
Классификация нервных волокон скорость проведения возбуждения в них
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
Данная классификация была предложена Джозефом Эрлангером и Гербертом Гассером в 1937 г. Они проводили сверхпороговое электрическое раздражение смешанного нерва, содержащего волокна разного диаметра (как миелинизированные, так и немиелинизированные), так, чтобы все волокна вовлекались в возбуждение и генерировали ПД. В другой точке нерва, отстоящей от места раздражения на несколько сантиметров, с помощью униполярного внеклеточного электрода регистрировали ответ — суммарный ток действия нерва, который имел полифазную форму. На рис. 2.26 видно несколько пиков ПД, отражающих разное время прихода ПД к точке их регистрации — вследствие разной скорости распространения ПД по разным волокнам. Различия в скоростях были связаны с различиями в диаметре и степени миелинизации отдельных нервных волокон смешанного нерва. Таким образом, ПД возникают одновременно во всех волокнах в месте сверхпорогового раздражения нерва. Однако ПД распространяются далее по нерву с разной скоростью в зависимости от диаметра и миелинизации волокон. На основании проведенного анализа скоростей распространения ПД все нервные волокна были разбиты на три группы: группу А, включающую четыре подтипа (Аа, Ар, Ay, А8), группу В и группу С, различающиеся

Рис. 2.26. Схематическое изображение сложного потенциала действия, возникающего в результате возбуждения различных нервных волокон при отведении на большом расстоянии от места раздражения:
А, В, С — группы волокон; а, р, у — типы волокон группы А но степени миелинизации, диаметрам аксонов и, соответственно, скорости распространения ПД (табл. 2.1).
Типы нервных волокон, их свойства и функциональное назначение
Двигательные волокна соматической нервной системы
Чувствительные волокна кожных рецепторов
Чувствительные волокна нроириорецеи- торов
Чувствительные волокна терморецепторов, ноцицепторов
Иреганглионарные волокна вегетативной нервной системы
Постганглионарные волокна вегетативной нервной системы; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Волокна группы А покрыты миелиновой оболочкой. Наиболее толстые из них (типа а) имеют диаметр 12—22 мкм и скорость проведения возбуждения 70—120 м/с. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от рецепторов мышц к соответствующим нервным центрам. Три других типа волокон группы А (Р, у, 5) имеют меньший диаметр — от 1 до 8 мкм и меньшую скорость проведения возбуждения — от 5 до 70 м/с. Волокна этих типов преимущественно проводят возбуждение от различных рецепторов (тактильных, температурных, болевых, рецепторов внутренних органов) в ЦНС, за исключением у-волокон, значительная часть которых проводит возбуждение от спинного мозга к рецепторам растяжения мышц — интрафузальным мышечным волокнам.
К волокнам группы В относятся миелинизированные иреганглионарные волокна вегетативной нервной системы. Их диаметр — 1—3,5 мкм, а скорость проведения возбуждения — 3—18 м/с.
К волокнам группы С относятся безмиелиновые нервные волокна малого диаметра — 0,5—2 мкм. Скорость проведения возбуждения в этих волокнах — не более 3 м/с (0,5—3 м/с). Большинство волокон группы С — это постганглионарные волокна симпатического и парасимпатического отделов вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления. Таким образом, волокна с разной скоростью проведения выполняют различные физиологические функции.
А – нервные волокна с самой толстой миелиновой оболочкой. Наиболее высокая скорость передачи нервного импульса.
В – миелиновая оболочка тоньше, скорость проведения возбуждения ниже
С – безмиелиновые волокна с относительно низкой скоростью передачи импульса.
Диаметр волокна (мк)
Скорость проведения (м/сек)
Длительность потенциала действия (мсек)
Длительность отрицательного следового потенциала (мсек)
Длительность положительного следового потенциала (мсек)
Моторные волокна скелетных мышц, афферентные волокна мышечных рецепторов
Афферентные волокна от рецепторов прикосновения
Афферентные волокна от рецепторов прикосновения и давления, эфферентные волокна к мышечным веретенам
Афферентные волокна от некоторых рецепторов тепла, давления, боли
Преганглионарные вегетативные волокна
Преганглионарные вегетативные волокна, афеерентные волокна от некоторых рецепторов боли, давления, тепла
Типы нервных волокон, их свойства и функциональное назначение
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
7. Проведение возбуждения по немиелинизированным волокнам
В безмиелиновых волокнах нервный импульс распространяется волнообразно, последовательно возбуждаются небольшие участки размером в несколько микрометров. Возбуждение распространяется гораздо медленнее, чем по миелиновым волокнам.
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
| Тип | Диаметр (мкм) | Миелинизация | Скорость про-ведения (м/с) | Функциональное назначение |
| А alpha | 12–20 | сильная | 70–120 | Двигательные волокна соматической НС; чувствительные волокна проприорецепторов |
| А beta | 5–12 | сильная | 30–70 | Чувствительные волокна кожных рецепторов |
| А gamma | 3–16 | сильная | 15–30 | Чувствительные волокна проприорецепторов |
| А delta | 2–5 | сильная | 12–30 | Чувствительные волокна терморецепторов, ноцицепторов |
| В | 1–3 | слабая | 3–15 | Преганглионарные волокна симпатической НС |
| С | 0,3–1,3 | отсутствует | 0,5–2,3 | Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов |
Нервные волокна всех групп обладают общими свойствами:
• нервные волокна практически неутомляемы;
• нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Законы проведения возбуждения по нервным волокнам
• Закон анатомической и физиологической непрерывности –возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
• Закон двустороннего проведения возбуждения– возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
• Закон изолированного проведения– возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
Закономерности проведения местного и распространяющегося возбуждения
Электротонический потенциал (местное возбуждение)
• распространяется по нервным волокнам с затуханием (с декрементом), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
• вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
• местное возбуждение распространяется пассивно, без затрат энергии клетки;
• механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим.
Нервные волокна по скорости проведения возбуждения, диаметру волокна, длительности различных фаз потенциала действия и строению принято подразделять на три типа: А, В, С. Волокна типа А в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта. Волокна типа Апокрыты миелиновой оболочкой. Наиболее толстые из них А - альфа, они имеют диаметр 12-22 мкм и высокую скорость проведения возбуждения - 70-120 м/с. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам. Три другие группы волокон тина А (бета, гамма, дельта) имеют меньший диаметр от 8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна этих групп относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых, рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам. К волокнам типа В относятся миелинизированные преганглио-нарные волокна вегетативной нервной системы. Их диаметр - 1-3,5 мкм, а скорость проведения возбуждения - 3-18 м/с. К волокнам типа Сотносятся безмиелиновые нервные волокна малого диаметра - 0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более 3 м/с (0,5-3,0 м/с) . Большинство волокон типа С - это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
Механизм проведения возбуждения по нервным волокнам.
Механизм распространения возбуждения у различных нервных волокон неодинаков. По современным представлениям распространение возбуждения по нервным волокнам осуществляется на основе ионных механизмов генерации потенциала действия. При распространении возбуждения по безмиелиновому нервному волокну местные электрические токи, которые возникают между его возбужденным участком, заряженным отрицательно, и невозбужденным, заряженным положительно, вызывают деполяризацию мембраны до критического уровня с последующей генерацией ПД в ближайшей точке невозбужденного участка мембраны. Этот процесс повторяется многократно. На всем протяжении нервного волокна происходит процесс репродукции нового ПД в каждой точке мембраны волокна. Такое проведение возбуждения называете и непрерывным. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки (перехватов Ранвье) создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Местные электрические токи возникают между соседними перехватами Ранвье, т. к. мембрана возбужденного перехвата становится заряженной отрицательно по отношению к поверхности соседнего невозбужденного перехвата. Эти местные токи деполярязуют мембрану невозбужденного перехвата до критического уровня и в нем возникает ПД (рис. 4). Следовательно, возбуждение как бы "перепрыгивает" через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Такой механизм распространения возбуждения называется сальтаторным или скачкообразным. Скорость такого способа проведения возбуждения значительно выше и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов.
Функциональная лабильность (Н.Е.Введенский).
Нервные волокна обладают лабильностью - способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в единицу времени без трансформации ритма раздражения. Лабильность определяется длительностью пика потенциала действия, т. е. фазой абсолютной рефрактерности. Так как длительность абсолютной рефрактерности у спайкового потенциала нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000 импульсов в секунду. Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется.
Дата добавления: 2018-08-06 ; просмотров: 367 ;
Классификация нервных волокон
Амплитуда электрических импульсов, отводимых от целого нервного ствола, зависит от силы приложенного раздражения. Раздражению слабой силы соответствует небольшой потенциал действия, по мере же усиления раздражения амплитуда пика возрастает, достигает максимальной величины и затем остается постоянной, несмотря на дальнейшее увеличение силы раздражителя ( рис. 156 ).
Рис. 156. Ответы седалищного нерва лягушки на электрические стимулы возрастающей силы (по Я. Бурешу). Отведение потенциалов действия двухфазное. В начале каждой записи артефакт раздражения. Сила тока увеличивается постепенно от 1 к 6. Отметка времени 1 мсек.
При слабой силе стимула возбуждение возникает в наиболее возбудимых поверхностно расположенных нервных волокнах. Усиление стимула приводит к увеличению числа возбужденных волокон, поэтому суммарный пик увеличивается до тех пор, пока все волокна не вовлекаются в реакцию.
Такую картину можно наблюдать, если отводящие электроды расположить на нерве вблизи раздражающих электродов. При увеличении расстояния между этими двумя парами электродов суммарный потенциал действия начинает расчленяться на несколько отдельных колебаний, которые становятся наиболее отчетливо выраженными при удалении отводящих электродов на 10—15 см от места раздражения ( рис. 157 ).
Впервые это явление было подробно изучено Эрлангером и Гассером (1937). Они показали, что причиной расчленения суммарного потенциала действия на компоненты является неодинаковая скорость проведения возбуждения по разным волокнам, вследствие чего к отводящим электродам нервные импульсы поступают по этим волокнам неодновременно.
Рис. 157. Комплекс составного потенциала действия смешанного нервного ствола лягушки (по Эрлангеру и Гассеру).
Детальный анализ показал, что между скоростью проведения импульса и диаметром нервного волокна существует примерно пропорциональная зависимость: нервные волокна проводят тем быстрее, чем они толще.
В настоящее время принято подразделять нервные волокна по скорости проведения возбуждения, длительности различных фаз потенциала действия и строению на три основных типа, обозначаемых буквами А, В и С ( рис. 158 ).
Рис. 158. Схематическое изображение всех составных частей потенциала действия смешанного нерва (по Эрлангеру и Гассеру).
Классификация нервных волокон
Три другие группы волокон типа А: бета (Aβ)-, гамма (Аγ)- и дельта волокна имеют меньший диаметр — от 2 до 12 мк, меньшую скорость проведения и более длительный потенциал действия. Это преимущественно чувствительные волокна, проводящие возбуждение от различных рецепторов (тактильных, некоторых болевых, температурных и рецепторов внутренних органов) в центральную нервную систему. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение в центробежном направлении — от клеток спинного мозга к так называемым интрафузальным мышечным волокнам которых расположены чувствительные нервные окончания — мышечные веретена.
К волокнам типа В относятся мпелиновые, преимущественно преганглионарные волокна автономной нервной системы. Скорость проведения возбуждения в этих волокнах у теплокровных животных составляет 3—14 м/сек. Продолжительность потенциала действия волокон типа В примерно в 2 раза превышает длительность потенциала действия волокон типа А. В среднем она составляет 1,2 мсек. Отличительной особенностью этих волокон является то, что в них не обнаруживается следового отрицательного потенциала: фаза реполяризации непосредственно переходит в следовой положительный потенциал, имеющий большую длительность — 100—300 мсек.
К волокнам типа С относят безмякотиые нервные волокна очень малого диаметра (порядка 0,5 мк). Скорость проведения возбуждения в этих волокнах 2—6 м/сек. Большинство С-волокон относится к постганглионарным волокнам симпатической нервной системы.
К С-волокнам относят также те нервные волокна, которые участвуют в проведении возбуждения от болевых рецепторов и некоторых рецепторов холода, тепла и давления.
Потенциалы действия С-волокон характеризуются наибольшей продолжительностью (2 мсек у теплокровных животных). Они имеют длительный следовой отрицательный потенциал (50—80 мсек), сопровождающийся еще более продолжительной следовой позитивностью (300—1000 мсек).
Все эти данные суммированы в таблице .
Свойства различных нервных волокон теплокровных
Длительность
потенциала
действия
(мсек)
Законы проведения возбуждения по нервным волокнам
• Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
• Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
• Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
По толщине и по скорости проведения нервного импульса волокна делятся на три основных типа: A, B и С.
Волокна типа А самые толстые волокна в нс.
Все волокна типа А миелинизированные. Их диаметр варьируется от 3 до 22 мкм (микрометров). Скорость проведения нервного импульса – от 5 до 120 м/с.Чем толще волокно, тем быстрее проведение по нему нервного импульса. Волокна типа А делятся на четыре вида:
- Аα. Диаметр от 13 до 22 мкм. Скорость проведения нервного импульса от 70 до 120 м/с. Это самые толстые и быстрые волокна в нервной системы. Эти волокна иннервируют скелетные мышцы. Это эфферентные волокна передних рогов спинного мозга.
- Аβ. Диаметр от 8 до 13 мкм. Скорость проведения нервного импульса от 40 до 70 м/с. Они относятся к афферентным волокнам рецепторов давления и прикосновения.
- Аγ. Диаметр от 4 до 8 мкм. Скорость проведения нервного импульса от 15 до 40 м/с. Эфферентные волокна мышечных веретен и часть афферентных волокон идущих от давления и прикосновения. Также, это волокна 1-го афферентного нейрона (? От себя).
- Аδ. Диаметр от 3 до 4 мкм. Скорость проведения нервного импульса от 5 до 15 м/с. Афферентные волокна от кожных температурных и кожных болевых рецепторов. Также, частично, рецепторы давления.
Волокна типа В частично миелинизированные. Они более тонкие, чем волокна типа А. Диаметр этих волокон от 1 до 3 мкм. Скорость проведения нервного импульса от 3 до 18 м/с. Это волокна вегетативной нервной системы (боковые рога спинного мозга) преганглионарные волокна.
Волокна типа Снемиелизированные. Диметр от 0.5 до 2 мкм. Скорость проведения нервного импульса от 1.5 до 3 м/с. Волокна вегетативной НС постганглионарные волокна и в соматической НС болевые и температурные.
Волокна типа А более лабильны, т.е. наиболее высокая частота передачи нервного импульса. У них очень короткое время рефрактерного периода. ПД в них протекает очень быстро, примерно 0.5 мс (миллисекунды).
Нейроглия, ее виды и значение.
Нейроглия - обширная гетерогенная группа элементов нервной ткани, обеспечивающая деятельность нейронов и выполняющая неспецифические функции: опорную, трофическую, разграничительную, барьерную, секреторную и защитную функции. Является вспомагательнымкомпанентом нервной ткани.
В мозге человека содержание глиальных клеток (глиоцитов) в 5-10 раз превышает число нейронов, причем они занимают около половины его объема. В отличие от нейронов, глиоциты взрослого способны к делению. В поврежденных участках мозга они размножаются, заполняя дефекты и образуя глиальные рубцы (глиоз); опухоли из клеток глии (глиомы) составляют 50% внутричерепных новообразований.
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
Артериальное давление — это давление крови в крупных артериях человека. Различают два показателя артериального давления:
Систолическое (верхнее) артериальное давление — это уровень давления крови в момент максимального сокращения сердца.
Диастолическое (нижнее) артериальное давление — это уровень давления крови в момент максимального расслабления сердца.
Артериальное давление измеряется в миллиметрах ртутного столба, сокращенно мм рт. ст. Значение величины артериального давления 120/80 означает, что величина систолического (верхнего) давления равна 120 мм рт. ст., а величина диастолического (нижнего) артериального давления равна 80 мм рт. ст.
Давление в правом предсердии регулируется соотношением двух факторов: (1) способности сердца перекачивать кровь из правого предсердия и правого желудочка в малый круг кровообращения; (2) поступления крови из периферических вен в правое предсердие. Если сокращения правых отделов сердца усиливаются, давление в правом предсердии уменьшается. С другой стороны, любые факторы, которые ускоряют приток крови в правое предсердие из периферических вен, вызывают повышение давления в правом предсердии. Такими факторами могут быть: (1) увеличение общего объема крови; (2) увеличение тонуса крупных вен всех сосудистых областей организма и повышение периферического венозного давления; (3) расширение артериол, которое приводит к понижению периферического сопротивления и обеспечивает быстрый отток крови из артерий в вены. Все факторы, которые регулируют давление в правом предсердии, регулируют также и сердечный выброс, т.к. количество крови, которое перекачивает сердце, зависит как от сократительной функции сердца, так и от поступления крови к сердцу из периферических вен, поэтому мы подробно обсуждаем регуляцию давления в правом предсердии, которая посвящена регуляции сердечного выброса. В норме давление в правом предсердии равно 0 мм рт. ст. (т.е. равно атмосферному давлению). В патологических случаях оно может увеличиваться до 20-30 мм рт. ст., например: (1) при сердечной недостаточности; (2) в результате массивного переливания крови, которое приводит к увеличению общего объема крови в организме и к избыточному поступлению крови к сердцу из периферических вен. Нижний предел давления в правом предсердии — примерно от -3 до -5 мм рт. ст., что ниже уровня атмосферного давления. Таким же отрицательным является давление в грудной полости вокруг сердца. Давление в правом предсердии достигает такого низкого уровня, когда сердце усиленно перекачивает кровь, или когда приток крови к сердцу уменьшен в результате большой кровопотери.
85. Спинальный шок
Когда спинной мозг внезапно пересекается в верхней части шеи, сначала практически все функции спинного мозга, включая спинномозговые рефлексы, мгновенно подавляются, вплоть до полного их выключения. Эту реакцию называют спинальным шоком. Причиной такой реакции является то, что нормальная активность спинальных нейронов зависит в большой степени от постоянного тонического возбуждения спинного мозга под действием импульсов, поступающих к нему по нисходящим нервным волокнам от высших центров, особенно по ретикулоспинальным, вестибулоспинальным и кортикоспинальным трактам. В течение нескольких часов или недель возбудимость спинальных нейронов постепенно восстанавливается. По-видимому, это общее естественное свойство нейронов повсюду в нервной системе, т.е. после потери источника облегчающих импульсов нейроны увеличивают собственную естественную степень возбудимости, чтобы компенсировать потерю хотя бы частично. Большинству неприматов, чтобы возвратить к норме возбудимость центров спинного мозга, требуется от нескольких часов до нескольких суток. Однако у человека восстановление часто задерживается на несколько недель, а иногда полного восстановления совсем не происходит. В других случаях, наоборот, происходит чрезмерное восстановление с результирующей повышенной возбудимостью некоторых или всех функций спинного мозга. Далее перечислены некоторые из спинальных функций, особенно страдающих во время или после спинального шока. 1. В начале спинального шока сразу и очень значительно падает артериальное давление, иногда опускаясь ниже 40 мм рт. ст., что свидетельствует о практически полной блокаде активности симпатической нервной системы. Давление обычно возвращается к норме в течение нескольких дней (даже у человека). 2. Все рефлексы скелетных мышц, интегрированные в спинном мозге, во время первых стадий шока блокируются. Животным для восстановления этих рефлексов до нормы требуется от нескольких часов до нескольких дней; людям — от 2 нед до нескольких месяцев. И у животных, и у человека некоторые рефлексы могут в итоге стать чрезмерно возбудимыми, особенно в тех случаях, когда на фоне пересечения основной части путей между головным и спинным мозгом некоторые облегчающие пути сохраняются. Первыми восстанавливаются рефлексы на растяжение, в дальнейшем постепенно восстанавливаются более сложные рефлексы в соответствующем порядке: сгибательные, антигравитационные позные и частично шагательные. 3. Рефлексы крестцового отдела спинного мозга, контролирующие опорожнение мочевого пузыря и прямой кишки, подавлены у человека в течение первых недель после пересечения спинного мозга, но в большинстве случаев они в итоге восстанавливаются.
Синдром Броун-Секара - синдром половинного поперечного поражения спинного мозга, проявляющийся параличом , утратой суставно-мышечной чувствительности и утратой вибрационной чувствительности на противоположной стороне. Иногда гемипарез сочетается с ипсилатеральной потерей проприоцептивной чувствительности и контралатеральным выпадением болевой чувствительности и контралатеральным выпадением температурной чувствительности .
Синдром Броун-Секара возникает при повреждении половины поперечника спинного мозга и характеризуется парезом (повреждение кортикоспинального пути ) и утратой проприоцептивной чувствительности (повреждение задних канатиков ) на стороне поражения, а утратой болевой и температурной чувствительности ( спиноталамический путь ) - на противоположной стороне (на 1-2 сегмента ниже уровня поражения). Все нарушения развиваются ниже уровня поражения. Сегментарные расстройства ( корешковая боль , атрофия мышц или утрата сухожильных рефлексов ) выявляются не всегда и бывают односторонними.
Классический синдром Броун-Секара встречается редко. Чаще наблюдается частичный синдром или полное поперечное повреждение спинного мозга. При повреждении одного из задних квадрантов спинного мозга утрачивается вибрационная и проприоцептивная чувствительность на стороне поражения, одного из передних квадрантов - возникает ипсилатеральный парез и контралатеральное нарушение болевой и температурной чувствительности .
23) Строение, классификация и функциональные свойства синапсов. Морфо-функциональные особенности электрических и химических синапсов.
Синапс —специализированная зона контакта между нейронами или нейронами и другими возбудимыми образованиями, обеспечивающая передачу сигналов с сохранением, изменением или исчезновением ее информационного значения.
Если иннервируемая клетка нервная, то синапс называется межнейронным, кроме межнейронных синапсов по месту расположения выделяют :
1. нейроэффекторные (иннервируемая клетка мышечная или железистая)
2. нейрорецепторные (контакт между нейроном и вторично-чувствующей рецепторной клеткой).
• Межнейронные синапсы делятся на:
3. аксо-аксональные в зависимости от того, с какой частью иннервируемой клетки контактирует аксон –соответственно с телом клетки, дендритом, аксоном.
Есть также дендро-дендритические, дендросоматические и сомато-соматические синапсы, но они встречаются редко и функция их не выяснена.
Нейроэффекторные синапсы в зависимости от органа-эффектора бывают нейромышечные, нейрососудистые, нейросекреторные.
• По действию на иннервируемую клетку синапсы делят на :
• По механизму передачи существуют
Структура и механизм действие синапса
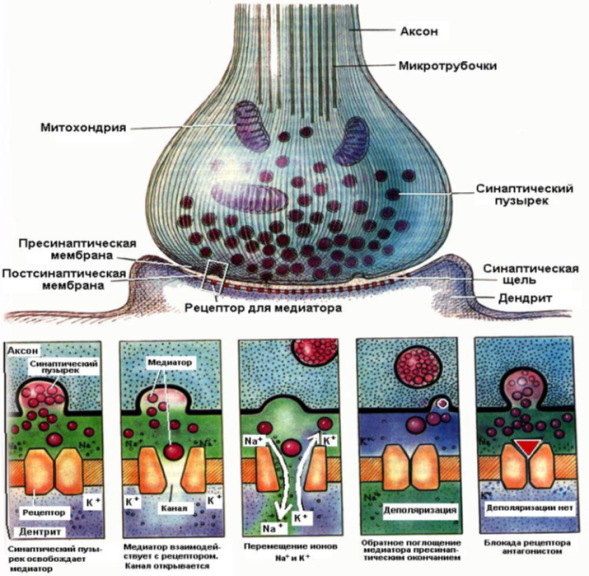
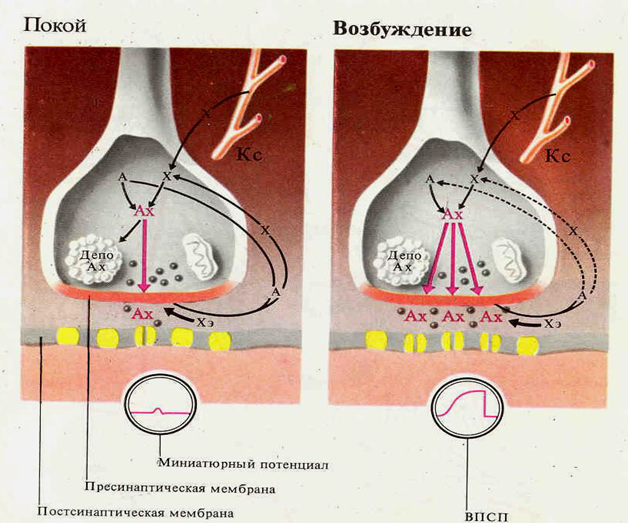
Синаптические процессы в возбуждённом и невозбуждённом синапсе
Контактное взаимодействие обусловлено специфическими контактами мембран нейронов, которые образуют так называемые электрические и химические синапсы.
Электрические синапсы. Морфологически представляют собой слияние, или сближение, участков мембран. В последнем случае синаптическая щель не сплошная, а прерывается мостиками полного контакта. Эти мостики образуют повторяющуюся ячеистую структуру синапса, причем ячейки ограничены участками сближенных мембран, расстояние между которыми в синапсах млекопитающих 0,15—0,20 нм. В участках слияния мембран находятся каналы, через которые клетки могут обмениваться некоторыми продуктами. Кроме описанных ячеистых синапсов, среди электрических синапсов различают другие — в форме сплошной щели; площадь каждого из них достигает 1000 мкм, как, например, между нейронами ресничного ганглия.
Электрические синапсы обладают односторонним проведением возбуждения. Это легко доказать при регистрировании электрического потенциала на синапсе: при раздражении афферентных путей мембрана синапса деполяризуется, а при раздражении эфферентных волокон — гиперполяризуется. Оказалось, что синапсы нейронов с одинаковой функцией обладают двусторонним проведением возбуждения (например, синапсы между двумя чувствительными клетками), а синапсы между разнофункциональными нейронами (сенсорные и моторные) обладают односторонним проведением. Функции электрических синапсов заключаются прежде всего в обеспечении срочных реакций организма. Этим, видимо, объясняется расположение их у животных в структурах, обеспечивающих реакцию бегства, спасения от опасности и т. д. Электрический синапс сравнительно мало утомляем, устойчив к изменениям внешней и внутренней среды. Видимо, эти качества наряду с быстродействием обеспечивают высокую надежность его работы.
Химические синапсы. Структурно представлены пресинаптической частью, синаптической щелью и постсинаптической частью. Пресинаптическая часть химического синапса образуется расширением аксона по его ходу или окончания (рис. 2.19). В пресинаптической части имеются агранулярные и гранулярные пузырьки. Пузырьки (кванты) содержат медиатор. В пресинаптическом расширении находятся митохондрии, обеспечивающие синтез медиатора, гранулы гликогена и др. При многократном раздражении пресинаптического окончания запасы медиатора в синаптических пузырьках истощаются. Считают, что мелкие гранулярные пузырьки содержат норадреналин, крупные — другие катехоламины. Агранулярные пузырьки содержат ацетилхолин. Медиаторами возбуждения могут быть также производные глутаминовой и аспарагиновой кислот. Для синапсов с химическим способом передачи возбуждения характерны синоптическая задержка проведения возбуждения, длящаяся около 0,5 мс, и развитие постсинаптического потенциала (ПСП) в ответ на пресинаптический импульс. Этот потенциал при возбуждении проявляется в деполяризации постсинаптической мембраны, а при торможении — в гиперполяризации ее, в результате чего развивается тормозной постсинаптический потенциал (ТПСП). При возбуждении проводимость постсинаптической мембраны увеличивается.
ВПСП возникает в нейронах при действии в синапсах ацетил холина, норадреналина, дофамина, серотонина, глутаминовой кислоты, вещества Р.
ТПСП возникает при действии в синапсах глицина, гамма-аминомасляной кислоты. ТПСП может развиваться и под действием медиаторов, вызывающих ВПСП, но в этих случаях медиатор вызывает пе-реход постсинаптической мембраны в состояние гиперполяризации. Для распространения возбуждения через химический синапс важно, что нервный импульс, идущий по пресинаптической части, полностью гасится в синаптической щели. Однако нервный импульс вызывает физиологические изменения в пресинаптической части мембраны. В результате у ее поверхности скапливаются синаптические пузырьки, изливающие медиатор в синаптическую щель.
Последнее изменение этой страницы: 2016-08-01; Нарушение авторского права страницы
Читайте также:
