
Топография симпатической нервной системы
Каждый из двух симпатических стволов подразделяют на четыре отдела: шейный, грудной, поясничный (или брюшной) и крестцовый (или тазовый).
Шейный отдел простирается от основания черепа до шейки I ребра; располагается позади сонных артерий на глубоких мышцах шеи. В его состав входят три шейных симпатических узла: верхний, средний и нижний.
Ganglion cervicale superius является самым крупным узлом симпатического ствола, имея длину около 20 мм и ширину 4-6 мм. Лежит он на уровне 11 и части Ill шейных позвонков позади внутренней сонной артерии и медиально от n.vagus.
Ganglion cervicale medium небольшой величины, располагается обыкновенно в месте перекреста a.thyroidea inferior с сонной артерией, нередко отсутствует или может распадаться на два узелка.
Ganglion cervicale inferius довольно значительной величины, расположен позади начальной части позвоночной артерии; нередко сливается с I, а иногда и 11 грудным узлом, образуя общий шейно-грудной, или звездчатый, узел, ganglion cervicothoracicum s.ganglion stellatum.
От шейных узлов отходят нервы для головы, шеи и груди. Их можно разделить на восходящую группу, направляющуюся к голове, на нисходящую - опускающуюся к сердцу, и группу для органов шеи.
Нервы для головы отходят от верхнего и нижнего шейных узлов и делятся на группу, проникающую в полость черепа, и группу, подходящую к голове снаружи.
Первая группа представлена n.caroticus internus, отходящим от верхнего шейного узла, и n.vertebralis, отходящим от нижнего шейного узла. Оба нерва, сопровождая одноименные артерии, образуют вокруг них сплетения: plexus caroticus internus и plexus vertebralis; вместе с артериями они проникают в полость черепа, где анастомозируют между собой и дают ветви к сосудам мозга, оболочкам, гипофизу, стволам III, IV, V, VI пар черепных нервов и барабанному нерву.
Plexus caroticus internus продолжается в plexus cavernosus, которое окружает a.carotis interna на участке прохождения ее через sinus cavernosus.
Ветви сплетений распространяются, кроме самой внутренней сонной артерии, также по ее разветвлениям. Из ветвей plexus caroticus internus следует отметить n.petrosus profundus, который присоединяется к n.petrosus major и вместе с ним образует n.canalis pterygoidei, подходящий через одноименный канал к ganglion pterygopalatinum.
Вторая группа симпатических нервов головы, наружная, составляется двумя ветвями верхнего шейного узла, nn.carotici externi, которые, образовав сплетение вокруг наружной сонной артерии, сопровождают ее разветвления на голове. От этого сплетения отходит стволик к ушному узлу, gangl. oticum; от сплетения, сопровождающего лицевую артерию, отходит ветвь к поднижнечелюстному узлу, gangl. submandibulare.
Через посредство ветвей, входящих в сплетения вокруг сонной артерии и ее ветвей, верхний шейный узел дает волокна к сосудам (вазоконстрикторы) и железам головы: потовым, слезной, слизистым и слюнным, а также к мышцам волос кожи и к мышце, расширяющей зрачок, m.dilatator pupillae. Центр расширения зрачка, centrum ciliospinale, находится в спинном мозге на уровне от VIII шейного до II грудного сегмента.
Органы шеи получают нервы от всех трех шейных узлов; кроме того, часть нервов отходит от межузловых участков шейного отдела симпатического ствола, а часть - от сплетений сонных артерий.
Веточки от сплетений следуют по ходу ветвей наружной сонной артерии, носят одноименные названия и вместе с ними подходят к органам, в силу чего число отдельных симпатических сплетений равно числу артериальных ветвей. Из нервов, отходящих от шейной части симпатического ствола, отмечают гортанно-глоточные ветви от верхнего шейного узла - rami laryngopharyngei, которые частые идут с n.laryngeus superior (ветвь n.vagi) к гортани, частые спускаются к боковой стенке глотки; здесь они вместе с ветвями языкоглоточного, блуждающего и верхнего гортанного нервов образуют глоточное сплетение, plexus pharyngeus.
Нисходящая группа ветвей шейной части симпатического ствола представлена nn.cardiaci cervicales superior, medius et inferior, отходящими от соответствующих шейных узлов. Шейные сердечные нервы спускаются в грудную полость, где вместе с симпатическими грудными сердечными нервами и ветвями блуждающего нерва участвуют в образовании сердечных сплетений.
Грудной отдел симпатического ствола располагается впереди шеек ребер прикрыт спереди плеврой. В его состав входят 10-12 узлов более или менее треугольной формы. Грудной отдел характеризуется присутствием белых соединительных ветвей, rami communicantes albi, соединяющих передние корешки спинномозговых нервов с узлами симпатического ствола. Ветви грудного отдела:
1) nn.cardiaci thoracici отходят от верхних грудных узлов и участвуют в образовании plexus cardiacus (подробное описание сердечных сплетений cм. при описании сердца);
2) rami communicantes grisei, безмиелиновые - к межреберным нервам (соматическая часть симпатического отдела);
3) rami pulmonales - к легким, образуют plexus pulmonalis;
4) rami aortici образуют сплетение на грудной аорте, plexus aorticus thoracicus, и частью на пищеводе, plexus esophageus, а также на грудном протоке (во всех указанных сплетениях принимает участие и n.vagus);
5) nn.splanchnici major et minor, большой и малый внутренностные нервы; n.splanchnicus major начинается несколькими корешками, отходящими от V-IX грудных узлов; корешки n.splanchnicus major идут в медиальном направлении и сливаются на уровне IX грудного позвонка в один общий ствол, проникающий через промежуток между мышечными пучками ножек диафрагмы в брюшную полость, где он входит в состав plexus coeliacus; n.splanchnicus minor начинается от X - XI грудных узлов и также входит в plexus coeliacus, проникая через диафрагму с большим внутренностным нервом. В этих нервах проходят сосудосуживающие волокна, как это видно из того обстоятельства, что при перерезке этих нервов сосуды кишечника сильно переполняются кровью; в nn.splanchnici содержатся волокна, тормозящие движение желудка и кишок, а также волокна, служащие проводниками ощущений от внутренностей (афферентные волокна симпатической части).
Поясничный, или брюшной, отдел симпатического ствола состоит из четырех, иногда из трех узлов . Симпатические стволы в поясничном отделе расположены на более близком расстоянии один от другого, чем в грудней полости, так что узлы лежат на переднебоковой поверхности поясничных позвонков вдоль медиального края m. psoas major. Rami communicantes albi имеются только с двумя или тремя верхними поясничными нервами.
От брюшного отдела симпатического ствола на всем протяжении отходит большое количество ветвей, которые вместе с nn.splanchnici major et minor и брюшными отделами блуждающих нервов образуют самое большое непарное чревное сплетение, plexus coeliacus. В формировании чревного сплетения участвуют также многочисленные спинномозговые узлы (C5 - L3), аксоны их нейроцитов. Оно лежит на передней полуокружности брюшной аорты, позади поджелудочной железы, и окружает начальные части чревного ствола (truncus coeliacus) и верхней брыжеечной артерии. Сплетение занимает участок между почечными артериями, надпочечниками и аортальным отверстием диафрагмы и включает парный чревный узел, ganglion coeliacum, и иногда непарный верхний брыжеечный узел, ganglion mesentericum superius.
От чревного сплетения отходит ряд меньших парных сплетений к диафрагме, надпочечникам, почкам, а также plexus testicularis (ovaricus), следующих по ходу одноименных артерий. Имеется также ряд непарных сплетений к отдельным органам по стенкам артерий, название которых они носят.
Из последних верхнее брыжеечное сплетение, plexus mesentericus superior, иннервирует поджелудочную железу, тонкую и толстую кишку до половины протяжения поперечной ободочной.
Вторым глазным источником иннервации органов полости живота является сплетение на аорте, plexus aorticus abdominalis, составленное из двух стволов, отходящих от чревного сплетения, и веточек от поясничных узлов симпатического ствола. От аортального сплетения отходит нижнее брыжеечное сплетение, plexus mesentericus inferior, для поперечной и нисходящей части ободочной кишки, сигмовидной и верхних отделов rectum (
plexus rectalis superior). У места отхождения plexus mesentericus inferior располагается одноименный узел, gangl. mesentericum inferius. Его постганглионарные волокна идут в тазе в составе nn.hypogastrici.
Аортальное сплетение продолжается вначале в непарное верхнее подчревное сплетение, plexus hypogastricus superior, которое у мыса раздваивается и переходит в сплетение таза, или нижнее подчревное сплетение (plexus hypogastricus inferior s.plexus pelvinus).
Волокна, происходящие из верхних поясничных сегментов, по своей функции являются сосудодвигательными (вазоконстрикторами) для полового члена, двигательными для матки и сфинктера мочевого пузыря.
Крестцовый, или тазовый, отдел имеет обычно четыре узла; располагаясь на передней поверхности крестца вдоль медиального края передних крестцовых отверстий, оба ствола книзу постепенно сближаются друг с другом, а затем оканчиваются в одном общем непарном узле - ganglion impar, находящемся на передней поверхности копчика. Узлы тазового отдела, как и поясничного, связаны между собой не только продольными, но и поперечными стволиками.
От узлов крестцового отдела симпатического ствола отходит ряд ветвей, которые соединяются с ветвями, отделяющимися от нижнего брыжеечного сплетения, и образуют пластинку, протягивающуюся от крестца к мочевому пузырю; это так называемое нижнее подчревное, или тазовое, сплетение, plexus hypogastricus inferior s.plexus pelvinus. Сплетение имеет свои узелки - ganglia pelvina. В сплетении различают несколько отделов:
1) передненижний отдел, в котором выделяют верхнюю часть, иннервирующую мочевой пузырь - plexus vesicalis, и нижнюю, снабжающую у мужчин предстательную железу (plexus prostaticus), семенные пузырьки и семявыносящий проток (plexus deferentialis) и пещеристые тела (nn.cavernosi penis);
2) задний отдел сплетения снабжает прямую кишку (plexus rectales medii et inferiores).
У женщин выделяют еще средний отдел, нижняя часть которого дает ветви к матке и влагалищу (plexus uterovaginalis), пещеристым телам клитора (
nn.cavernosi clitoridis), а верхняя - к матке и яичникам.
От узлов крестцового отдела симпатического ствола отходят соединительные ветви, rami communicantes, присоединяющиеся к спинномозговым нервам, иннервирующим нижнюю конечность. Эти соединительные ветви составляют соматическую часть симпатического отдела вегетативной нервной системы, иннервирующую нижнюю конечность. В составе rami communicantes и спинномозговых нервов нижней конечности находятся постганглионарные волокна, которые распространяются в сосудах, железах и мышцах волос кожи, а также в скелетной мускулатуре, обеспечивая ее трофику и тонус.
4.49 Морфо-функциональные понятия об анализаторах: определение, составные части, пример.
Органами чувств, или анализаторами, называются приборы, посредством которых нервная система получает раздражения от внешней среды, а также от органов самого тела и воспринимает эти раздражения в виде ощущений.
Показания органов чувств являются источниками представлений об окружающем нас мире.
Процесс чувственного познания совершается у человека по шести каналам: осязание, слух, зрение, вкус, обоняние, земное тяготение. Шесть органов чувств дают человеку многообразную информацию об окружающем объективном мире, которая отражается в сознании в виде субъективных образов - ощущений восприятий к представлений памяти.
Живая протоплазма обладает раздражимостью и способностью отвечать на раздражение. В процессе филогенеза эта способность особенно развивается у специализированных клеток покровного эпителия под влиянием внешних раздражений и клеток кишечного эпителия под влиянием раздражения пищей. Специализированные клетки эпителия уже у кишечнополостных оказываются связанными с нервной системой. В некоторых участках тела, например на щупальцах, в области рта, специализированные клетки, обладающие повышенной возбудимостью, образуют скопления, из которых возникают простейшие органы чувств. В дальнейшем в зависимости от положения этих клеток происходит их специализация по отношению к раздражителям. Так, клетки ротовой области специализируются к восприятию химических раздражений (обоняние, вкус), клетки на выступающих частях тела - к восприятию механических раздражений (осязание) и т.д.
Развитие органов чувств обусловлено значением их для приспособления к условиям существования. Например, собака тонко воспринимает запах ничтожных концентраций органических кислот, выделяемых телом животных (запах следов), и плохо разбирается в запахе растений, которые не имеют для нее биологического значения.
Возрастание тонкости анализа внешнего мира обусловлено не только усложнением строения и функции органов чувств, но прежде всего усложнением нервной системы.Возникающие в силу тех или иных раздражений нервные возбуждения воспринимаются нами в форме различных ощущений.Так, например, световая энергия, действуя на сетчатку глаза, вызывает нервные импульсы, которые, передаваясь по нервной системе, вызывают в нашем сознании зрительные ощущения.
Для возникновения ощущений необходимы: приборы, воспринимающие раздражение, нервы, по которым передается это раздражение, и мозг, где оно превращается в факт сознания. Весь этот аппарат, необходимый для возникновения ощущения, И.П.Павлов назвал анализатором. "Анализатор - это такой прибор, который имеет своей задачей разлагать сложность внешнего мира на отдельные элементы" (Павлов И.П. Лекции по физиологии).
Каждый анализатор состоит из трех частей:
1) рецептор - трансформатор энергии раздражения в нервный процесс;
2) кондуктор-проводник нервного возбуждения;
3) корковый конец анализатора, где возбуждение воспринимается как ощущение.
Различают две группы ощущений:
1. Ощущения, отражающие свойства предметов и явлений окружающего материального мира: осязание, т.е. ощущение прикосновения и давления, температурное чувство (тепла, холода) и боль; затем ощущения слуховые, зрительные, вкусовые, обонятельные и земного притяжения.
2. Ощущения, отражающие движения отдельных частей тела и состояние внутренних органов (двигательные ощущения, ощущение равновесия тела, ощущения органов). Соответственно этому все органы чувств делят на две группы:
1. Органы внешних чувств, получающие нервные импульсы из экстероцептивного поля, - экстеpоцептоpы. Их шесть: органы кожного чувства, чувства земного тяготения (гравитации), слуха, зрения, вкуса и обоняния.
2. Органы внутренних ощущений:
а) получающие импульсы из проприоцептивного поля (мышечно-суставное чувство, тесно связанное с чувством земного притяжения)-проприоцепторы;
б) органы, воспринимающие нервные импульсы из интероцептивного поля (внутренностей и сосудов), - интеpоцeптоpы.
Ощущения, идущие из внутренних органов, обычно неопределенны и при нормальном состоянии этих органов не достигают сознания, сказываясь только "общим самочувствием". Вообще все внутренние процессы, регулируемые вегетативной нервной системой, протекают без нашего ведома и только при болезненных расстройствах дают о себе знать обычно более или менее сильной болью.
Подробно об интероцептивном анализаторе было изложено в специальной главе. Из возбуждений, идущих от проприоцептивного поля, надо упомянуть только мышечно-суставное чувство, благодаря которому воспринимается ощущение положения частей тела и происходит координация движений. С одной стороны, это чувство комбинируется с кожной чувствительностью (чувство стереогноза), в с другой, стоит в связи с органом гравитации, дающим ориентацию по отношению к гравитационному полю, который может быть рассмотрен также как статокинетический аппарат, обеспечивающий равновесие тела. Нервные окончания (в мышцах, костях, сухожилиях и суставах) и проводники мышечно-суставного чувства были описаны при изложении двигательного анализатора. В данном разделе будут рассмотрены только органы, воспринимающие ощущения, получаемые из внешнего мира, - экстероцепторы.
Общий план воспринимающих приборов у всех классов животных более или менее одинаков, несмотря на последующие значительные усложнения в деталях. Основным элементом, за исключением органов кожного чувства, у наземных животных являются особые чувствительные клетки, которые в процессе развития всегда происходят из эпителия наружного листка (эктодермы), который уже по своему положению находится в соприкосновении с окружающим миром. Каждая такая клетка на одном конце, обращенном к наружной поверхности, несет штифтик или воспринимающие волоски, а с другой стороны отдает (в органе обоняния и зрения) отросток, идут на соединение с отростками нервных клеток проводящих нейронов.
В других органах (вкуса и слуха) чувствительная клетка, не давая центрального отростка, оплетается концевыми разветвлениями подходящего к ней афферентного нерва. Первый тип чувствительных клеток сравнительно со вторым видом нужно считать первичным. У водных животных такая форма воспринимающих элементов встречается и в кожных покровах, где эти элементы подвергаются увлажнению окружающей жидкостью. В коже наземная животных чувствительных клеток не бывает, и рецепторные нервные волокна оканчиваются или свободно между клетками эпителиального покрова, или же имеют на своих концах особого рода концевые тельца. В образовании органов чувств принимает также участие мезодерма, но только вторичным порядком, образуя для них защитные, поддерживающие и вспомогательные приспособления. Эти приспособления, обрастающие и дополняющие чувствительные клетки, т.е. рецепторы, образуют вместе с ними периферические отделы органов чувств: кожа, ухо, глаз, язык, нос. Например, зрительным рецептором являются чувствительные клетки сетчатки (палочки и колбочки), а периферическим отделом - весь глаз.
Кроме деления органов чувств на 2 группы, все анализаторы можно классифицировать с точки зрения учения И.П.Павлова о двух сигнальных системах следующим образом:
I. Анализаторы первой сигнальной системы (конкретно - наглядное мышление):
А. Анализаторы внешнего мира - экстероцепторы (органы кожного чувства, слуха, зрения, вкуса, обоняния и гравитации).
Б. Анализаторы внутреннего мира организма:
1. Проприоцепторы, несущие раздражение от органов животной жизни (мышечно-суставное чувство).
2. Интероцепторы, несущие раздражение от органов растительной жизни (внутренности, сосуды).
II. Анализаторы второй сигнальной системы (абстрактно-логическое мышление):
1. Анализаторы устной речи.
2. Анализаторы письменной речи.
Анализаторы первой и второй сигнальных систем имеют резкие анатомические отличия. Анализаторы первой сигнальной системы обладают каждый всеми тремя компонентами (рецептор, кондуктор и корковый конец). Анализаторы второй сигнальной системы лишены своих рецепторов и кондукторов, а имеют только корковые концы (корковые концы речевых анализаторов); они воспринимают свои сигналы (вторые сигналы) на базе первых сигналов, составляющих первую сигнальную систему, без которой они не функционируют. Этим подчеркивается и разделение, и объединение корковых концов всех анализаторов, составляющих единую кору большого мозга.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Симпатическая нервная система влияет на гладкие мышцы кровеносных сосудов, внутренних органов брюшной полости, мочевого пузыря, прямой кишки, волосяных фолликулов и зрачков, а также на сердечную мышцу, потовые железы, слезные, слюнные и пищеварительные железы. Симпатическая система ингибирует функцию гладких мышц внутренних органов брюшной полости, мочевого пузыря, прямой кишки и пищеварительных желез, а другие органы-мишени, напротив, — стимулирует.
Диаметр артерий регулируется преимущественно симпатической нервной системой. Повышение симпатической активности приводит к вазоконстрикции, а снижение — к вазодилатации.
Анатомия симпатической нервной системы. Преганглионарные симпатические волокна начинаются в грудных (Th1—Th12) и двух первых поясничных сегментах. Некоторые преганглионарные волокна оканчиваются на вторых нейронах в узлах правого и левого симпатических стволов (на рисунке показан только левый симпатический ствол). Остальные волокна пронизывают симпатический ствол, не прерываясь в нем, и завершаются на телах вторых нейронов, размещающихся в превертебральных узлах. В каждом случае постганглионарные волокна вторых нейронов несут к органам-мишеням импульсы симпатической иннервации.
Узлы симпатического ствола. Преганглионарные волокна, начинающиеся от нейронов боковых рогов спинного мозга (в боковом промежуточном столбе), впоследствии примыкают к аксонам соматических мотонейронов и вместе с ними покидают спинной мозг в составе переднего корешка. На уровне спинномозгового узла вегетативное волокно вновь ответвляется от соматического двигательного волокна и, образуя белую соединительную ветвь, входит в симпатический ствол. Белый цвет этих волокон обусловлен высокой степенью миелинизации.
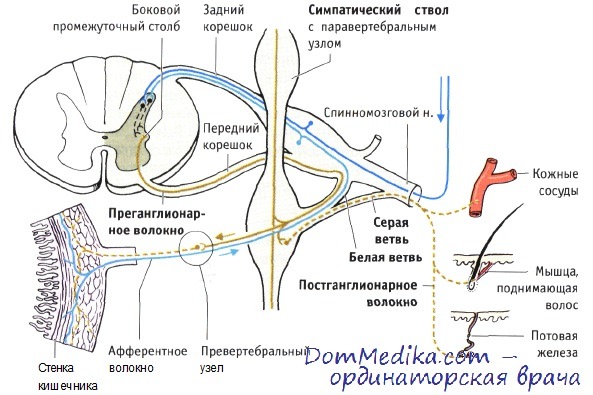
В симпатическом стволе преганглионарные волокна оканчиваются на вторых нейронах на уровне того же сегмента, который они покинули, или на несколько сегментов выше или ниже. Другие преганглионарные волокна проходят через симпатический ствол и, не прерываясь в синапсах, оканчиваются на вторых нейронах в превертебральных узлах. Безмиелиновые постганглионарные волокна выходят из симпатического ствола в виде серых соединительных ветвей и на уровне того же сегмента воссоединяются со спинномозговым нервом, волокна которого подходят к соответствующему кожному дерматому и иннервируют сосуды, мышцы, поднимающие волосы и потовые железы.
Симпатическая иннервация головы и шеи. Как уже говорилось выше, одни постганглионарные волокна достигают своих зон периферической иннервации в составе сегментарных спинномозговых нервов, а другие следуют строго вдоль кровеносных сосудов и их ветвей, что, в частности, свойственно области головы и шеи. В шейном отделе спинного мозга симпатические нейроны отсутствуют, поэтому симпатическая иннервация головы и шеи осуществляется клетками боковых рогов (боковой промежуточный столб) первых (верхние) четырех — пяти грудных сегментов.
Постганглионарные волокна, исходящие из этих сегментов, поднимаются по симпатическому стволу и завершаются в трех верхних узлах симпатического ствола: верхнем шейном узле, среднем шейном узле и шейно-грудном (звездчатом) узле. В этих узлах симпатические волокна переключаются на вторые нейроны, которые дают начало постганглионарным волокнам. Некоторые волокна в составе спинномозговых нервов подходят к шейным дерматомам. Другие, безмиелиновые волокна, начинающиеся от клеток верхнего шейного узла, образуют наружное сонное сплетение, которое распластано по одноименной артерии и ее ветвям в области головы и лица, иннервируя потовые железы, гладкие мышцы волосяных фолликулов и кровеносные сосуды.
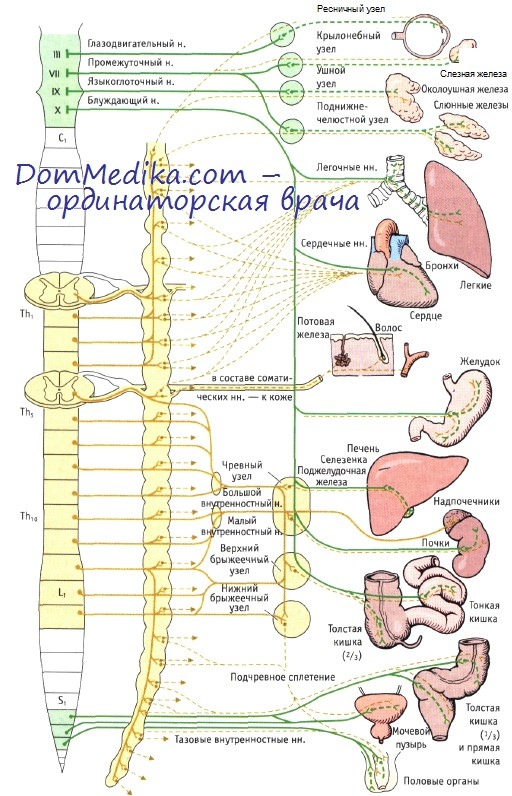
Некоторые волокна сопровождают внутреннюю сонную артерию, образуя внутреннее сонное сплетение, иннервирующее глаза (дилататор зрачка, глазничную мышцу и мышцу хряща века), а также слезные и слюнные железы. Симпатическая иннервация сердца и легких. Постганглионарные волокна шейных и верхних четырех или пяти грудных узлов проходят в составе сердечных нервов к сердечному сплетению, иннервирующему сердце. Легочные нервы иннервируют бронхи и легкие.
Симпатическая иннервация органов брюшной полости и тазовых органов. Преганглионарные волокна начинаются в грудных сегментах (Th5—Th|2) и в составе большого и малого внутренностных нервов следуют к непарным превертебральным узлам (чревный, верхний и нижний брыжеечные узлы), которые размещаются вдоль аорты рядом с местами отхождения соответствующих по названию ее ветвей. В них волокна внутренностных нервов образуют синапсы на вторых симпатических нейронах, которые дают начало постганглионарным волокнам, иннервирующим органы брюшной полости и таза. Симпатические постганглионарные волокна, в отличие от парасимпатических, обладают исключительной длиной и прежде, чем попасть в органы-мишени, образуют различные сплетения.
Мозговое вещество надпочечников. В структуре симпатической нервной системы надпочечники занимают особое место. Сходство их мозгового вещества с симпатическими узлами заключается в том, что оно напрямую иннервируется преганглионарными волокнами. В мозговом веществе надпочечников эти волокна образуют синапсы с видоизмененными вторыми нейронами, которые не рассылают нервные импульсы по аксонам, а секретируют адреналин и норадреналин непосредственно в кровоток. Усилившаяся активность симпатической нервной системы побуждает мозговое вещество надпочечников выделять адреналин и норадреналин, переносящие симпатическое влияние на периферию. В условиях стресса это имеет первостепенное значение.
По топографическому принципу вегетативную нервную систему, как и анимальную, делят на центральную и периферическую части (отделы).
Центральный отдел вегетативной нервной системы
Центры ВНС расположены в спинном и головном мозге. Их следует представлять как согласованно действующие ансамбли нейронов, ответственных за выполнение той или иной функции. Вегетативные центры разделяют на высшие (надсегментарные) и низшие (сегментарные). Координирующее влияние сегментарных центров распространяется на отдельные функции и осуществляется через определенные нервы. Надсегментарные центры контролируют деятельность сегментарных вегетативных центров, осуществляют их интеграцию с центрами соматической нервной системы и другими регулирующими системами – эндокриннной, кровеносной и т. д.
Сегментарные вегетативные нервные центры образованы телами нейронов, которые по своему положению в рефлекторной дуге являются вставочными.
По функции выделяют симпатические и парасимпатические вегетативные центры.
По топографии различают центры головного мозга (краниальные) и спинного мога (спинальные).
В отличие от строго сегментарного расположения соматических (анимальных) центров, для вегетативных нервных центров характерна очаговость. Имеются четыре таких очага:
1. мезенцефалический (парасимпатический) – добавочное ядро III пары черепных нервов, nucl. accessorius.
2. Понто-бульбарный (парасимпатический) – верхнее и нижнее слюноотделительные ядра VII и IX пары – nucl. salivatorius superior, nucl. salivatorius inferior, и вегетативное ядро X пары – nucl. dorsalis n. vagi.
Оба эти очага являются краниальными.
3. Тораколюмбальный (симпатический) – в боковых рогах спинного мозга (nucl. intermediolaterales) на протяжении сегментов С8, Th1-L2.
4. Сакральный (парасимпатический) – nucl. parasympathici sacrales, в сером веществе сегментов S2-S4.
Перечисленные очаги, или сегментарные вегетативные центры, находятся под контролирующим и коррегирующим воздействием надсегментарных (высших) центров, которые расположены в стволе мозга, мозжечке, подкорковых структурах и в коре полушарий головного мозга. Эти центры не являются специализированными (симпатическими или парасимпатическими), а объединяют в себе регуляцию обоих отделов вегетативной нервной системы. Так, в стволе головного мозга существенную роль в регуляции вегетативных функций играет ретикулярная формация, formatio reticularis, (около 100 ядер), ядра которой формируют дыхательный, сосудодвигательный, пищеварительный центры. В мозжечке – центры, регулирующие трофику кожи, сосудодвигательные рефлексы, сокращение мышц, поднимающих волосы, mm. arrectores pili. Важную роль в обеспечении вегетативных функций отводят гипоталамической области. Здесь сосредоточены центры, ответственные за поддержание постоянства внутренней среды организма (гомеостаз). Благодаря наличию обширных нервных и сосудистых связей между гипоталамусом и гипофизом, обе эти структуры объединяют в единую гипоталамо-гипофизарную систему, осуществляющую нейро-гуморальную регуляцию деятельности всех органов растительной жизни, желез внутренней секреции. В подкорковых базальных ядрах (corpus striatum et corpus amygdaloideum) содержатся центры терморегуляции, слюно- и слезоотделения.
Особое место среди высших вегетативных центров занимает лимбическая система. Это – структуры среднего, промежуточного и конечного мозга (сводчатая извилина, миндалевидное тело, мозговая полоска таламуса, гипоталамус, гиппокамп, свод, прозрачная перегородка и др.). все эти структуры объединяют в общее понятие – висцеральный мозг, в который поступает весь поток сенсорной информации и на базе ее первичного синтеза формируются определенные биологические потребности – мотивации, обеспечивается эмоциональная окраска как вегетативных так и соматических реакций организма.
И, наконец, корковые вегетативные центры, которые сосредоточены преимущественно в лобных и теменных долях и осуществляют объединение (интеграцию) вегетативных и анимальных функций всего организма.
Как отмечалось выше, в основе взаимоотношения вегетативных центров лежит принцип иерархии – объем регулирующего влияния тем больше, чем выше положение центра, причем действие высших центров реализуется не только через низшие, но и через другие регулирующие системы – эндокринную, кровеносную (например, гипоталамо-гипофизарная система).
Периферический отдел вегетативной нервной системы
К периферическому отделу ВНС относятся:
1. Вегетативные рецепторы, receptorus autonomici.
2. Вегетативные нервы, ветви и нервные волокна, выходящие из головного и спинного мозга, nn., rr. et neurofibrae autonomici.
3. Вегетативные узлы, ganglia autonomici.
4. Вегетативные (висцеральные) спетения, plexus autonomici.
Рассмотрим последовательно каждый из перечисленных компонентов периферического отдела ВНС
Вегетативные рецепторы
Рецепторы, расположенные во внутренних органах и стенках сосудов и воспринимающие изменения внутренней среды организма, называются висцерорецепторами. С их помощью осуществляется рефлекторная регуляция деятельности органов. Эти рецепторы реагируют на изменение давления в просвете сосудов (барорецепторы), степень растяжения стенки органа (механорецепторы), на электролитный состав жидкостей организма (осмо- и хеморецепторы) и др. Сигналы от этих рецепторов направляются по висцеральным афферентам или в спинной мозг вместе с соматическими чувствительными волокнами (в этом случае тела их клеток лежат в спинальных ганглиях), либо идут в составе блуждающего, лицевого, языкоглоточного нервов, – и тогда их тела располагаются в чувствительных узлах соответствующих черепных нервов.
Поступающая в ЦНС информация о состоянии внутренних органов необходима для возникновения различных мотиваций (жажды, голода и др.). следствием их является формирование сложных реакций организма.
Вегетативные нервы
Большая часть вегетативных волокон (афферентных и эфферентных) входит в состав всех спинномозговых и III, VII, IX, X пар черепных нервов, образуя их вегетативную порцию. Однако существуют и строго специализированные вегетативные нервы. К ним можно отнести белые соединительные ветви, rr. communicantes albi, серые соединительные ветви, rr. communicantes grisei, межузловые ветви, rr. interganglionares, а также внутренностные (органные) нервы, nn. splanchnici, - симпатические и парасимпатические.
Вегетативные волокна
Вегетативные волокна подразделяются, как и анимальные, на афферентные и эфферентные.
Источниками афферентных вегетативных волокон являются:
· нейроны спинномозговых узлов;
· нейроны чувствительных узлов черепных нервов;
· собственные чувствительные нейроны вегетативной нервной системы (клетки Догеля II типа).
Первые два вида волокон имеют миелиновую оболочку, их диаметр составляет 3-22 мкм, скорость проведения нервного импульса – 12-120 м/с. Отростки клеток Догеля II типа (находятся в вегетативных ганглиях) не имеют миелиновой оболочки, их диаметр не более 2 мкм, скорость проведения сигнала – 1-2 м/с.
Эфферентные нервные волокна подразделяются на преганглионарные (предузловые), постганглионарные (послеузловые) и межузловые.
Преганглионарные волокна, rr. preganglionares, представлены двумя группами волокон.
Первая группа – это аксоны нейронов вегетативных симпатических ядер спинного мозга (nucll. intermediolaterales cornulates C8, Th1-L2), которые выходят из спинного мозга в составе переднего корешка, radix anterior, затем входят в состав ствола спинномозгового нерва, truncus n. spinalis, и по выходе из позвоночного канала отделяются, образуя белые соединительные ветви, rr. communicantes albi. Эти волокна покрыты миелиновой оболочкой, благодаря чему имеют беловатый цвет. Эти ветви подходят ко всем грудным (включая шейно-грудной) и двум верхним поясничным узлам симпатического ствола, образуя синапсы на телах третьих нейронов.
Вторая группа преганглионарных волокон формируется из отростков нейронов вегетативных ядер черепных нервов (III, VII, IX, X пар) и входит в состав соответствующих нервов. Эти преганглионарные волокна подходят к конечным (парасимпатическим) ганглиям III порядка, образуя синапсы на телах клеток этих ганглиев.
Можно выделить еще и третью группу преганглионарных волокон. Это аксоны клеток парасимпатических ядер крестцового отдела, nucl. parasympathici sacrales, которые входят в состав крестцовых нервов, а затем, отделяясь от них, подходят к терминальным ганглиям в составе внутренностных крестцовых нервов, n. splanchnici sacrales s. pelvini.
Диаметр преганглионарных волокон колеблется от 1,5 до 5 мкм и более, скорость проведения возбуждения – 1,5-4 м/с в симпатических и 10-20 м/с в парасимпатических образованиях. Длина преганглионарных волокон неодинакова и зависит от удаленности вегетативных узлов от ЦНС. Поэтому наибольшую длину имеют парасимпатические преганглионарные волокна. Все преганглионарные волокна в своих концевых отделах многократно ветвятся (явление мультипликации) и образуют синапсы на нейронах вегетативных узлов.
Последнее изменение этой страницы: 2016-07-16; Нарушение авторского права страницы
Читайте также:
