
Тормозные пути центральной нервной системы 1971
Эта книга представляет собой сборник очерков, посвященных разбору идейного наследия Эвальда Васильевича Ильенкова — едва ли не единственного представителя советской философии, идеи которого не только пережили советскую эпоху, но и, судя по тому, что работы его продолжают все активнее переиздаваться. (Подробнее)
Неужели человек — это только электричество между нейронами, биологическая машина, социальное животное? Являются ли наши ценности — вера, любовь, надежда и свобода — всего лишь очень убедительной иллюзией? Неужели современная нейронаука права, и я — это мой мозг?
В своей ломающей стереотипы книге. (Подробнее)
Одни предлагают верить в то, что никакого коронавируса не существует или что его создали искусственно, чтобы обвалить мировую экономику. Другие – что это совершенно невиданный ранее вирус. (Подробнее)
Перед Вами — блистательное исследование чилийских ученых-биологов Умберто Матураны и Франсиско Варелы. В нем представлены основы альтернативной теории познания, главные положения которой противопоставлены классической теории отображения действительности (репрезентационизму).
Сюжетом этой книги. (Подробнее)
В последние 30 лет на передовых рубежах научного знания формируется принципиально новый взгляд на феномен жизни, в рамках которого жизнь предстает как системное явление. Все больше внимания уделяется вопросам, связанным с теорией сложности, с понятиями сетей и моделей организации, что. (Подробнее)
Книга посвящена изучению биосферы и общества как единой системы. В ней дается развернутое изложение мирового эволюционного процесса. Показана общность процессов, протекающих в неживой материи, в биоте и обществе. Значительное место в книге занимает проблема места информатики и вычислительной. (Подробнее)
Т. 1. Процесс производства капитала. 910 с.
Т. 2. Процесс обращения капитала. 648с.
Т. 3 (части 1,2). Процесс капиталистического производства, взятый в целом. 784 с. Издание, подготовленное Ф. Энгельсом. Т. 3, Часть 1, 508с.; Т. 3, Часть 2, 574с.
Эта в своем роде революционно новая книга дает основу для невероятно богатого и глубокого подхода к психике, основанного на объективном знании того, как перевести смысл образов (сновидений, фантазий, образов из фильмов, литературы, искусства и даже последних новостных заголовков) на язык, который. (Подробнее)
Вниманию читателя предлагается книга, в которую включены Конституция СССР (последняя редакция, с изменениями и дополнениями, внесенными Законом СССР от 1 декабря 1988 года), Конституция РСФСР (последняя редакция, с изменениями и дополнениями, внесенными законами РСФСР 1989 и 1990 годов). (Подробнее)
Перед читателями — научный бестселлер американского писателя Джона Дербишира, удостоенный премии имени Эйлера за лучшее популярное изложение математической проблемы. Книга посвящена великой догадке немецкого математика Бернхарда Римана, выдвинутой им в работе «О числе простых чисел, не превышающих. (Подробнее)
| Выберите страну доставки |
Для получения полной информации о книгах
нужно указать страну доставки
Вашего возможного заказа:
Функциональная значимость постсинаптического Т. разнообразна. Афферентное (прямое) Т. служит для ослабления возбуждения функционально антагонистических элементов и тем самым способствует координированному, пространственно направленному протеканию возбуждения в цепях нейронов. В спинном мозге, в частности, такое Т. является основой так называемого реципрокного (взаимообратного) Т. мотонейронов, иннервирующих мышцы-антагонисты (см. Реципрокная иннервация ). Возвратное (коллатеральное) Т., осуществляемое через систему возвратных коллатералей (ветвей) аксонов эфферентных нейронов и специализированных вставочных тормозных нейронов, стабилизирует собственный уровень возбуждения определённого структурно-функционального объединения (блока) нейронов и ограничивает распространение возбуждения на соседние популяции нейронов.
Менее изучено так называемое пресинаптическое Т., выражающееся в угнетении возбуждения в нервных терминалиях, то есть на входе постсинаптического клеточного элемента. Это Т. имеет необычайно большую длительность (сотни мсек ) и совпадает во времени с проявлением деполяризации приходящих афферентов. Предполагают, что на деполяризации основано пресинаптическое Т., а его морфологическим субстратом являются аксо-аксональные синапсы, происхождение пресинаптических элементов которых неизвестно. Имеются веские аргументы в пользу роли гамма-аминомасляной кислоты как медиатора пресинаптического Т., по крайней мере в нервно-мышечных соединениях ракообразных и в спинном мозге позвоночных. По-видимому, сеченовское торможение у лягушки осуществляется по механизму преспнаптического Т. Известно также пессимальное, или вторичное, Т., выражающееся в блокировании возбуждения вследствие его чрезмерности (см. Парабиоз ). Этот феномен, описанный впервые Н. Е. Введенским , трудно выявить при физиологических условиях эксперимента, но можно демонстрировать при аномальных (в частности, судорожных) состояниях.
Изучая условнорефлекторную деятельность, И. П. Павлов выделил внешнее торможение , заключающееся в Т. какой-либо текущей деятельности ориентировочным рефлексом на посторонний раздражитель, и внутреннее торможение , наблюдаемое при угасании условных рефлексов , их дифференцировании, при образовании запаздывающих и следовых условных рефлексов. В особый вид Павлов выделял охранительное Т., предохраняющее нервные центры от чрезмерно сильного раздражения или переутомления. При нарушении взаимоотношений между Т. и возбуждением возникают различные нервные и психических заболевания. См. также Биоэлектрические потенциалы , Высшая нервная деятельность , Гипноз .
Лит.: Экклс Дж.. Физиология синапсов, пер. с англ., М., 1966; Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968; Костюк П. Г., Торможение, в кн.: Общая и частная физиология нервной системы, Л., 1969; Экклс Дж., Тормозные пути центральной нервной системы, пер. с англ., М., 1971.
Л. С. Батуев, Д. Н. Ленков.
Торможе'ние противовключе'нием , торможение электрическое , осуществляемое таким переключением питания обмоток исполнительного электродвигателя, при котором направление тягового усилия изменяется на противоположное. Достигается либо сменой полярности напряжения, подводимого к обмотке вращающегося якоря (ротора) двигателя, либо переключением двух фаз обмотки статора. Величина тормозящего момента регулируется изменением сопротивления в цепи якоря (ротора). При Т. п. сразу же после остановки электропривода он должен быть отключен от сети во избежание изменения направления движения исполнительного двигателя на противоположное. Т. п. применяется в электроприводах грузоподъёмных и транспортных машин, а также прокатных станов и рольгангов.
Координированная деятельность ЦНС - согласованная работа нейронов центральной нервной системы основанная на их взаимодействии между собой.
- осуществляется точное выполнение строго определенной функции (рефлекторного акта);
- согласованная работа центров различных рефлексов, сложная рефлекторная деятельность;
- осуществляется взаимодействие рядом расположенных нервных центров.
Принципы координированной деятельности центральной нервной системы и их нейронные механизмы
Координированная деятельность центральной нервной системы основывается на нескольких принципах:
1. Принцип конвергенции (схождения).При возбуждении большого количества рецепторов импульсы сходятся к одним и тем же нейронам центральной нервной системы. Относительная конвергенция - в спинном и стволовом мозге - конвергенция импульсов от различных рецепторных полей одного и того же рефлекса. Абсолютная конвергенция - в коре головного мозга имеются полимодальные (полисенсорные) нейроны, к ним сходятся импульсы от различных рецепторов.
Значение: обеспечивается центральное облегчение и окклюзия; обеспечивается принцип общего конечного пути.
2. Иррадиация возбуждения - распространение возбуждения на весь нервный центр и другие нервные центры. Он противоположен принципу конвергенции.Причины:
- наличие ветвящихся отростков в пределах центральной нервной системы;
- наличие вставочных нейронов;
- наличие ретикулярной формации.
В ретикулярную формацию поступают импульсы и распространяются по всей коре головного мозга. Распространение возбуждения зависит от силы наносимых раздражений (прямопропорционально), до определенных пределов из-за наличия тормозных нейронов. За счет иррадиации обеспечивается дивергенция (расхождение) возбуждения в центральной нервной системе.
Значение: осуществляется определенная связь между характером ответной реакции центральной нервной системы и силой наносимых раздражений.
3. Принцип реципрокности (сопряжения) - в центральной нервной системе существует взаимосвязь между центрами противоположных рефлексов. Механизм: при возбуждении афферентных нейронов импульсы поступают в центральную нервную систему, там возникают несколько разветвлений: вызывает возбуждение центра мышц сгибателей; образует синапс на тормозной клетке, а она образует синапс на центре мышц разгибателей; идет на симметричную сторону и вызывает противоположные изменения. В результате - повышается тонус мышц сгибателей на стороне раздражения, там же снижается тонус мышц разгибателей, как следствие - сгибание конечности. На противоположной стороне - разгибательный рефлекс. Наблюдается сопряжение между центрами сгибания и разгибания. При возбуждении одного центра центр противоположного рефлекса тормозится. Одновременно наблюдается сопряжение между нервными центрами обеих сторон.
Значение: обеспечивается двигательные реакции, осуществляются взаимодействия между центрами противоположных рефлексов.
4. Принцип доминанты. Доминанта - это преобладающий очаг возбуждения в центральной нервной системе, возникающий под действием сильных и сверхсильных раздражителей.
- доминанта - это нераспространенная форма возбуждения (стационарное возбуждение) - новая форма;
- повышенная возбудимость в очаге доминанты;
- инертность (после прекращения действия раздражения очаг продолжает существовать какое-то время);
- суммация возбуждения и притягивание возбуждения из других центров;
- тормозит деятельность других нервных центров.
Существуют 2 вида доминант:
- экзогенного происхождения - вызвана факторами внешней среды, например: чтение за едой;
- эндогенное происхождение - вызвана факторами внутренней среды.
Значение: обеспечивает внимание, формирование условных рефлексов.
5. Принцип общего конечного пути. При раздражении различных рецепторов в ответную реакцию вовлекаются одни и те же органы. В центральной нервной системе афферентных нейронов больше, чем эфферентных, поэтому от нескольких афферентных нейронов импульсы сходятся к одним и тем же эфферентным.
Значение: за счет небольшого количества рабочих органов организм реагирует на возбуждение большого количества рецепторов; конкуренция раздражителей: эффект некоторых тормозится, а других - усиливается. И победителем является биологически более важный раздражитель.
6. Принцип обратной связи. Обратная связь - поток импульсов от рецепторов в центральную нервную систему, которые несут информацию о происходящем на периферии.
Выделяют 2 вида обратной связи:
- положительная - вызывает усиление ответной реакции;
- отрицательная - вызывает торможение ответной реакции.
Значение: саморегуляция деятельности организма.
Вывод: координированная деятельность центральной нервной системы обеспечивает взаимосвязь в работе нервных центров, за счет этого обеспечивается точное выполнение сложных рефлекторных функций.











- Физиология
![]()
- История физиологии
![]()
- Методы физиологии



Торможение ЦНС
Торможение существует наряду с возбуждением и представляет собой одну из форм деятельности нейрона.
Начало изучения торможения в центральной неравной системе связывают с выходом в свет работы И.М. Сеченова "Рефлексы головного мозга", в которой он показал возможность торможения двигательных рефлексов лягушки при химическом раздражении зрительных бугров головного мозга.
Торможение в центральной нервной системе - активный нервный процесс, проявляющийся в подавлении или ослаблении процесса возбуждения.
Центральное торможение (опыт И.М. Сеченова) - процесс, характеризующийся увеличением времени рефлекса или его полным отсутствием, возникающий при раздражении кристалликом поваренной соли поперечного разреза ствола мозга в области зрительных чертогов.
Классический опыт Сеченова заключается в следующем: у лягушки с перерезанным головным мозгом на уровне зрительных бугров определяли время сгибательного рефлекса при раздражении лапки серной кислотой. После этого на зрительные бугры накладывали кристаллик поваренной соли и снова определяли время рефлекса. Оно постепенно увеличивалось, вплоть до полного исчезновения реакции. После снятия кристаллика соли и промывания мозга физиологическим раствором время рефлекса постепенно восстанавливалось. Это позволило говорить о том, что торможение — активный процесс, возникающий при раздражении определенных отделов ЦНС.
Позже И.М. Сеченовым и его учениками было показано, что торможение в ЦНС может возникнуть при нанесении сильного раздражения на любые афферентные пути.
Периферическое торможение открыто братьями Вебер в 1845 г. Они установили, что раздражение блуждающего нерва тормозит работу сердца до полной его остановки.
Благодаря микроэлектродной технике исследования стало возможным изучение процесса торможения на клеточном уровне.
Различают два вида торможения в зависимости от механизмов его возникновения: деполяризационное и гиперполяризационное. Деполяризационное торможение возникает вследствие длительной деполяризации мембраны, а гиперполяризационное - вследствие гиперполяризации мембраны.
Наступлению деполяризационного торможения предшествует состояние возбуждения. Вследствие длительного раздражения это возбуждение переходит в торможение. В основе возникновения деполяризационного торможения лежит инактивация мембраны для натрия, вследствие чего уменьшаются потенциал действия и его раздражающее влияние на соседние участки, в итоге прекращается проведение возбуждения.
Один из видов этого торможения — пессимальное, описанное Н.Е. Введенским (1886), который показал, что возбуждение может сменяться торможением в любом участке, обладающем низкой лабильностью.
Гиперполяризациоиное торможение осуществляется с участием особых тормозных структур и связано с изменением проницаемости мембраны по отношению к калию и хлору, что вызывает увеличение мембранного и порогового потенциалов, в результате чего становится невозможной ответная реакция.

Центральное торможение (опыт И.М. Сеченова): а — двигательный рефлекс на болевой раздражитель; 6 — раснространснне нервных импульсов от тормозных нейронов ствола мозга к спинному мозгу при наложении кристалла NaCI на область зрительных чертогов и отсутствие двигательного рефлекса на болевой раздражитель

Первичное торможение — процесс активации тормозных нейронов, образующих синаптические связи с клеткой, на которую направлено торможение, при этом данный процесс для клетки является первичным, не связанным с ее предварительным возбуждением.
Вторичное торможение — процесс, который развивается в клетке без участия специфических тормозных структур и является следствием ее собственного возбуждения.
Запредельное торможение - истощение нервных клеток при действии раздражителей высокой интенсивности.
Пессималыюе торможение — блокирование высокочастотных импульсов в немиелинизированных нервных терминалях вследствие их более низкой лабильности.
Пресинаптическое торможение - процесс, реализующийся при активации аксо-аксонального тормозного синапса и блокирующий возбуждающие импульсы, направленные на данную клетку.
Постсинантическое торможение - процесс, развивающийся при активации аксо-соматических и аксо-дендритических тормозных синапсов и локализующийся на собственной мембране клетки, на которую направлено торможение.
Рецинрокное торможение — взаимное подавление активности антагонистических нервных структур.
Афферентное коллатеральное торможение - частный случай реципрокного торможения, локализуемый в афферентной части рефлекторной дуги.
Эфферентное коллатеральное (возвратное) торможение — процесс, при котором тормозные вставочные нейроны действуют на те же нервные клетки, которые их активировали, при этом торможение тем сильнее, чем интенсивнее предшествующее возбуждение.
Латеральное торможение — процесс, при котором вставочные тормозные нейроны подавляют активность не только клетки, которая их инициировала, но и других, рядом расположенных.

Латеральное торможение (Т — тормозной нейрон)

Возвратное торможение (Т-тормозной вставочный нейрон (клетка Реншоу); М — мотонейрон)

Рецинрокное торможение (Т — тормозной вставочный нейрон (клетка Реншоу); М — мотонейрон)

Поступательное торможение (Т — тормозной нейрон)
Процессы торможения в центральной нервной системе
Процессы возбуждения и торможения в нервной системе тесно взаимосвязаны.
Торможение в ЦНС способствует определенной координации выполняемой функции. При этом блокируется деятельность нейронов и центров, которые в данный момент не требуются для выполнения приспособительной реакции. Кроме того, торможение выполняет и защитную функцию, предохраняя нервные клетки от перевозбуждения и истощения при действии сильных раздражителей.
Различают несколько видов торможения в нервной системе.
Постсипаптическое торможение развивается в случаях, когда тормозной медиатор, выделяемый нервным окончанием, изменяет свойства постсинаптической мембраны таким образом, что нервная клетка не может генерировать потенциал действия. Постсипаптическое торможение может быть обусловлено длительной деполяризацией или гиперполяризацией, возникающей в постсинаптической мембране вследствие взаимодействия медиатора с рецепторами, открывающими калиевые и хлорные каналы. Наиболее распространенными тормозными медиаторами являются гамма-аминомасляная кислота и глицин. Глицин выделяется особыми тормозными клетками (клетки Реншоу) в синапсах, образуемых этими клетками на мембране другого нейрона. Действуя на рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для ионов СI-, при этом ионы хлора поступают в клетку согласно концентрационному градиенту, в результате чего развивается гиперполяризация. При действии гамма-аминомасляной кислоты на постсинаптическую мембрану постсинаптическое торможение развивается в результате входа ионов хлора в клетку или выхода ионов калия из клетки. Концентрационные градиенты ионов К + в процессе развития торможения нейронов поддерживается Na + /К + -насосом, а ионов СI - — СI - -насосом.
Возвратное постсинаптическое торможение - это такое торможение, при котором тормозные вставочные нейроны (клетки Реншоу) действуют на те же нервные клетки, которые их иннервируют. Примером возвратного постсинаптического торможения может служить торможение в мотонейронах спинного мозга. Этот вид торможения обеспечивает, например, поочередное сокращение и расслабление скелетных мышц — сгибателей и разгибателей, что необходимо для координации движений конечностей при ходьбе.
Латеральное постсинаптическое торможение обусловлено тем, что тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями. В результате в этих соседних клетках развивается очень глубокое торможение, называемое латеральным, так как образующаяся зона торможения находится сбоку по отношению к возбужденному нейрону и инициируется им.
Реципрокное торможение, примером которого является торможение нервных центров мышц-антагонистов, заключается в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны. Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей. Если бы возбуждались одновременно центры мышц-сгибателей и мышц- разгибателей, сгибание конечности в суставе было бы невозможно.
Пресинаптическое торможение связано с тем, что в пресинаптическом окончании может развиваться продолжительная деполяризация мембраны, которая приводит к развитию торможения. В очаге деполяризации нарушается процесс распространения возбуждения и импульсы не могут пройти через зону деполяризации. Следовательно, не происходит выделения медиатора в синаптическую щель в достаточном количестве и не возбуждается постсинаптический нейрон. В ЦНС имеется огромное число тормозных нейронов, в частности клетки Реншоу. Эти тормозные нейроны синтезируют специфические тормозные медиаторы и осуществляют реакцию торможения. Активация тормозного нейрона вызывает деполяризацию мембраны терминалей в афферентных нейронах, что затрудняет процесс проведения потенциала действия. Медиатором втакихаксо аксональных синапсах служит гамма-аминомасляная кислота или другой тормозной медиатор. Деполяризация является следствием повышения проницаемости мембраны для ионов хлора, в результате эти ионы выходят из клетки.
РЕЦИПРОКНОСТЬ (лат. reciprocus — возвращающийся, обратный, взаимный) — один из физиологических механизмов координации деятельности нервных центров, обеспечивающий взаимное, противоположно направленное регулирующее влияние на функции органов и тканей.
Впервые реципрокные отношения между мышцей сердца и мышечными элементами стенки артерий были описаны в 1866 г. И. Ф. Ционом; Э. Геринг и Брейер (J. Breuer, 1868) показали Р. в работе дыхательного центра; Мельтцер и Кронеккер (S. J. Meltzer, H. Kronecker, 1883) открыли реципрокное торможение при осуществлении акта глотания, а H. Е. Введенский (1896) установил
Р. на уровне коры головного мозга. Механизм реципрокной иннервации скелетных мышц был впервые объяснен учеником И. М. Сеченова П. А. Спиро (1874) и детально проанализирован английским физиологом Ч. Шеррингтоном (1906).
Наиболее изученной является Р. на уровне спинного мозга (см.), обеспечивающая взаимно противоположную деятельность скелетных мышц. При осуществлении организмом строго координированных двигательных актов (ходьба, бег, плавание, чесание, глотание и др.) спинальные центры мышц-антагонистов находятся в противоположных функциональных состояниях (см. Движения). Напр., возбуждение группы мотонейронов, вызывающих сокращение мышц-сгибателей, сопровождается реципрокным торможением другой группы мотонейронов, что приводит к расслаблению мышц-разгибателей.
Прямое исследование процессов возбуждения (см.) и торможения (см.) в одиночных нервных клетках, осуществленное с помощью микроэлектродов, позволило объяснить нейрофизиологический механизм Р. Показано, в частности, что афферентные нервные волокна, идущие от проприоцепторов скелетных мышц, сигнализируют в спинной мозг о степени их растяжения и по одной ветви аксона вызывают возбуждение мотонейронов, приводящее к сокращению мышц-разгибателей. Вторая веточка аксона сенсорного нейрона через вставочный тормозной нейрон одновременно тормозит мотонейроны, обеспечивающие иннервацию мышц-сгибателей сустава. Подобный механизм Р. описан в других структурах ц. н. с.: таламусе, гиппокампе, ретикулярной формации, коленчатых телах, мозжечке и коре больших полушарий. Так, механизм смены вдоха и выдоха включает Р. между инспираторным и экспираторным центрами. Р. характерна для центров голода и насыщения, а также для прессорных и депрессорных отделов сосудодвигательного центра.
Нарушения Р. чаще всего наблюдаются при заболеваниях, связанных с отравлением токсическими препаратами (напр., стрихнином) или с инфекционными заболеваниями (напр., столбняком), к-рые сопровождаются функциональными нарушениями ц. н. с.
Библиография: Бериташвили И. С. Общая физиология мышечной и нервной системы, т. 2, М., 1966; Гранит Р. Основы регуляции движений, пер. с англ., М., 1973; Шеррингтон Ч. Интегративная деятельность нервной системы, пер. с англ., JI., 1969; Экклс Д ж. Тормозные пути центральной нервной системы, пер. с англ., М., 1971, библиогр.; С у о n E. u. Ludwig С. F. W. Die Reflexe eines der sensiblen Nerven des Herzens auf die motorischen der Blutgefasse, Arb. physiol. Anst. Lpz., Bd 1, S. 128, 1867.
Теория по нормальной физиологии: Координация и торможение. Принципы координации, классификация торможения в ЦНС, роль торможения…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Координация
Координация — это оптимальное взаимодействие центров, направленное на достижение полезного результата.
Иррадиация возбуждения — распространение возбуждения с одного нервного центра на другие.
- направленная (из центра в конкретно другой),
- генерализованная (возбуждение всех нервных центров ЦНС)
Иррадиация зависит от силы раздражителя, в норме иррадиация возникает при действии сильного раздражителя.
Возбудимость ЦНС — влияет на иррадиацию.
Дивергенция — возбуждение расходится.
Соотношение чувствительных и двигательных нейронов: 10:1. В основе этого принципа лежит конвергентная сеть — воронка Ч. Шеррингтона.
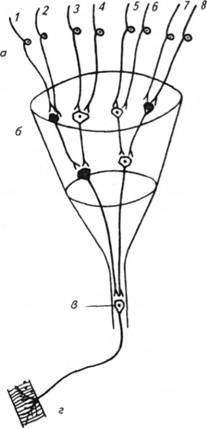
Независимо от рецептора, который раздражается, все сводится к одному.
Чем выше уровень организации ЦНС, тем больше рефлекторных центров подчиняется коре больших полушарий.
При этом спинальные двигательные программы не могут функционировать самостоятельно, но с помощью коры больших полушарий вовлекается в разнообразные формы двигательной активности.
Нижележащие отделы подчиняются вышележащим.
Влияние вышележащих центров настолько велико, что при прекращении импульсов от головного и спинного мозга может наблюдаться явление спинального шока — это обратимое выключение спинальных рефлексов.
Обратный поток афферентных импульсов, возникающих в организме в результате деятельности органов и тканей, получили название вторичных афферентных импульсов .
Различают 2 вида обратной связи:
- Положительная обратная связь — усиливает.
- Отрицательная обратная связь — уменьшает.
Принцип обратной связи характерна и для гуморальной регуляции.
ЦНС — очень надежная система. Даже при потери большого количества нейронов может быть скомпенсирована оставшимися — произойдет компенсация функций.
В основе надежной работы лежит:
- избыточность элементов — резервирование;
- взаимозаменяемость и дублирование функций;
- пластичность и обучаемость;
- повышение гуморальной чувствительности при денервации органа.
При действии адекватного стимула рефлекторная реакция может состояться (разное время произошедшего стимула и т.д.), но может и не состояться.
В каждый данный момент времени в ЦНС присутствует определяющий (доминантный) очаг возбуждения, подчиняющий себе деятельность всей нервной системы и определяющий характер приспособительной реакции.
Таким образом, создаются определенные условия для реакции на раздражитель, имеющий наибольшее биологическое значение.
Свойства доминантного центра:
- повышенная возбудимость и устойчивость возбуждения;
- способность к суммации возбуждений;
- торможение других нервных центров.
Функция доминантного центра: осуществление поведенческой реакции для удовлетворения соответствующей потребности.
При возбуждении одних нервных центров деятельность других может затормаживаться.
Торможение — это активный процесс, результатом которого является прекращение или ослабление возбуждения.
Классификация торможения в ЦНС:
- Первичное:
- постсинаптическое: прямое, возвратное, реципрокное, латериальное,
- пресинаптическое.
- Вторичное:
- торможение вслед за возбуждением,
- пессимум Введенского.
Первичное торможение идет с участием тормозных структур : тормозной нейрон, тормозной синапс, тормозной медиатор.
Вторичное — без участия тормозных структур на самих возбуждающих клетках.
Механизм постсинаптического торможения — тормозные клетки возбуждаются и выделяют тормозной медиатор (ГАМК или глицин); на постсинаптической мембране возникает гиперполяризация ПСП.
Постсинаптическое:
Прямое торможение — тормозные клетки получают возбуждение напрямую от чувствительного нейрона.
Возвратное торможение — это самоторможение, когда тормозная клетка возбуждается от коллатерального затормаживаемого нейрона; функция — ограничение перевозбуждения.
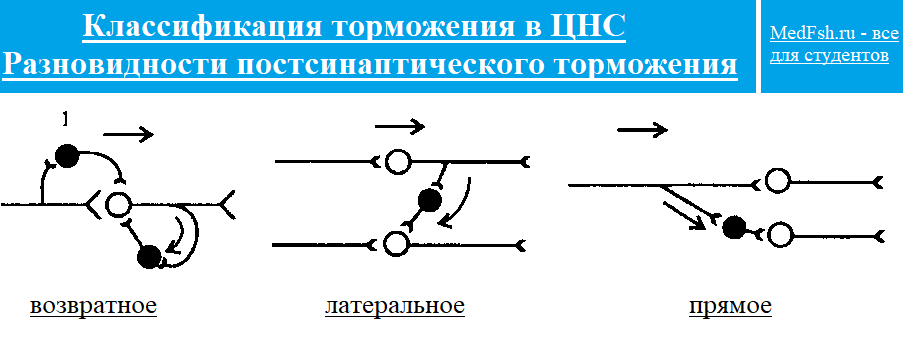
Реципрокное торможение
Возбуждение одного центра сопровождается торможением другого центра, осуществляющего антагонистический рефлекс (работа мышц антагонистов и принцип координации нервных центров). При активации одной мышцы, например сгибателя, от афферентного нейрона импульс идет к тормозному нейрону, который затормаживает мотонейрон мышцы антагониста — разгибателя.
Латеральное торможение — это торможение соседнего нейрона или соседнего нервного центра. Данный вид торможения увеличивает контрастность восприятия от рецепторов.
Пресинаптическое торможение — в основе лежат аксоаксональные синапсы в ядрах тройничного нерва, в ядрах таламуса.
Тормозная клетка возбуждается и выделяет тормозной медиатор . Взаимодействие медиатора с рецепторами на мембране приводит к стойкой длительной деполяризации мембраны аксона; возбудимость и проводимость в этом участке аксона уменьшается.
Отличия от постсинаптического: открытие Na и Cl каналов, которые увеличивают стойкость.
Значение пресинаптического торможения — регулирует приток сенсорных импульсов, блокирует слабые, незначительные сигналы.
Вторичное торможение — торможение вслед за возбуждением возникает из-за выраженной фазы следовой гиперполяризации.
Пессимальное торможение — возникает при очень высокой частоте приходящих импульсов.
Читайте также:
