
В коре больших полушарий содержится свыше 14 млрд нервных клеток

Кора головного мозга насчитывает более 15 миллиардов нервных клеток и волокон. Каждая из них имеет разную форму, и образуют несколько уникальных слоев, отвечающих за определенные функции. К примеру, функциональность клеток второго и третьего слоя заключается в трансформации возбуждения и правильное перенаправление в определенные отделы головного мозга. А, например, центробежные импульсы представляют собой работоспособность пятого слоя. Рассмотрим каждый слой более тщательно.
Нумерация слоев головного мозга начинается от поверхности и идет глубже:
Кора мозга состоит из нервных клеток, их отростков и нейроглии. У взрослого человека толщина коры в большинстве областей составляет около 3 мм. Площадь коры больших полушарий благодаря многочисленным складкам и бороздам составляет 2500 см2. Для большинства участков коры головного мозга характерно шестислойное расположение нейронов*(1). Кора больших полушарий состоит из 14—17 млрд. клеток. Клеточные структуры коры головного мозга представленыпирамидными, веретенообразными и звездчатыми нейронами.
Звездчатые клеткивыполняют главным образом афферентную функцию.Пирамидные и веретенообразные клетки— это преимущественно эфферентные нейроны.
В коре больших полушарий имеются высокоспециализированные нервные клетки, воспринимающие афферентные импульсы от определенных рецепторов (например, от зрительных, слуховых, тактильных и т. д.). Имеются также нейроны, которые возбуждаются нервными импульсами, идущими от разных рецепторов организма. Это так называемые полисенсорные нейроны.
Отростки нервных клеток коры головного мозга связывают ее различные отделы между собой или устанавливают контакты коры больших полушарий с нижележащими отделами центральной нервной системы. Отростки нервных клеток, соединяющие между собой различные участки одного и того же полушария называются ассоциативными, связывающие чаще всего одинаковые участки двух полушарий —комиссуральнымии обеспечивающие контакты коры головного мозга с другими отделами центральной нервной системы и через них со всеми органами и тканями тела —проводящими(центробежными). Схема этих путей приведена на рисунке.
Функции коры головного мозга.
1) Кора головного мозга осуществляет взаимодействие организма с окружающей средой за счет безусловных и условных рефлексов;
2) она является основой высшей нервной деятельности (поведения) организма;
3) за счет деятельности коры головного мозга осуществляются высшие психические функции: мышление и сознание;
4) кора головного мозга регулирует и объединяет работу всех внутренних органов и регулирует такие интимные процессы, как обмен веществ.
Таким образом, с появлением коры головного мозга она начинает контролировать все процессы, протекающие в организме, а также всю деятельность человека, т.
Кора́ больших полуша́рий головного мозга
Слой серого вещества толщиной 1—5 мм, покрывающий полушария большого мозга млекопитающих животных и человека. Эта часть головного мозга (См. Головной мозг), развившаяся на поздних этапах эволюции животного мира, играет исключительно важную роль в осуществлении психической, или высшей нервной деятельности (См. Высшая нервная деятельность), хотя эта деятельность является результатом работы мозга как единого целого. Благодаря двусторонним связям с нижележащими отделами нервной системы, кора может участвовать в регуляции и координации всех функций организма. У человека кора составляет в среднем 44% от объёма всего полушария в целом. Её поверхность достигает 1468—1670 см 2 .
Строение коры. Характерной особенностью строения коры является ориентированное, горизонтально-вертикальное распределение составляющих её нервных клеток по слоям и колонкам; таким образом, корковая структура отличается пространственно упорядоченным расположением функционирующих единиц и связей между ними (рис. 1). Пространство между телами и отростками нервных клеток коры заполнено нейроглией (См. Нейроглия) и сосудистой сетью (капиллярами). Нейроны коры подразделяются на 3 основных типа: пирамидные (80—90% всех клеток коры), звездчатые и веретенообразные. Основные функциональный элемент коры — афферентно-эфферентный (т. е. воспринимающий центростремительные и посылающий центробежные стимулы) длинноаксонный пирамидный нейрон (рис. 2). Звездчатые клетки отличаются слабым развитием Дендритов и мощным развитием Аксонов, которые не выходят за пределы поперечника коры и охватывают своими разветвлениями группы пирамидных клеток. Звездчатые клетки выполняют роль воспринимающих и синхронизирующих элементов, способных координировать (одновременно тормозить или возбуждать) пространственно близкие группы пирамидных нейронов. Корковый нейрон характеризуется сложным субмикроскопическим строением (см. Клетка). Различные по топографии участки коры отличаются плотностью расположения клеток, их величиной и другими характеристиками послойной и колончатой структуры. Все эти показатели определяют архитектуру коры, или её цитоархитектонику (см. рис. 1 и 3).
Наиболее крупные подразделения территории коры — древняя (палеокортекс), старая (архикортекс), новая (неокортекс) и межуточная кора. Поверхность новой коры у человека занимает 95,6%, старой 2,2%, древней 0,6%, межуточной 1,6%.
Если представить себе кору мозга в виде единого покрова (плаща), одевающего поверхность полушарий, то основная центральная часть его составит новая кора, в то время как древняя, старая и межуточная займут место на периферии, т. е. по краям этого плаща. Древняя кора у человека и высших млекопитающих состоит из одного клеточного слоя, нечетко отделённого от нижележащих подкорковых ядер; старая кора полностью отделена от последних и представлена 2—3 слоями; новая кора состоит, как правило, из 6—7 слоев клеток; межуточные формации — переходные структуры между полями старой и новой коры, а также древней и новой коры — из 4—5 слоев клеток. Неокортекс подразделяется на следующие области: прецентральную, постцентральную, височную, нижнетеменную, верхнетеменную, височно-теменно-затылочную, затылочную, островковую и лимбическую. В свою очередь, области подразделяются на подобласти и поля. Основной тип прямых и обратных связей новой коры — вертикальные пучки волокон, приносящие информацию из подкорковых структур к коре и посылающие её от коры в эти же подкорковые образования. Наряду с вертикальными связями имеются внутрикортикальные — горизонтальные — пучки ассоциативных волокон, проходящие на различных уровнях коры и в белом веществе под корой. Горизонтальные пучки наиболее характерны для I и III слоев коры, а в некоторых полях для V слоя. Горизонтальные пучки обеспечивают обмен информацией как между полями, расположенными на соседних извилинах, так и между отдалёнными участками коры (например, лобной и затылочной).
Функциональные особенности коры обусловливаются упомянутым выше распределением нервных клеток и их связей по слоям и колонкам. На корковые нейроны возможна конвергенция (схождение) импульсов от различных органов чувств. Согласно современным представлениям, подобная конвергенция разнородных возбуждений — нейрофизиологический механизм интегративной деятельности головного мозга, т. е. анализа и синтеза ответной деятельности организма. Существенное значение имеет и то, что нейроны сведены в комплексы, по-видимому, реализующие результаты конвергенции возбуждений на отдельные нейроны. Одна из основных морфо-функциональных единиц коры — комплекс, называемый колонкой клеток, который проходит через все корковые слои и состоит из клеток, расположенных на одном перпендикуляре к поверхности коры. Клетки в колонке тесно связаны между собой и получают общую афферентную веточку из подкорки. Каждая колонка клеток отвечает за восприятие преимущественно одного вида чувствительности. Например, если в корковом конце кожного анализатора (См. Кожный анализатор) одна из колонок реагирует на прикосновение к коже, то другая — на движение конечности в суставе. В зрительном анализаторе (См. Зрительный анализатор) функции восприятия зрительных образов также распределены по колонкам. Например, одна из колонок воспринимает движение предмета в горизонтальной плоскости, соседняя — в вертикальной и т. п.
Локализация функций в коре характеризуется динамичностью в силу того, что, с одной стороны, имеются строго локализованные и пространственно отграниченные зоны коры, связанные с восприятием информации от определенного органа чувств, а с другой — кора является единым аппаратом, в котором отдельные структуры тесно связаны и в случае необходимости могут взаимозаменяться (т. н. пластичность корковых функций). Кроме того, в каждый данный момент корковые структуры (нейроны, поля, области) могут образовывать согласованно действующие комплексы, состав которых изменяется в зависимости от специфических и неспецифических стимулов, определяющих распределение торможения (См. Торможение) и возбуждения (См. Возбуждение) в коре. Наконец, существует тесная взаимозависимость между функциональным состоянием корковых зон и деятельностью подкорковых структур. Территории коры резко различаются по своим функциям. Большая часть древней коры входит в систему обонятельного анализатора. Старая и межуточная кора, будучи тесно связанными с древней корой как системами связей, так и эволюционно, не имеют прямого отношения к обонянию. Они входят в состав системы, ведающей регуляцией вегетативных реакций и эмоциональных состояний организма (см. Ретикулярная формация, Лимбическая система). Новая кора — совокупность конечных звеньев различных воспринимающих (сенсорных) систем (корковых концов Анализаторов).
Кроме того, локализация функций в первичных зонах многократно дублируется по механизму, напоминающему голографию (См. Голография), когда каждый самый маленький участок запоминающего устройства содержит сведения о всём объекте. Поэтому достаточно сохранности небольшого участка первичного сенсорного поля, чтобы способность к восприятию почти полностью сохранилась. Вторичные поля получают проекции от органов чувств через дополнительные переключения в подкорке, что позволяет производить более сложный анализ того или иного образа. Наконец, третичные поля, или ассоциативные зоны, получают информацию от неспецифических подкорковых ядер, в которых суммируется информация от нескольких органов чувств, что позволяет анализировать и интегрировать тот или иной объект в ещё более абстрагированной и обобщённой форме. Эти области называются также зонами перекрытия анализаторов. Первичные и отчасти вторичные поля — возможный субстрат первой сигнальной системы (См. Первая сигнальная система), а третичные зоны (ассоциативные) — второй сигнальной системы (См. Вторая сигнальная система), специфичной для человека (И. П. Павлов). Эти межанализаторные структуры определяют сложные формы мозговой деятельности, включающие и профессиональные навыки (нижнетеменная область), и мышление, планирование и целенаправленность действий (лобная область), и письменную и устную речь (нижняя лобная подобласть, височная, височно-теменно-затылочная и нижнетеменная области). Основные представители первичных зон в затылочной области — поле 17, где спроецирована сетчатка, в височной — поле 41, где спроецирован Кортиев орган, в прецентральной области — поле 4, где осуществляется проекция проприорецепторов в соответствии с расположением мускулатуры, в постцентральной — поля 3 и 1, где спроецированы экстерорецепторы в соответствии с их распределением в коже. Вторичные зоны представлены полями 8 и 6 (Двигательный анализатор), 5 и 7 (кожный анализатор), 18 и 19 (зрительный анализатор), 22 (Слуховой анализатор). Третичные зоны представлены обширными участками лобной области (поля 9, 10, 45, 44 и 46), нижнетеменной (поля 40 и 39), височно-теменно-затылочной (поле 37).
Корковые структуры играют первостепенную роль в обучении животных и человека. Однако образование некоторых простых условных рефлексов (См. Условные рефлексы), главным образом с внутренних органов, может быть обеспечено подкорковыми механизмами. Эти рефлексы могут образовываться и на низших уровнях развития, когда ещё нет коры. Сложные условные рефлексы, лежащие в основе целостных актов поведения (См. Поведение), требуют сохранности корковых структур и участия не только первичных зон корковых концов анализаторов, но и ассоциативных — третичных зон. Корковые структуры имеют прямое отношение и к механизмам памяти (См. Память). Электрораздражение отдельных областей коры (например, височной) вызывает у людей сложные картины воспоминаний.
Характерная особенность деятельности коры — её спонтанная электрическая активность, регистрируемая в виде электроэнцефалограммы (ЭЭГ). В целом кора и её нейроны обладают ритмической активностью, которая отражает происходящие в них биохимические и биофизические процессы. Эта активность имеет разнообразную амплитуду и частоту (от 1 до 60 гц) и изменяется под влиянием различных факторов.
Ритмическая активность коры нерегулярна, однако можно по частоте потенциалов выделить несколько разных типов её (альфа-, бета-, дельта- и тета-ритмы). ЭЭГ претерпевает характерные изменения при многих физиологических и патологических состояниях (различных фазах сна (См. Сон), при опухолях, судорожных припадках и т. и.). Ритм, т. е. частота, и амплитуда биоэлектрических потенциалов (См. Биоэлектрические потенциалы) коры задаются подкорковыми структурами, которые синхронизируют работу групп корковых нейронов, что и создаёт условия для их согласованных разрядов. Этот ритм связан с апикальными (верхушечными) дендритами пирамидных клеток. На ритмическую деятельность коры накладываются влияния, идущие от органов чувств. Так, вспышка света, щелчок или прикосновение к коже вызывают в соответствующих зонах т. н. первичный ответ, состоящий из ряда позитивных волн (отклонение электронного луча на экране осциллографа вниз) и негативной волны (отклонение луча вверх). Эти волны отражают деятельность структур данного участка коры и меняются в её различных слоях.
Филогенез и онтогенез коры. Кора — продукт длительного эволюционного развития, в процессе которого сначала появляется древняя кора, возникающая в связи с развитием обонятельного анализатора у рыб. С выходом животных из воды на сушу начинает интенсивно развиваться т. н. плащевидная, полностью обособленная от подкорки часть коры, которая состоит из старой и новой коры. Становление этих структур в процессе приспособления к сложным и разнообразным условиям наземного существования связано (совершенствованием и взаимодействием различных воспринимающих и двигательных систем. У земноводных кора представлена древней и зачатком старой коры, у пресмыкающихся хорошо развиты древняя и старая кора и появляется зачаток новой коры. Наибольшего развития новая кора достигает у млекопитающих, а среди них у приматов (обезьяны и человек), хоботных (слоны) и китообразных (дельфины, киты). В связи с неравномерностью роста отдельных структур новой коры её поверхность становится складчатой, покрываясь бороздами и извилинами. Совершенствование коры конечного мозга у млекопитающих неразрывно связано с эволюцией всех отделов центральной нервной системы. Этот процесс сопровождается интенсивным ростом прямых и обратных связей, соединяющих корковые и подкорковые структуры. Т. о., на более высоких этапах эволюции функции подкорковых образований начинают контролироваться корковыми структурами. Данное явление получило название кортиколизации функций. В результате кортиколизации ствол мозга образует с корковыми структурами единый комплекс, а повреждение коры на высших этапах эволюции приводит к нарушению жизненно важных функций организма. Наибольшие изменения и увеличение в процессе эволюции новой коры претерпевают ассоциативные зоны, в то время как первичные, сенсорные поля уменьшаются по относительной величине. Разрастание новой коры приводит к вытеснению старой и древней на нижнюю и срединную поверхности мозга.
Корковая пластинка появляется в процессе внутриутробного развития человека сравнительно рано — на 2-м месяце. Раньше всего выделяются нижние слои коры (VI—VII), затем — более высоко расположенные (V, IV, III и II; см. рис. 1). К 6 месяцам у эмбриона уже имеются все цитоархитектонические поля коры, свойственные взрослому человеку. После рождения в росте коры можно выделить три переломных этапа: на 2—3-м месяце жизни, в 2,5—3 года и в 7 лет. К последнему сроку цитоархитектоника коры полностью сформирована, хотя тела нейронов продолжают увеличиваться до 18 лет. Корковые зоны анализаторов завершают своё развитие раньше, и степень их увеличения меньше, чем у вторичных и третичных зон. Отмечается большое разнообразие в сроках созревания корковых структур у разных индивидуумов, что совпадает с разнообразием сроков созревания функциональных особенностей коры. Т. о., индивидуальное (Онтогенез) и историческое (Филогенез) развитие коры характеризуется сходными закономерностями.
Лит.: Орбели Л. А., Вопросы высшей нервной деятельности, М.— Л., 1949; Цитоархитектоника коры большого мозга человека. Сб. ст., М., 1949; Филимонов И. Н., Сравнительная анатомия коры большого мозга млекопитающих, М., 1949; Павлов И. П., Двадцатилетний опыт объективного изучения высшей нервной деятельности животных, Полн. собр. соч., 2 изд., т. 3, кн. 1—2, М., 1951; Брейзье М., Электрическая активность нервной системы, пер. с англ., М., 1955; Сепп Е. К., История развития нервной системы позвоночных, 2 изд., М., 1959; Лурия А. Р., Высшие корковые функции человека и их нарушения при локальных поражениях мозга, М., 1962; Воронин Л. Г., Курс лекций по физиологии высшей нервной деятельности, М., 1965; Поляков Г. И., О принципах нейронной организации мозга, М., 1965; Корковая регуляция деятельности подкорковых образований головного мозга. Сб. ст., Тб., 1968; Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968; Беритов И. С., Структура и функции коры большого мозга, М., 1969.
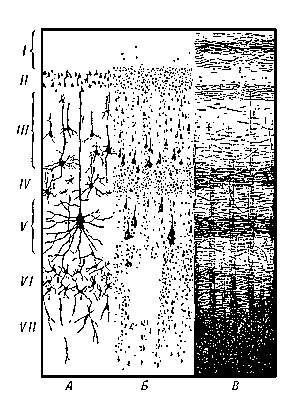
Рис. 1. Схема строения коры головного мозга человека: I — зональный слой, II — наружный зернистый слой, III — пирамидный слой, IV — внутренний зернистый слой, V — ганглионарный слой, VI — слой треугольных клеток, VII — слой веретеновидных клеток. А — нейронное строение, Б — цитоархитектоника, В — волоконная структура.
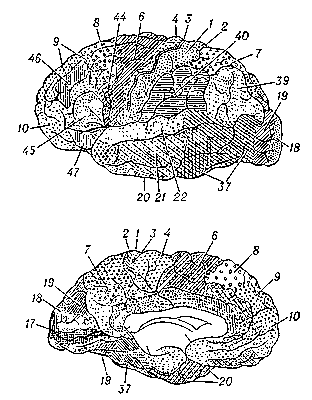
Рис. 3. Карта цитоархитектонических полей коры головного мозга человека: А — наружная поверхность полушария, Б — внутренняя поверхность полушария. Номерами и различной штриховкой обозначены цитоархитектонические поля коры.
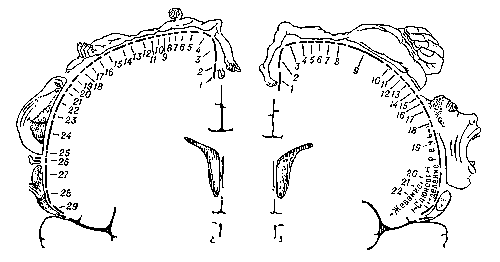
Рис. 4. Представительство чувствительных функций тела в задней центральной извилине (А) и двигательных функций — в передней центральной извилине (Б). А: 1 — половые органы; 2 — пальцы; 3 — ступня; 4 — голень; 5 — бедро; 6 — туловище; 7 — шея; 8 — голова; 9 — плечо; 10 — рука; 11 — локоть; 12 — предплечье; 13 — запястье; 14 — кисть; 15 — мизинец; 16 — безымянный палец; 17 — средний палец; 18 — указательный палец; 19 — большой палец; 20 — глаз; 21 — нос; 22 — лицо; 23 — верхняя губа; 24 — губы; 25 — нижняя губа; 26 — зубы, дёсны и челюсть; 27 — язык; 28 — глотка; 29 — внутренние органы. Б: 1 — пальцы; 2 — лодыжка; 3 — колено; 4 — бедро; 5 — туловище; 6 — плечо; 7 — локоть; 8 — запястье; 9 — кисть; 10 — мизинец; 11 — безымянный палец; 12 — средний палец; 13 — указательный палец; 14 — большой палец; 15 — шея; 16 — бровь; 17 — веко и глазное яблоко; 18 — лицо; 19 — губы; 20 — челюсть; 21 — язык; 22 — глотание. Размеры частей тела, изображенного на рисунке, соответствуют представительству двигательных и чувствительных функций организма в передней и задней центральных извилинах коры.
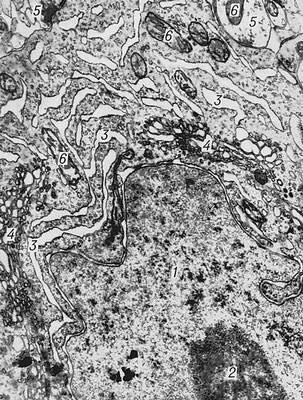
Рис. 2. Электронномикроскопическое строение пирамидной клетки коры головного мозга белой крысы: 1 — ядро; 2 — ядрышко; 3 — канальцы эндоплазматической сети; 4 — комплекс Гольджи; 5 — синапсы; 6 — митохондрии.
Кора больших полушарий является высшим отделом центральной нервной системы, который в процессе филогенетического развития появляется позже всего и формируется в ходе индивидуального (онтогенетического) развития позже других отделов мозга. Кора представляет собой слой серого вещества толщиной 2—3 мм, содержащий в среднем около 14 млрд. (от 10 до 18 млрд.) нервных клеток, нервные волокна и межуточную ткань (нейроглию). На поперечном ее срезе по расположению нейрон ов и их связей различают 6 горизонтальных слоев. Благодаря многочисленным извилинам и бороздам площадь поверхности коры достигает 0,2 м2. Непосредственно под корой находится белое вещество, состоящее из нервных волокон, которые передают возбуждение в кору и из нее, а также от одних участков коры другим.
Корковые нейрон ы и их связи. Несмотря на огромное число нейрон ов в коре, известно очень немного их разновидностей. Основными типами их являются пирамидные и звездчатые нейрон ы.
.
В афферентной функции коры и в процессах переключения возбуждения на соседние нейрон ы основная роль принадлежит звездчатым нейрон ам. Они составляют у человека более половины всех клеток коры. Эти клетки имеют короткие ветвящиеся аксон ы, не выходящие за пределы серого вещества коры, и короткие ветвящиеся дендрит ы. Звездчатые нейрон ы участвуют в процессах восприятия раздражении и объединении деятельности различных пирамидных нейрон ов.
Пирамидные нейрон ы осуществляют эфферентную функцию коры и внутрикорковые процессы взаимодействия между удаленными друг от друга нейрон ами. Они делятся на крупные пирамиды, от которых начинаются проекционные, или эфферентные, пути к подкорковым образованиям, и мелкие пирамиды, образующие ассоциативные пути к другим отделам коры. Наиболее крупные пирамидные клетки — гигантские пирамиды Беца — находятся в передней центральной извилине, в так называемой моторной зоне коры. Характерная особенность крупных пирамид — их вертикальная ориентация в толще коры. От тела клетки вертикально вверх к поверхности коры направлен наиболее толстый (верхушечный) дендрит , через который в клетку поступают различные афферентные влияния от других нейрон ов, а вертикально вниз отходит эфферентный отросток — аксон .
Многочисленность контактов (например, только на дендрит ах крупной пирамиды их насчитывают от 2 до 5 тыс.) обеспечивает возможность широкой регуляции деятельности пирамидных клеток со стороны множества других нейрон ов. Это позволяет координировать ответные реакции коры (в первую очередь ее моторную функцию) с разнообразными воздействиями из внешней среды и внутренней среды организма.
Для коры больших полушарий характерно обилие меж нейрон ных связей. По мере развития мозга человека после его рождения увеличивается число межцентральных взаимосвязей, особенно интенсивно до 18 лет.
.
Первичные, вторичные и третичные поля коры. Особенности строения и функционального значения отдельных участков коры позволяют выделить отдельные корковые поля.
Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
Первичные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в онтогенез е, имеют наиболее крупные клетки. Это так называемые ядерные зоны анализаторов, по И. П. Павлову (например, поле болевой, температурной, тактильной и мышечно-суставной чувствительности в задней центральной извилине коры, зрительное поле в затылочной области, слуховое поле в височной области и двигательное поле в передней центральной извилине коры) (рис. 54). Эти поля осуществляют анализ отдельных раздражений, поступающих в кору от соответствующих рецептор ов. При разрушении первичных полей возникают так называемая корковая слепота, корковая глухота и т. п. Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными органами только через первичные поля. Они служат для обобщения и дальнейшей обработки поступающей информации. Отдельные ощущения синтезируются в них в комплексы, обусловливающие процессы восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит их значения. Первичные и вторичные поля имеются и у человека, и у животных.
Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть только у человека. Они занимают почти половину территории коры и имеют обширные связи с другими отделами коры и с неспецифическими системами мозга. В этих полях преобладают наиболее мелкие и разнообразные клетки. Основным клеточным элементом здесь являются звездчатые нейрон ы. Третичные поля находятся в задней половине коры — на границах теменных, височных и затылочных ее областей и в передней половине — в передних частях лобных областей. В этих зонах оканчивается наибольшее число нервных волокон, соединяющих левое и правое полушария, поэтому роль их особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее сложные функции коры. Здесь происходят процессы высшего анализа и синтеза. В третичных полях на основе синтеза всех афферентных раздражении и с Учетом следов прежних раздражении вырабатываются цели и задачи поведения. Согласно им происходит программирование двигательной деятельности. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях.
При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит лишь бес смысл енные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т. п.).
Воспринимая и оценивая все сигналы из внутренней и внешней среды, кора больших полушарий осуществляет высшую регуляцию всех двигательных и эмоционально-вегетативных реакций.
Функции коры больших полушарии. Кора больших полушарий выполняет наиболее сложные функции организации приспособительного поведения организма во внешней среде. Это прежде всего функция высшего анализа и синтеза всех афферентных раздражении.
Афферентные сигналы поступают в кору по разным каналам, в разные ядерные зоны анализаторов (первичные поля), а затем синтезируются во вторичных и третичных полях, благодаря деятельности которых создается целостное восприятие внешнего мира. Этот синтез лежит в основе сложных психи ческих процессов восприятия, представления, мышлени я. Кора больших полушарий представляет собою орган, тесно связанный с возникновением у человека сознания и регуляцией его общественного поведения. Важной стороной деятельности коры больших полушарий является замыкательная функция — образование новых рефлексов и их систем (условные рефлексы, динамические стереотипы—см. главу XV).
Благодаря необычайно большой продолжительности сохранения в коре следов прежних раздражении (памяти) в ней накапливается огромный объем информации. Это имеет большое значение для сохранения индивидуального опыта, который используется по мере необходимости.
.
Экспериментально показано, что у высших представителей животного мира после полного оперативного удаления коры высшая нервная деятельность резко ухудшается. Они теряют способность тонко приспосабливаться к внешней среде и самостоятельно существовать в ней.
Источник: Физиология центральной нервной системы
Дата создания: 12.03.2008
Последнее редактирование: 17.03.2008
Кора представлена слоем серого вещества толщиной 3-5 мм. В коре насчитывают до 15 и более млрд. нейронов, а число глиоцитов в головном мозге — более 100 млрд.
Развитие. Формирование коры больших полушарий происходит путем закономерной миграции нейробластов эпендимного слоя вдоль вертикально ориентированных радиальных глиоцитов. Первыми возникают наиболее поверхностный и глубокий слои коры. Затем возникают очередные последовательные волны миграции групп нейробластов, которые дифференцируются в нейроны V-гo, затем IV-гo слоя и д. т. Таким образом, нейробласты очередной волны миграции преодолевают слой нейронов, возникших от более ранней волны миграции. Это создает послойную (экранную) цитоархитектонику коры большого мозга.
Между нейронами устанавливаются сложные взаимосвязи в соответствии с их местом в составе рефлекторных дуг. Формируются ядерные и экранные нервные центры. Тесные взаимоотношения в ходе гистогенеза складываются между нейронами и клетками глии.
Строение. Все нейроны коры мулътиполярные. Среди них по форме клеток различают пирамидные и непирамидные (звездчатые, корзинчатые, веретенообразные, паукообразные и горизонтальные) нейроны. Пирамидные нейроны, наиболее характерны для коры, имеют тело, по форме напоминающее пирамиду, вершина которой обращена к поверхности коры.
От основания пирамидной клетки отходит аксон, имеющий коллатерали (возвратные, горизонтальные, косые). От вершины и боковых поверхностей тела отходят длинные дендриты (апикальные и базальные). Апикальные дендриты группы нейронов объединяются в дендритические пучки. На поверхности дендритов одного пирамидного нейрона может быть до 4-6 тыс. специальных рецепторных аппаратов — шипиков. Наличие в последних актомиозинового комплекса позволяет изменять площадь синаптического контакта, следовательно, влиять на синаптическую связь.
Величина тела пирамидных клеток варьирует от 10 до 150 мкм. Различают малые, средние, крупные и гигантские пирамиды. Пирамидные клетки — это эфферентные нейроны коры, их аксонные коллатерали образуют 3/4 всех синапсов в коре.
Звездчатые нейроны имеют тело в форме звезды. Дендриты отходят во все стороны от тела звездчатого нейрона. Они в большинстве случаев короткие и лишены шипиков. Аксоны звездчатых клеток образуют сложные разветвления около клетки. Это так называемые околоклеточные паутинообразные аксонные сети. Эти клетки встречаются в нижних слоях коры.
Корзинчатые клетки (мелкие и крупные), расположены во П-м и Ш-м слоях коры, своими многочисленными отростками образуют синаптические связи с телами пирамидных нейронов V-ro слоя. Клетки содержат медиатор (ГАМК), который тормозит передачу возбуждения.
Нейроглиоморфные клетки встречаются во всех слоях коры. Это мелкие мультиполярные нейроны с короткими ветвящимися денд-ритами и аксонами.
Биполярные нейроны — малочисленная группа клеток, от тела которых отходят аксон и дендрит. В целом соотношение между пирамидными и другими формами нейронов равно 85:15, то есть в пользу пирамидных нейронов.
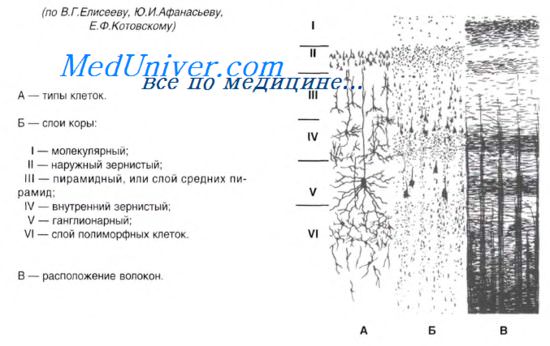
Цитоархитектоника. В двигательной зоне коры различают шесть основных слоев: молекулярный, наружный зернистый, пирамидный, внутренний зернистый, ганглионарный, слой полиморфных клеток.
В первом (наружном) молекулярном слое почти нет тел нейронов. Обнаруживаются единичные горизонтально ориентированные нейроны, тангенциальные ветвления нервных волокон нижележащих нейронов и глиальные клетки.
Второй, или наружный зернистый, слой содержит мелкие звездчатые и пирамидные нейроны размером около 10 мкм. Аксоны этих нейронов оканчиваются в III-м, IV-м и VI-м слоях коры, а дендриты поднимаются в молекулярный слой.
Третий слой — это слой средних и крупных пирамидных нейронов. Аксоны этих клеток образуют ассоциативные нервные волокна, идущие через белое вещество и соединяющие соседние участки коры.
Четвертый, или внутренний зернистый, слой содержит в основном мелкие звездчатые нейроны. Аксоны этих клеток, разветвляются в пределах соседних как выше, так и ниже лежащих слоев коры. Этот слой сильно развит в зрительной и слуховой зонах коры. Он состоит из сенсорных звездчатых нейронов, имеющих многочисленные ассоциативные связи с нейронами других типов.
Пятый — ганглионарный — слой образован большими пирамидными нейронами (клетками Беца). Апикальные дендриты нейронов направляются в молекулярный слой. Аксоны этих клеток идут в белое вещество, образуя комиссуральные и проекционные нервные волокна, и прежде всего пирамидные пути.
Шестой слой — слой полиморфных нейронов — содержит также много эфферентных пирамидных нейронов. Кроме того, здесь есть веретенообразные нейроны. Дендриты нейронов шестого слоя пронизывают всю толщину коры, достигая молекулярного слоя.
Читайте также:
