
Торможение в нервной системе характеризуется тем что этот процесс











- Физиология
![]()
- История физиологии
![]()
- Методы физиологии



Торможение ЦНС
Торможение существует наряду с возбуждением и представляет собой одну из форм деятельности нейрона.
Начало изучения торможения в центральной неравной системе связывают с выходом в свет работы И.М. Сеченова "Рефлексы головного мозга", в которой он показал возможность торможения двигательных рефлексов лягушки при химическом раздражении зрительных бугров головного мозга.
Торможение в центральной нервной системе - активный нервный процесс, проявляющийся в подавлении или ослаблении процесса возбуждения.
Центральное торможение (опыт И.М. Сеченова) - процесс, характеризующийся увеличением времени рефлекса или его полным отсутствием, возникающий при раздражении кристалликом поваренной соли поперечного разреза ствола мозга в области зрительных чертогов.
Классический опыт Сеченова заключается в следующем: у лягушки с перерезанным головным мозгом на уровне зрительных бугров определяли время сгибательного рефлекса при раздражении лапки серной кислотой. После этого на зрительные бугры накладывали кристаллик поваренной соли и снова определяли время рефлекса. Оно постепенно увеличивалось, вплоть до полного исчезновения реакции. После снятия кристаллика соли и промывания мозга физиологическим раствором время рефлекса постепенно восстанавливалось. Это позволило говорить о том, что торможение — активный процесс, возникающий при раздражении определенных отделов ЦНС.
Позже И.М. Сеченовым и его учениками было показано, что торможение в ЦНС может возникнуть при нанесении сильного раздражения на любые афферентные пути.
Периферическое торможение открыто братьями Вебер в 1845 г. Они установили, что раздражение блуждающего нерва тормозит работу сердца до полной его остановки.
Благодаря микроэлектродной технике исследования стало возможным изучение процесса торможения на клеточном уровне.
Различают два вида торможения в зависимости от механизмов его возникновения: деполяризационное и гиперполяризационное. Деполяризационное торможение возникает вследствие длительной деполяризации мембраны, а гиперполяризационное - вследствие гиперполяризации мембраны.
Наступлению деполяризационного торможения предшествует состояние возбуждения. Вследствие длительного раздражения это возбуждение переходит в торможение. В основе возникновения деполяризационного торможения лежит инактивация мембраны для натрия, вследствие чего уменьшаются потенциал действия и его раздражающее влияние на соседние участки, в итоге прекращается проведение возбуждения.
Один из видов этого торможения — пессимальное, описанное Н.Е. Введенским (1886), который показал, что возбуждение может сменяться торможением в любом участке, обладающем низкой лабильностью.
Гиперполяризациоиное торможение осуществляется с участием особых тормозных структур и связано с изменением проницаемости мембраны по отношению к калию и хлору, что вызывает увеличение мембранного и порогового потенциалов, в результате чего становится невозможной ответная реакция.

Центральное торможение (опыт И.М. Сеченова): а — двигательный рефлекс на болевой раздражитель; 6 — раснространснне нервных импульсов от тормозных нейронов ствола мозга к спинному мозгу при наложении кристалла NaCI на область зрительных чертогов и отсутствие двигательного рефлекса на болевой раздражитель

Первичное торможение — процесс активации тормозных нейронов, образующих синаптические связи с клеткой, на которую направлено торможение, при этом данный процесс для клетки является первичным, не связанным с ее предварительным возбуждением.
Вторичное торможение — процесс, который развивается в клетке без участия специфических тормозных структур и является следствием ее собственного возбуждения.
Запредельное торможение - истощение нервных клеток при действии раздражителей высокой интенсивности.
Пессималыюе торможение — блокирование высокочастотных импульсов в немиелинизированных нервных терминалях вследствие их более низкой лабильности.
Пресинаптическое торможение - процесс, реализующийся при активации аксо-аксонального тормозного синапса и блокирующий возбуждающие импульсы, направленные на данную клетку.
Постсинантическое торможение - процесс, развивающийся при активации аксо-соматических и аксо-дендритических тормозных синапсов и локализующийся на собственной мембране клетки, на которую направлено торможение.
Рецинрокное торможение — взаимное подавление активности антагонистических нервных структур.
Афферентное коллатеральное торможение - частный случай реципрокного торможения, локализуемый в афферентной части рефлекторной дуги.
Эфферентное коллатеральное (возвратное) торможение — процесс, при котором тормозные вставочные нейроны действуют на те же нервные клетки, которые их активировали, при этом торможение тем сильнее, чем интенсивнее предшествующее возбуждение.
Латеральное торможение — процесс, при котором вставочные тормозные нейроны подавляют активность не только клетки, которая их инициировала, но и других, рядом расположенных.

Латеральное торможение (Т — тормозной нейрон)

Возвратное торможение (Т-тормозной вставочный нейрон (клетка Реншоу); М — мотонейрон)

Рецинрокное торможение (Т — тормозной вставочный нейрон (клетка Реншоу); М — мотонейрон)

Поступательное торможение (Т — тормозной нейрон)
Процессы торможения в центральной нервной системе
Процессы возбуждения и торможения в нервной системе тесно взаимосвязаны.
Торможение в ЦНС способствует определенной координации выполняемой функции. При этом блокируется деятельность нейронов и центров, которые в данный момент не требуются для выполнения приспособительной реакции. Кроме того, торможение выполняет и защитную функцию, предохраняя нервные клетки от перевозбуждения и истощения при действии сильных раздражителей.
Различают несколько видов торможения в нервной системе.
Постсипаптическое торможение развивается в случаях, когда тормозной медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом, что нервная клетка не может генерировать потенциал действия. Постсипаптическое торможение может быть обусловлено длительной деполяризацией или гиперполяризацией, возникающей в постсинаптической мембране вследствие взаимодействия медиатора с рецепторами, открывающими калиевые и хлорные каналы. Наиболее распространенными тормозными медиаторами являются гамма-аминомасляная кислота и глицин. Глицин выделяется особыми тормозными клетками (клетки Реншоу) в синапсах, образуемых этими клетками на мембране другого нейрона. Действуя на рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для ионов СI-, при этом ионы хлора поступают в клетку согласно концентрационному градиенту, в результате чего развивается гиперполяризация. При действии гамма-аминомасляной кислоты на постсинаптическую мембрану постсинаптическое торможение развивается в результате входа ионов хлора в клетку или выхода ионов калия из клетки. Концентрационные градиенты ионов К + в процессе развития торможения нейронов поддерживается Na + /К + -насосом, а ионов СI - — СI - -насосом.
Возвратное постсинаптическое торможение - это такое торможение, при котором тормозные вставочные нейроны (клетки Реншоу) действуют на те же нервные клетки, которые их иннервируют. Примером возвратного постсинаптического торможения может служить торможение в мотонейронах спинного мозга. Этот вид торможения обеспечивает, например, поочередное сокращение и расслабление скелетных мышц — сгибателей и разгибателей, что необходимо для координации движений конечностей при ходьбе.
Латеральное постсинаптическое торможение обусловлено тем, что тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями. В результате в этих соседних клетках развивается очень глубокое торможение, называемое латеральным, так как образующаяся зона торможения находится сбоку по отношению к возбужденному нейрону и инициируется им.
Реципрокное торможение, примером которого является торможение нервных центров мышц-антагонистов, заключается в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны. Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей. Если бы возбуждались одновременно центры мышц-сгибателей и мышц- разгибателей, сгибание конечности в суставе было бы невозможно.
Пресинаптическое торможение связано с тем, что в пресинаптическом окончании может развиваться продолжительная деполяризация мембраны, которая приводит к развитию торможения. В очаге деполяризации нарушается процесс распространения возбуждения и импульсы не могут пройти через зону деполяризации. Следовательно, не происходит выделения медиатора в синаптическую щель в достаточном количестве и не возбуждается постсинаптический нейрон. В ЦНС имеется огромное число тормозных нейронов, в частности клетки Реншоу. Эти тормозные нейроны синтезируют специфические тормозные медиаторы и осуществляют реакцию торможения. Активация тормозного нейрона вызывает деполяризацию мембраны терминалей в афферентных нейронах, что затрудняет процесс проведения потенциала действия. Медиатором втакихаксо аксональных синапсах служит гамма-аминомасляная кислота или другой тормозной медиатор. Деполяризация является следствием повышения проницаемости мембраны для ионов хлора, в результате эти ионы выходят из клетки.
Торможение — это нервный процесс, при котором возбуждение одних нейронов подавляет или предупреждает возбуждение других нейронов. Торможение никогда не распространяется по нервным волокнам.
Это местный процесс. Для торможения в ЦНС имеются специальные тормозные нейроны и тормозные синапсы. Некоторые медиаторы в ЦНС оказывают только тормозное действие, например, глицин, ГАМК, но есть медиаторы, которые оказывают как тормозное, так и возбуждающее действие, — ацетилхолин, глутамат.
Классификация торможения
Первичное
Постсинаптическое=гиперполяризационное
Пресинаптическое=деполяризационное
Торможение вслед за перевозбуждением (стрессом) → отключка.
1. Постсинаптическое, гиперполяризационное торможение связано с формированием на мембране ТПСП. Постсинаптическая мембрана при этом гиперполяризуется, возбудимость нейрона понижается, в результате он или уменьшает частоту, или прекращает генерировать ПД.
Виды постсинаптического торможения:
а) Поступательное опережающее торможение по Сеченову: при раздражении ретикулярной формации ствола мозга импульсы по ретикулоспинальному тракту поступают в спинной мозг и активируют тормозные интернейроны, которые тормозят альфа-мотонейроны. Торможение альфа-мотонейронов проявляется в увеличении латентного времени спинальных рефлексов.
б) Поступательное реципрокное торможение по Шеррингтону: при активации рецепторов растяжения какой-либо мышцы возбуждаются ее альфа-мотонейроны, при этом одновременно (реципрокно, сопряженно) тормозятся альфа-мотонейроны мышцы антагониста. Значение: автоматическое облегчение работы сустава.
в) Возвратное торможение по Реншоу: при увеличении частоты импульсации альфа-мотонейрона колатераль его аксона возбуждает тормозный интернейрон (клетка Реншоу). Тормозный нейрон выделяет медиатор глицин, который, вызывая процесс гиперполяризации, снижает величину деполяризации сомы альфа-мотонейрона, что приводит к понижению частоты ПД. Значение: обеспечивает сохранение средней частоты импульсации альфа-мотонейрона, нейрон импульсирует стабильно в течение длительного времени, обеспечивая поддержание полного тонуса.
г) Латеральное (окружающее) торможение: при активации одного нейрона одновременно активируются интернейроны, которые тормозят рядом лежащие нейроны. Это торможение хорошо изучено в сенсорных системах, предполагается, что оно обеспечивает контрастность восприятия сигнала.
2. Пресинаптическое деполяризационное торможение уменьшает эффект действия на клетку возбуждающих синапсов (торможение на входе). Морфологической основой его являются аксо-аксональные синапсы. Медиатор — ГАМК. При выделении ГАМК в пресинаптической терминали возбуждающего синапса развивается длительная деполяризация (до 100 мс), что приводит к уменьшению выделения возбуждающего медиатора и падению амплитуды ВПСП. Ионный механизм пресинаптического торможения до конца не выяснен. Предполагают, что происходит повышение проницаемости для ионов хлора, но двигается он согласно электрохимическому потенциалу из клетки.
Принципы взаимодействия нервных центров. Учение А.А. Ухтомского о доминанте.
Понятие о центральных двигательных программах.
Строение и связи нейронов нервного центра генетически детерминированы. За счет ветвления отростков и установления множества синапсов между клетками они образуют три основных типа нейронных сетей:
1. Иерархическая сеть распространяет афферентную информацию либо к увеличивающемуся число нейронов (принцип дивергенции), либо к меньшему числу нейронов (принцип конвергенции).
2. Локальная сеть – состоит из нейронов с короткими аксонами и обеспечивает взаимосвязь и сохранение информации в пределах одного уровня. Возбуждение в сети может циркулировать по замкнутому кольцу. Дублирование и взаимосвязанность элементов локальной сети обеспечивают надежность нервной регуляции.
3. Дивергентная сеть с одним входом – это нейронный ансамбль, который образует выходные связи с большим количеством разных НЦ. Сеть обеспечивает интеграцию разных рефлекторных актов и общую активность многочисленных нейронов разных отделов мозга.
Доминанта – это господствующий в данный момент очаг возбуждения в нервном центре, обуславливающий работу остальных нервных центров и определяющий направленность поведенческих реакций.
Доминанта — господствующий очаг возбуждения, предопределяющий характер текущих реакций центров в данный момент. Такой доминантный центр может возникнуть в различных этажах ЦНС при достаточно длительном действии гуморальных или рефлекторных раздражителей.
Физиологическую основу доминанты составляют отрицательная индукция и концентрация возбуждения. А сама доминанта является физиологической основой внимания, воли, восприятия и мышления. Принцип доминанты допускает, что если в коре мозга одновременно возникают два очага возбуждения, то один из них оказывается господствующим (доминирующим). Рефлексом, связанным с этим очагом в данный момент, направляется и трансформируется деятельность всего нервного аппарата.
Характеристика:
1. возбудимость обеспечивается таламусом
2. к недоминантным центрам оказывает тормозящее действие (осуществляют ядра таламуса)
3. доминантный очаг обладает стойкостью
4. наличие повышенной возбудимости
5. инерционность, обусловленная длительными следовыми процессами
6. способность к суммации и сопряженному торможению других центров, функционально не совместимых с деятельностью центров доминантного очага.
Способы смены доминанты:
ñ исчезает надобность
ñ появляется более сильная доминанта
Смену осуществляет лобная кора.
Дата добавления: 2018-08-06 ; просмотров: 537 ;
Регуляция нервной деятельности представляет собой процессы возбуждения и торможения в ЦНС. Вначале она возникает как элементарная реакция на раздражение. В процессе эволюции произошло усложнение нейрогуморальных функций, приводящее к образованию основных отделов нервной и эндокринной систем. В данной статье мы изучим один из главных процессов – торможение в ЦНС, виды и механизмы его осуществления.
Нервная ткань, её строение и функции
Одна из разновидностей животных тканей, названная нервной, имеет особое строение, обеспечивающее как процесс возбуждения, так и приводящее в действие функции торможения в ЦНС. Нервные клетки состоят из тела и отростков: коротких (дендритов) и длинного (аксона), который обеспечивает передачу нервных импульсов от одного нейроцита к другому. Окончание аксона нервной клетки контактирует с дендритами следующего нейроцита в местах, называемых синапсами. Они обеспечивают передачу биоэлектрических импульсов по нервной ткани. Причем возбуждение всегда движется в одном направлении – с аксона на тело или дендриты другого нейроцита.
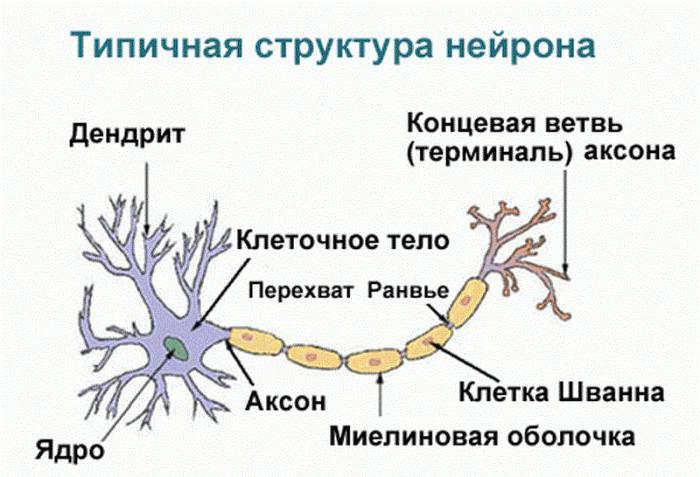
Еще одно свойство, кроме возбуждения, протекающее в нервной ткани, – торможение в ЦНС. Оно является ответной реакцией организма на действие раздражителя, ведущей к снижению или полному прекращению двигательной или секреторной активности, в которой участвуют центробежные нейроны. Торможение в нервной ткани может возникать и без предварительного возбуждения, а только лишь под воздействием тормозного медиатора, например ГАМК. Он является одним из главных трансмиттеров торможения. Здесь же можно назвать такое вещество, как глицин. Эта аминокислота участвует в усилении тормозных процессов и стимулирует в синапсах выработку молекул гаммааминомаслянной кислоты.
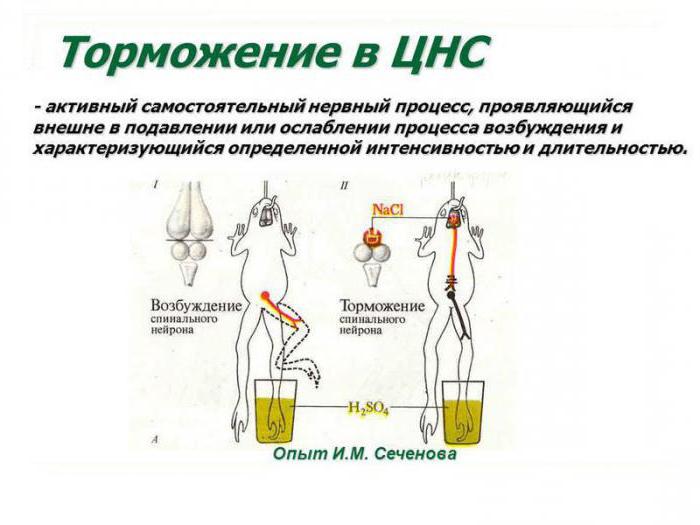
И. М. Сеченов и его работы в нейрофизиологии
Выдающийся российский ученый, создатель теории рефлекторной деятельности головного мозга доказал наличие в центральных отделах нервной системы особых комплексов клеток, способных к инактивации биоэлектрических процессов. Открытие центров торможения в ЦНС стало возможным благодаря применению И. Сеченовым трех видов экспериментов. К ним относятся: перерезание участков коры в различных зонах головного мозга, стимуляция отдельных локусов серого вещества физическими или химическими факторами (электрическим током, раствором хлорида натрия), а также метод физиологического возбуждения мозговых центров. И. М. Сеченов был прекрасным экспериментатором, проводя сверхточные разрезы в зоне между зрительными буграми и непосредственно в самом таламусе лягушки. Он наблюдал уменьшение и полное прекращение двигательной активности конечностей животного.
Взаимодействуют ли нервные процессы?
Исследования выдающихся российских физиологов И. П. Павлова и И. М. Сеченова доказали, что работа центральной нервной системы характеризуется координацией рефлекторных реакций организма. Взаимодействие процессов возбуждения и торможения в ЦНС приводит к согласованной регуляции функций организма: двигательной активности, дыхания, пищеварения, выделения. Биоэлектрические процессы одновременно происходят в нервных центрах и могут последовательно меняться во времени. Это обеспечивает корреляцию и своевременное прохождение ответных рефлексов на сигналы внутренней и внешней среды. Многочисленные опыты, проведенные нейрофизиологами, подтвердили тот факт, что возбуждение и торможение в ЦНС – это ключевые нервные явления, в основе которых лежат некоторые закономерности. Остановимся на них подробнее.
Нервные центры коры головного мозга способны распространять оба вида процессов по всей нервной системе. Это свойство называется иррадиацией возбуждения или торможения. Противоположное явление – уменьшение или ограничение участка мозга, распространяющего биоимпульсы. Оно названо концентрацией. Оба вида взаимодействий ученые наблюдают в течение образования условных двигательных рефлексов. Во время начальной стадии формирования двигательных навыков, вследствие иррадиации возбуждения одновременно сокращаются сразу несколько групп мышц, не обязательно участвующих в выполнении формируемого двигательного акта. Только после многократных повторений формируемого комплекса физических движений (катания на коньках, лыжах, велосипеде), в результате концентрации процессов возбуждения в конкретных нервных очагах коры, все движения человека становятся высококоординированными.

Переключения в работе нервных центров могут происходить также вследствие индукции. Она проявляется при выполнении следующего условия: сначала происходит концентрация торможения или возбуждения, причем эти процессы должны быть достаточной силы. В науке известны два вида индукции: S-фаза (центральное торможение в ЦНС усиливает возбуждение) и отрицательная форма (возбуждение вызывает процесс торможения). Встречается также последовательная индукция. В этом случае нервный процесс меняется на противоположный в самом нервном центре. Исследования нейрофизиологов доказали тот факт, что поведение высших млекопитающих и человека определяется явлениями индукции, иррадиации и концентрации нервных процессов возбуждения и торможения.
Безусловное торможение
Рассмотрим более подробно виды торможения в ЦНС и остановимся на такой его форме, которая присуща как животным, так и человеку. Сам термин был предложен И. Павловым. Ученый считал этот процесс одним из врождённых свойств нервной системы и выделил два его вида: гаснущее и постоянное. Остановимся на них детальнее.
Допустим, в коре существует очаг возбуждения, генерирующий импульсы к рабочему органу (к мышцам, секреторным клеткам желез). Вследствие изменения условий внешней или внутренней среды возникает еще один возбужденный участок коры головного мозга. Он вырабатывает биоэлектрические сигналы большей интенсивности, что тормозит возбуждение в ранее активном нервном центре и его рефлекторной дуге. Гаснущее торможение в ЦНС приводит к тому, что интенсивность ориентировочного рефлекса постепенно уменьшается. Объяснение этому следующее: первичный раздражитель уже не вызывает процесса возбуждения в рецепторах афферентного нейрона.
Другой вид торможения, наблюдаемого как у человека, так и у животных, демонстрирует опыт, проведенный лауреатом нобелевской премии в 1904 году И. П. Павловым. Во время кормления собаки (с выведенной из щеки фистулой) экспериментаторы включали резкий звуковой сигнал – выделение слюны из фистулы прекращалось. Такой вид торможения ученый назвал запредельным.
Являясь врождённым свойством, торможение в ЦНС протекает по безусловно-рефлекторному механизму. Оно достаточно пассивно и не вызывает расхода большого количества энергии, приводя к прекращению условных рефлексов. Постоянное безусловное торможение сопровождает многие психосоматические заболевания: дискинезии, спастический и вялый параличи.
Что такое гаснущий тормоз
Продолжая изучать механизмы торможения в ЦНС, рассмотрим, что представляет собой один из его видов, названный гаснущим тормозом. Хорошо известно, что ориентировочный рефлекс представляет собой реакцию организма на воздействие нового постороннего сигнала. В этом случае в коре мозга образуется нервный центр, находящийся в состоянии возбуждения. Он и формирует рефлекторную дугу, отвечающую за реакцию организма и называемую ориентировочным рефлексом. Этот рефлекторный акт вызывает торможение условного рефлекса, происходящего в данный момент. После многоразового повторения постороннего раздражителя рефлекс, называемый ориентировочным, постепенно снижается и наконец исчезает. А значит, не вызывает больше торможения условного рефлекса. Такой сигнал и получил название гаснущего тормоза.
Таким образом, внешнее торможение условных рефлексов связано с влиянием на организм постороннего сигнала и является врождённым свойством центральной и периферической нервной системы. Внезапный или новый раздражитель, например, болевое ощущение, посторонний звук, изменение освещенности, не только вызывает ориентировочный рефлекс, но также способствует ослаблению или даже полному прекращению условно-рефлекторной дуги, активной в данный момент. Если посторонний сигнал (кроме болевого) действует повторно, торможение условного рефлекса проявляется меньше. Биологическая роль безусловной формы нервного процесса заключается в проведении ответной реакции организма на раздражитель, наиболее важный в данный момент.
Внутреннее торможение
Его другое название, используемое в физиологии высшей нервной деятельности, – условное торможение. Главная предпосылка возникновения такого процесса – отсутствие подкрепления сигналов, поступающих из внешнего мира, врождёнными рефлексами: пищеварительным, слюноотделительным. Возникшие в этих условиях процессы торможения в ЦНС требуют определенного временного интервала. Рассмотрим их виды более подробно.
Например, дифференцировочное торможение возникает как ответ на сигналы окружающей среды, совпадающие по амплитуде, интенсивности и силе к условному раздражителю. Эта форма взаимодействия нервной системы и окружающего мира позволяет организму более тонко различать раздражители и вычленять из их совокупности тот, который получает подкрепление врожденным рефлексом. Например, на звук звонка с силой 15 Гц, подкрепленный кормушкой с пищей, у собаки выработали условную слюноотделительную реакцию. Если к животному применить еще один звуковой сигнал, силой 25 Гц, не подкрепляя его пищей, в первой серии опытов у собаки из фистулы слюна будет выделяться на оба условных раздражителя. Через некоторое время у животного произойдет дифференциация этих сигналов, и на звук, силой 25 Гц слюна из фистулы перестанет выделяться, то есть разовьется дифференцировочное торможение.
Освободить мозг от информации, потерявшей жизненно значимую роль для организма, – эту функцию как раз и выполняет торможение в ЦНС. Физиология опытным путем доказала, что условные двигательные реакции, хорошо закрепленные выработанными навыками, могут сохраняться на протяжении всей жизни человека, например, катание на коньках, езда на велосипеде.
Подводя итог, можно сказать, что процессы торможения в ЦНС – это ослабление или прекращение определенных реакций организма. Они имеют очень большое значение, так как все рефлексы организма корригируются в соответствии с измененными условиями, а если условный сигнал потерял свое значение, то даже полностью могут исчезать. Различные виды торможения в ЦНС являются базовыми для таких способностей психики человека, как сохранение самообладания, различение раздражителей, ожидание.
Запаздывающий вид нервного процесса
Опытным путем можно создать ситуацию, при которой ответ организма на условный сигнал из внешней среды проявляется еще до воздействия безусловного раздражителя, например пищи. При увеличении промежутка времени между началом воздействия условного сигнала (свет, звук, например, удары метронома) и моментом подкрепления до трех минут выделение слюны на вышеназванные условные раздражители все более запаздывает и проявляется только в момент, когда перед животным появляется кормушка с пищей. Отставание ответа на условный сигнал характеризует процессы торможения в ЦНС, названные запаздывающим видом, при котором его время протекания соответствует интервалу запаздывания безусловного раздражителя, например пищи.
Значение торможения в ЦНС
Биологические основы поведения человека
Оба вида активности нервной ткани, как возбуждение, так и торможение в ЦНС, являются основой высшей нервной деятельности. Она обуславливает физиологические механизмы психической деятельности человека. Учение высшей нервной деятельности было сформулировано И. П. Павловым. Современная его трактовка звучит следующим образом:
- Возбуждение и торможение в ЦНС, происходящие во взаимодействии, обеспечивают сложные психические процессы: память, мышление, речь, сознание, а также формируют сложные поведенческие реакции человека.
Чтобы составить научно обоснованный режим учебы, труда, отдыха, ученые применяют знания закономерностей высшей нервной деятельности.
Биологическое значение такого активного нервного процесса, как торможение, можно определить следующим образом. Изменение условий внешней и внутренней среды (отсутствие подкрепления условного сигнала врождённым рефлексом) влечет за собой адекватные изменения приспособительных механизмов в организме человека. Поэтому приобретенный рефлекторный акт угнетается (гаснет) или вовсе исчезает, так как становится для организма нецелесообразным.
Что такое сон?
И. П. Павлов в своих работах экспериментально доказал тот факт, что процессы торможения в ЦНС и сон имеют единую природу. В период бодрствования организма на фоне общей активности коры головного мозга все же диагностируются отдельные её участки, охваченные внутренним торможением. Во время сна оно иррадиирует по всей поверхности больших полушарий, достигая подкорковых образований: зрительных бугров (таламуса), гипоталамуса, ретикулярной формации и лимбической системы. Как указывал выдающийся нейрофизиолог П. К. Анохин, все вышеперечисленные части центральной нервной системы, ответственные за поведенческую сферу, эмоции и инстинкты, во время сна снижают свою активность. Это влечет за собой снижение генерирования нервных импульсов, поступающих из-под корки. Таким образом, активизация коры снижается. Это обеспечивает возможность покоя и восстановления обмена веществ как в нейроцитах большого мозга, так и во всем организме в целом.

Опытами других ученых (Гесса, Экономо) были установлены особые комплексы нервных клеток, входящие в неспецифические ядра зрительных бугров. Процессы возбуждения, диагностируемые в них, вызывают снижение частоты биоритмов коры, которые можно расценивать как переход от активного состояния (бодрствования) ко сну. Исследования таких участков головного мозга, как Сильвиев водопровод и ІІІ желудочек, подтолкнули ученых к идее о наличии центра регуляции сна. Он анатомически связан с участком мозга, ответственным за бодрствование. Поражение этого локуса коры вследствие травмы или в результате наследственных нарушений у человека приводит к патологическим состояниям бессонницы. Также отметим тот факт, что регуляция такого жизненно важного для организма процесса торможения, как сон, осуществляется нервными центрами промежуточного мозга и подкорковых ядер: хвостатого, миндалевидного, ограды и чечевицеобразного.
Читайте также:
