белое вещество спинного мозга
1. а) Микроскопическое строение белого вещества спинного мозга однообразно:
на поперечном срезе это сечения миелиновых (как правило) нервных волокон.
б) В центре каждого волокна - осевой цилиндр, который окружён толстой миелиновой оболочкой.
2. Но в функциональном отношении проводящие пути белого вещества различны. –
1. Восходящие пучки
(Голля и Бурдаха) -
2. Восходящие пучки:
3. Нисходящие пучки:
а) от коры больших полушарий
(боковой пирамидный тракт );
а) Содержат аксоны нейронов головного мозга.
4. Нисходящие пучки:
а) от коры больших полушарий
( передний пирамидный тракт) ,
б) от среднего мозга
(от подкорковых центров зрения и слуха),
Медицинский эксперт статьи


Вегетативная (автономная) нервная система (systema nervosum autonomicum) представляет собой часть нервной системы, которая контролирует функции внутренних органов, желез, сосудов, осуществляет адаптационно-трофическое влияние на все органы человека. Вегетативная нервная система поддерживает постоянство внутренней среды организма (гомеостаз). Функция вегетативной нервной системы неподконтрольна человеческому сознанию, однако она находится в подчинении спинного мозга, мозжечка, гипоталамуса, базальных ядер конечного мозга, лимбической системы, ретикулярной формации и коры полушарий большого мозга.
Выделение вегетативной (автономной) нервной системы обусловлено некоторыми особенностями ее строения. К этим особенностям относятся следующие:
- очаговость расположения вегетативных ядер в ЦНС;
- скопление тел эффекторных нейронов в виде узлов (ганглиев) в составе периферических вегетативных сплетений;
- двухнейронность нервного пути от ядер в ЦНС к иннервируемому органу;
- сохранение признаков, отражающих более медленную эволюцию вегетативной нервной системы (в сравнении с анимальной): меньший калибр нервных волокон, меньшая скорость проведения возбуждения, отсутствие у многих нервных проводников миелиновой оболочки.
Вегетативная (автономная) нервная система подразделяется на центральный и периферический отделы.
К центральному отделу относятся:
- парасимпатические ядра III, VII, IX и X пар черепных нервов, лежащие в мозговом стволе (средний мозг, мост, продолговатый мозг);
- парасимпатические крестцовые ядра, залегающие в сером веществе трех крестцовых сегментов спинного мозга (SII-SIV);
- вегетативное (симпатическое) ядро, расположенное в боковом промежуточном столбе [латеральное промежуточное (серое) вещество] VIII шейного, всех грудных и двух верхних поясничных сегментов спинного мозга (СVIII-ТhI-LII).
К периферическому отделу вегетативной (автономной) нервной системы относятся:
- вегетативные (автономные) нервы, ветви и нервные волокна, выходящие из головного и спинного мозга;
- вегетативные (автономные) висцеральные сплетения;
- узлы вегетативных (автономных, висцеральных) сплетений;
- симпатический ствол (правый и левый) с его узлами, межузловыми и соединительными ветвями и симпатическими нервами;
- узлы парасимпатической части вегетативной нервной системы;
- вегетативные волокна (парасимпатические и симпатические), идущие на периферию (к органам, тканям) от вегетативных узлов, входящих в состав сплетений и расположенных в толще внутренних органов;
- нервные окончания, участвующие в вегетативных реакциях.
Нейроны ядер центрального отдела вегетативной нервной системы являются первыми эфферентными нейронами на путях от ЦНС (спинного и головного мозга) к иннервируемому органу. Волокна, образованные отростками этих нейронов, носят название предузловых (преганглионарных) нервных волокон, так как они идут до узлов периферической части вегетативной нервной системы и заканчиваются синапсами на клетках этих узлов.
Вегетативные узлы входят в состав симпатических стволов, крупных вегетативных сплетений брюшной полости и таза, а также располагаются в толще или возле органов пищеварительной, дыхательной систем и мочеполового аппарата, которые иннервируются вегетативной нервной системой.
Размеры вегетативных узлов обусловлены количеством расположенных в них клеток, которое колеблется от 3000-5000 до многих тысяч. Каждый узел заключен в соединительнотканную капсулу, волокна которой, проникая в глубь узла, разделяют его на дольки (секторы). Между капсулой и телом нейрона расположены клетки-сателлиты - разновидность глиальных клеток.
К глиальным клеткам (шванновские клетки) относят нейролеммоциты, образующие оболочки периферических нервов. Нейроны вегетативных ганглиев подразделяются на два основных типа: клетки Догеля I типа и II типа. Клетки Догеля I типа эфферентные, на них заканчиваются преганглионарные отростки. Для этих клеток типичны длинный тонкий неветвящийся аксон и множество (от 5 до нескольких десятков) дендритов, ветвящихся возле тела этого нейрона. Эти клетки имеют несколько маловетвящихся отростков, среди которых имеется аксон. Они крупнее нейронов Догеля I типа. Их аксоны вступают в синаптическую связь с эфферентными нейронами Догеля I типа.
Преганглионарные волокна имеют миелиновую оболочку, благодаря чему они отличаются беловатым цветом. Они выходят из мозга в составе корешков соответствующих черепных и спинномозговых нервов. Узлы периферической части вегетативной нервной системы содержат тела вторых эфферентных (эффекторных) нейронов, лежащих на путях к иннервируемым органам. Отростки этих вторых нейронов, несущих нервный импульс из вегетативных узлов к рабочим органам (гладкой мускулатуре, железам, сосудам, тканям), являются послеузловыми (постганглионарными) нервными волокнами. У них нет миелиновой оболочки, и поэтому они имеют серый цвет.
Скорость проведения импульсов по симпатическим преганглионарным волокнам составляет 1,5-4 м/с, а парасимпатическими - 10-20 м/с. Скорость проведения импульса по постганглионарным (безмиелиновым) волокнам не превышает 1 м/с.
Тела афферентных нервных волокон вегетативной нервной системы располагаются в спинномозговых (межпозвоночных) узлах, а также в чувствительных узлах черепных нервов; в собственных чувствительных узлах вегетативной нервной системы (клетки Догеля II типа).
Строение рефлекторной вегетативной дуги отличается от строения рефлекторной дуги соматической части нервной системы. У рефлекторной дуги вегетативной нервной системы эфферентное звено состоит не из одного нейрона, а из двух. В целом простая вегетативная рефлекторная дуга представлена тремя нейронами. Первое звено рефлекторной дуги - это чувствительный нейрон, тело которого располагается в спинномозговых узлах или узлах черепных нервов. Периферический отросток такого нейрона, имеющий чувствительное окончание - рецептор, берет начало в органах и тканях. Центральный отросток в составе задних корешков спинномозговых нервов или чувствительных корешков черепных нервов направляется к соответствующим вегетативным ядрам спинного или головного мозга. Эфферентный (выносящий) путь вегетативной рефлекторной дуги представлен двумя нейронами. Тело первого из этих нейронов, второго по счету в простой вегетативной рефлекторной дуге, располагается в вегетативных ядрах центральной нервной системы. Этот нейрон можно называть вставочным, так как он находится между чувствительным (афферентным, приносящим) звеном рефлекторной дуги и третьим (эфферентным, выносящим) нейроном эфферентного пути. Эффекторный нейрон представляет собой третий нейрон вегетативной рефлекторной дуги. Тела эффекторных нейронов лежат в периферических узлах вегетативной нервной системы (симпатический ствол, вегетативные узлы черепных нервов, узлы вне- и внутриорганных вегетативных сплетений). Отростки этих нейронов направляются к органам и тканям в составе органных вегетативных или смешанных нервов. Заканчиваются постганглионарные нервные волокна в гладких мышцах, железах, в стенках сосудов и в других тканях соответствующими концевыми нервными аппаратами.
На основании топографии вегетативных ядер и узлов, различий в длине первого и второго нейронов эфферентного пути, а также особенностей функций вегетативная нервная система подразделяется на две части: симпатическую и парасимпатическую.
Физиология вегетативной нервной системы
Вегетативная нервная система контролирует артериальное давление (АД), частоту сердечных сокращений (ЧСС), температуру и массу тела, пищеварение, метаболизм, водно-электролитный баланс, потоотделение, мочеиспускание, дефекацию, половые реакции и другие процессы. Многие органы управляются в основном либо симпатической, либо парасимпатической системой, хотя они могут получать входящие импульсы из обоих отделов вегетативной нервной системы. Чаще действие симпатической и парасимпатической систем на один и тот же орган прямо противоположное, например симпатическая стимуляция повышает частоту сердечных сокращений, а парасимпатическая - снижает.
Парасимпатическая нервная система способствует восстановлению затраченных организмом ресурсов, т.е. обеспечивает процессы анаболизма. Парасимпатическая вегетативная нервная система стимулирует секрецию пищеварительных желез и моторику желудочно-кишечного тракта (включая эвакуацию), снижает частоту сердечных сокращений и артериальное давление, а также обеспечивает эрекцию.
Функции вегетативной нервной системы обеспечивают два основных нейромедиатора - ацетилхолин и норадреналин. В зависимости от химической природы медиатора нервные волокна, секретирующие ацетилхолин, называют холинергическими; это все преганглионарные и все постганглионарные парасимпатические волокна. Волокна, секретирующие норадреналин, называют адренергическими; ими являются большинство постганглионарных симпатических волокон, за исключением иннервирующих кровеносные сосуды, потовые железы и мышцы arectores pilorum, которые являются холинергическими. Ладонные и подошвенные потовые железы частично отвечают и на адренергическую стимуляцию. Подтипы адренергических и холинергических рецепторов различают в зависимости от их локализации.
Оценка вегетативной нервной системы
Заподозрить вегетативную дисфункцию можно при наличии таких симптомов, как ортостатическая гипотензия, отсутствие толерантности к высокой температуре и потеря контроля над функцией кишечника и мочевого пузыря. Эректильная дисфункция - один из ранних симптомов дисфункции вегетативной нервной системы. Ксерофтальмия и ксеростомия не являются специфическими симптомами дисфункции вегетативной нервной системы.

[1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12]
Устойчивое снижение систолического артериального давления более чем на 20 мм рт. ст. или диастолического более чем на 10 мм рт. ст. после принятия вертикального положения (при отсутствии обезвоживания организма) предполагает наличие вегетативной дисфункции. Следует обращать внимание на изменение частоты сердечных сокращений (ЧСС) во время дыхания и при перемене положения тела. Отсутствие дыхательной аритмии и недостаточный прирост ЧСС после принятия вертикального положения указывают на вегетативную дисфункцию.
Миоз и умеренный птоз (синдром Горнера) свидетельствуют о поражении симпатического отдела вегетативной нервной системы, расширенный и не реагирующий на свет зрачок (зрачок Эйди) - о поражении парасимпатической вегетативной нервной системы.
Патологические мочеполовые и ректальные рефлексы могут также быть симптомами недостаточности вегетативной нервной системы. Исследование включает оценку кремастерного рефлекса (в норме штриховое раздражение кожи бедра приводит к поднятию яичек), анального рефлекса (в норме штриховое раздражение перианальной кожи приводит к сокращению анального сфинктера) и бульбо0кавернозного рефлекса (в норме сдавление головки полового члена или клитора приводит к сокращению анального сфинктера).
При наличии симптомов вегетативной дисфункции с целью определения степени выраженности патологического процесса и объективной количественной оценки вегетативной регуляции сердечно0сосудистой системы проводятся кардиовагальная проба, пробы на чувствительность периферических а-дренорецепторов, а также количественная оценка потоотделения.
Количественным судомоторным аксонрефлекстестом проверяется функция постганглионарных нейронов. Локальное потоотделение стимулируется ионофорезом ацетилхолина, электроды устанавливают на голени и запястье, выраженность потоотделения регистрируется специальным судометром, передающим в аналоговой форме информацию на компьютер. Результатом теста может быть снижение потоотделения, либо его отсутствие, либо продолжение потоотделения после прекращения стимуляции. С помощью терморегуляторной пробы оценивают состояние преганглионарных и постганглионарных проводящих путей. Значительно реже для оценки функции потоотделения используют красящие пробы. После нанесения на кожу краски пациента помещают в закрытое помещение, которое нагревается до достижения максимального потоотделения; потоотделение приводит к изменению цвета краски, что выявляет области ангидроза и гипогидроза и позволяет провести их количественный анализ. Отсутствие потоотделения свидетельствует о поражении эфферентной части рефлекторной дуги.
Кардиовагальные пробы оценивают реакцию ЧСС (регистрация и анализ ЭКГ) на глубокое дыхание и пробу Вальсальвы. Если вегетативная нервная система интактна, то максимальное увеличение ЧСС отмечается после 15-го сердечного удара и снижение - после 30-го. Отношение между интервалами RR на 15-30-м ударах (т.е. самого длинного интервала к самому короткому) - отношение 30:15 - в норме составляет 1,4 (отношение Вальсальвы).
Пробы на чувствительность периферических адренорецепторов включают изучение сердечного ритма и артериального давления в тилт-тесте (пассивной ортопробе) и пробе Вальсальвы. При проведении пассивной ортопробы происходит перераспределение объема крови в нижележащие части тела, что вызывает рефлекторные гемодинамические реакции. В пробе Вальсальвы проводят оценку изменений АД и ЧСС в результате повышения внутри грудного давления (и снижения венозного притока), что вызывает характерные изменения артериального давления и рефлекторную вазоконстрикцию. В норме изменения гемодинамических показателей происходят на протяжении 1,5-2 мин и имеют 4 фазы, в период которых АД повышается (1-я и 4-я фазы) или снижается после быстрого восстановления (2-я и 3-я фазы). ЧСС увеличивается в первые 10 с. При поражении симпатического отдела возникает блокада ответа во 2-й фазе.
Читайте также:
