
Возбудимость мембраны в нервном волокне выше
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
Вопросы для заключительного контроля уровня знаний.
1. Какие электроды используют для регистрации мембранного потенциала нервной клетки?
2. Какими растворами заполняют стеклянные микроэлектроды?
3. Какие ионы принимают участие в поддержании потенциала покоя?
4. В чем различия между распространяющимся и местным возбуждением?
5. Что называют латентным периодом?
6. Как изменится потенциал действия скелетной мышцы при нанесении на нее ритмического раздражения?
7. Объясните возникновение потенциала действия с точки зрения мембранно-ионной теории.
8. Зарисуйте основные структурные элементы плазматической мембраны.
9. Какие анионы не проникают через плазматическую мембрану?
10. Как изменяется проницаемость ионов Na при развитии потенциала действия?
Приложение 5.
1. Микроэлектродным методом измеряют потенциал покоя нервной клетки. Что показывает прибор, если микроэлектрод: а) находится на наружной поверхности мембраны; б) проколол мембрану; в) введен в глубь клетки?
Ответ: а) прибор показывает ноль, поскольку оба электрода (микро- и макро-) находятся снаружи, в области положительного потенциала; б) величину, равную потенциалу покоя; в) величину мембранного потенциала, потому что величина потенциала внутри клетки одинакова в любом участке.
2. Если бы клеточная мембрана была абсолютно непроницаемая для ионов, как бы изменилась величина потенциала покоя?
Ответ: Если бы мембрана была непроницаемой для ионов, в том числе и для ионов калия, то потенциал покоя не мог возникнуть (равнялся бы нулю).
3. Как изменится кривая потенциала действия при замедлении процесса инактивации натриевых каналов?
Ответ: инактивация натриевых каналов полностью прекращает процесс деполяризации мембраны и он сменяется реполяризацией, что приводит к восстановлению исходного уровня мембранного потенциала.
4. Возбудимость нервных волокон выше, чем мышечных. В чем причина этого?
Ответ: для того, чтобы возник потенциал действия, мембранный потенциал должен уменьшиться до критического уровня деполяризации (КУД).В нервном волокне мембранный потенциал – 70 мВ, а в мышечном – 90 мВ. Соответственно, величина порогового потенциала для нервного волокна 20 мВ, а для мышечного – 40. Поэтому нервные волокна обладают более высокой возбудимостью.
5. Почему гиперполяризация мембраны приводит к снижению возбудимости?
Ответ: гиперполяризация – увеличение мембранного потенциала. Но если мембранный потенциал увеличивается, то возрастает величина порогового раздражения. Значит возбудимость снизится.
6. Нерв раздражают с частотой 10, 100 раз в секунду. Сколько потенциал действия будет возникать в каждом случае?
Ответ: при частоте 10 Гц интервал между раздражителями составляет 0,1 с. Нерв все 10 раз ответит возникновением потенциала действия. При частоте 100 Гц интервал между раздражителями (0,01 с) тоже достаточно велик и мы получим 100 потенциала действия.
7. При положительном следовом потенциале наблюдается:
Ответ: понижение возбудимости ткани; гиперполяризация мембраны.
8. Фазу быстрой реполяризации обеспечивает:
Ответ: инактивация натриевой проницаемости мембраны; открытие потенциалозависимых калиевых каналов.
9. В потенциале действия выделяют фазы:
Ответ: начальной деполяризации; быстрой деполяризации; быстрой реполяризации; следовых потенциалов.
10. Повышение возбудимости ткани соответствует фазам потенциала действия:
Ответ: начальной деполяризации; отрицательному следовому потенциалу.
1. Потенциал действия и его происхождение.
2. Критический уровень деполяризации.
3. Фаза потенциала действия и их ионные механизмы.
4. Методы регистрации потенциала действия.
5. Уравнения Нернста и Гольдмана.
6. Изменение возбудимости ткани при возбуждении.
7. Локальный ответ, отличительные особенности местного и распространяющегося возбуждения.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Физиологические свойства нервных волокон. Мякотные и безмякотные нервные волокна
Нерв, или нервный ствол, представляет собой сложное образование, состоящее из большого количества нервных волокон, заключенных в общую соединительнотканную оболочку. Нервные волокна - это отростки нервных клеток. В состав нерва входят длинные отростки нервных клеток (аксоны), которые несут возбуждение в центральную нервную систему или от нее на периферию. Нервы, как правило, являются смешанными, в их состав входят двигательные и чувствительные нервные волокна.
Нервные волокна, как и все возбудимые структуры, обладают следующими физиологическими свойствами: возбудимостью, проводимостью, рефрактерностью, лабильностью.
При сравнении основных физиологических свойств нервной и мышечной ткани обращает на себя внимание то обстоятельство, что возбудимость и лабильность нервного волокна выше, а рефрактерный период короче, чем у мышечной ткани. Данная особенность связана с более высоким уровнем обменных процессов в нерве. Например, абсолютный рефрактерный период мякотного нерва продолжается 0,002 с, а скелетной мышцы - 0,005 с. Лабильность двигательного нерва нервно-мышечного препарата лягушки составляет 500-1000 имп/с, лабильность же мышцы - 200-250 имп/с.
Проведение возбуждения является специализированной функцией нервных волокон. Скорость проведения возбуждения по нервным волокнам в основном зависит от диаметра и гистологических особенностей строения нервных волокон. Чем больше диаметр нервного волокна, тем скорость распространения возбуждения в нем выше. Так, скорость распространения возбуждения по нервному волокну с диаметром 12-22 мкм составляет 70-120 м/с, а по нервному волокну с диаметром 8-12 мкм - 40-70 м/с.
В зависимости от гистологических особенностей строения нервные волокна делятся на мякотные (миелиновые) и безмякотные (безмиелиновые).
Миелинизированное волокно (рис. 57) состоит из осевого цилиндра и покрывающих его миелиновой и шванновской оболочек. Миелиновая оболочка состоит из жироподобных веществ, обладает высоким удельным сопротивлением и выполняет в связи с этим роль своеобразного изолятора. Миелиновая оболочка через промежутки равной длины прерывается, оставляя открытыми участки осевого цилиндра шириной около 1 мкм. Эти участки получили название перехватов Ранвье. Длина межперехватных участков зависит от диаметра волокна и колеблется в пределах от 0,2·10 -3 м (0,2 мм) до 1·10 -3 -2·10 -3 м (1-2 мм). Поверхность осевого цилиндра представлена плазматической мембраной, а его содержимое - аксоплазмой.
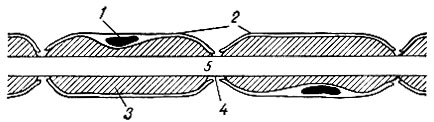
Рис. 57. Строение миелинизированного нервного волокна (схема). 1 - ядро шванновской плетки; 2 - шванновская оболочка; 3 - миелин; 4 - перехват Ранвье; 5 - аксоплазма
Безмякотные нервные волокна не имеют миелиновой оболочки, они покрыты только шванновскими клетками (леммоциты). Между шванновскими клетками и осевым цилиндром имеется щель 15 нм (150 А), которая заполнена межклеточной жидкостью. В связи с указанной особенностью строения поверхностная мембрана осевого цилиндра сообщается с окружающей нервное волокно средой (межклеточная жидкость).
Основоположником учения о проведении возбуждения по нервным волокнам следует считать немецкого физиолога Германа (1885). Он полагал, что возбуждение по нервным волокнам распространяется за счет малых круговых токов, которые возникают внутри волокна и в окружающей его жидкости (рис. 58).
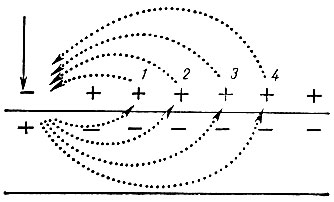
Рис. 58. Схема распространения возбуждения по нервному волокну за счет малых круговых токов. Вертикальной стрелкой обозначено место нанесения раздражения. Круговые стрелки показывают направление движения электрического тока внутри волокна и в окружающей жидкости
В настоящее время теория круговых токов Германа получила теоретическую разработку и экспериментальное подтверждение в работах Ходжкина и других исследователей.
Между возбужденным и невозбужденным участками нервного волокна в аксоплазме и в окружающей жидкости ток будет распространяться от плюса к минусу. Это приведет к возникновению так называемых малых, или круговых, токов, которые, выходя из нервного волокна, последовательно возбуждают его участки (1, 2 и т. д.). По мере удаления от очага возбуждения (участки 3 и 4) раздражающее действие круговых токов ослабевает и они становятся неспособными вызвать возбуждение. Таким образом, в случае последовательного распространения возбуждения по каждому участку нервного волокна нервные импульсы передаются с затуханием за счет непрерывно бегущей волны.
Из-за гистологических особенностей строения мякотных нервных волокон, в частности наличия миелиновой оболочки, обладающей высоким сопротивлением, электрические токи могут входить в волокна указанного типа и выходить из них только в области перехватов Ранвье (рис. 59).

Рис. 59. Скачкообразное распространение возбуждения в мякотном нервном волокне от перехвата к перехвату. Стрелками показано направление тока, возникающего между возбужденным (А) и соседним покоящимся (Б) перехватом
При нанесении раздражения возникает деполяризация в области ближайшего перехвата Ранвье - А. Соседний перехват Ранвье - Б находится в состоянии поляризации. Между перехватами возникает разность потенциалов, которая приводит к появлению круговых токов (см. рис. 58). Ионный поток в аксоплазме и в окружающей среде течет от плюса к минусу. Выход круговых токов в перехвате Б приводит к его деполяризации и возникновению потенциала действия. Далее за счет круговых токов возбуждаются последующие перехваты Ранвье. Таким образом, возбуждение в мякотных нервных волокнах передается скачкообразно (сальтаторно) от одного перехвата Ранвье к другому, Сальтаторный способ передачи возбуждения более экономичен, нежели распространение возбуждения по безмякотным нервным волокнам. Возбуждение по мякотным нервным волокнам распространяется без затухания. Скорость распространения возбуждения по мякотным нервным волокнам гораздо выше, чем по безмякотным. Так, скорость распространения возбуждения по двигательным нервным волокнам (мякотные нервы) составляет 80-120 м/с, по волокнам, не покрытым миелиновой оболочкой, - от 0,5 до 2 м/с.
При нанесении раздражения на нервное волокно происходит двустороннее распространение возбуждения - в центростремительном и центробежном направлениях. Это доказывается следующим опытом. К нервному волокну прикладывают две пары электродов - А и Б, связанных с электроизмерительными приборами (рис. 60). Раздражение наносят между электродами А и Б с помощью раздражающих электродов С. В результате двустороннего проведения возбуждения вдоль клеточной мембраны приборы зарегистрируют прохождение нервных импульсов как под электродом А, так и под электродом Б.
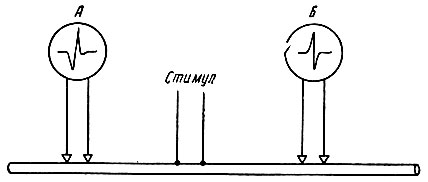
Рис. 60. Схема опыта для доказательства двустороннего проведения возбуждения по нерву. Объяснение в тексте
Двустороннее проведение возбуждения по нервному волокну впервые описано русскими учеными Р. И. Бабухиным (1887) и Кюне (1886). Оно не противоречит одностороннему распространению нервных импульсов в целостном организме, которое объясняется местом возникновения нервных импульсов (рецептор или нервный центр), а также наличием специальных образований - синапсов.
Возбуждение проводится только по одному нервному волокну, не распространяясь на соседние волокна, что обусловливает осуществление строго координированной рефлекторной деятельности. Периферический нервный ствол обычно состоит из большого количества нервных волокон. Так, в состав седалищного нерва входят тысячи нервных волокон: мякотные и безмякотные, афферентные и эфферентные, соматические и вегетативные. В случае неизолированного проведения возбуждения наблюдалась бы хаотическая ответная реакция.
Изолированное проведение возбуждения в мякотных нервных волокнах обеспечивается миелиновой оболочкой, в безмякотных - высоким удельным сопротивлением окружающей нервное волокно жидкости. Доказать наличие изолированного проведения возбуждения можно в эксперименте на препарате задней лапки лягушки, раздражая отдельные корешки седалищного нерва. Более строгое доказательство может быть получено при отведении потенциала действия от отдельных нервных волокон, входящих в состав нервного ствола.
Химические изменения в нервных волокнах при возбуждении. Неспецифическим признаком возбуждения, как уже отмечалось, является повышение обменных процессов. Возбуждение нерва сопровождается увеличением распада богатых энергией фосфорных соединений АТФ (аденозинтрифосфорная кислота) и креатинфосфата, а также повышенным образованием молочной кислоты. Кроме того, наблюдается усиление процессов распада и синтеза углеводов, белков и липидов.
Стимуляция обменных процессов приводит к увеличению потребления кислорода и выделению соответствующих количеств углекислого газа. Так, нерв лягушки в состоянии покоя потребляет 1·10 -3 м 3 (1 мм 3 ) кислорода на 1·10 -3 кг/мин (1 г/мин), при возбуждении потребление кислорода возрастает на 10-20%, то же происходит и с продукцией углекислого газа.
Обмену веществ и его усилению при возбуждении соответствуют и термические соотношения. По данным Хилла, 1·10 -3 кг (1 г) вещества нерва в состоянии покоя образует в 1 с 2·10 -5 кал, а при возбуждении - 6,9·10 -5 кал. Таким образом, по теплопродукции нерва можно судить о динамике обмена веществ в нем.
Утомление нервного волокна. Н. Е. Введенский в 1883 г. впервые установил, что нерв мало утомляем. Он проводил опыт на нервно-мышечном препарате лягушки, раздражая седалищный нерв индукционным током в течение 6-8 ч. О возбуждении нерва Н. Е. Введенский судил по сокращению мышцы. Для того чтобы исключить утомление мышцы, распространение нервных импульсов к ней прерывалось действием катода постоянного тока. При выключении постоянного тока возбуждение вновь достигало мышцы и она сокращалась. В этих условиях установлено, что нерв длительно (на протяжении многих часов) сохранял способность возбуждаться и проводить волны возбуждения. Данные экспериментов Н. Е. Введенского подтверждены современными исследованиями с регистрацией биотоков нерва.
Малая утомляемость нервных волокон объясняется тем, что энергетические затраты в них при возбуждении незначительны, а восстановительные процессы протекают быстро.
В целостном организме малой утомляемости нервных волокон способствует также их работа с постоянной недогрузкой. Так, двигательное нервное волокно обладает высокой лабильностью (может воспроизводить до 2500 имп/с, из нервных же центров на периферию обычно проводится прерывисто не более 50-100 волн возбуждения в 1 с).
Таким образом, практическая неутомляемость нервных волокон связана с низкими энергетическими затратами при возбуждении, с высокой лабильностью нервных волокон и постоянной работой их с недогрузкой.
Синапс - функциональный контакт между возбудимыми клетками, цитоплазма каждой из которых заключена в отдельную электрогенную мембрану.
В зависимости от локализации синапсы делят на центральные и периферические.
Центральные синапсы осуществляют контакт между нервными клетками центральной нервной системы.
Периферические синапсы могут быть мионевральными и невроэпителиальными. За счет невроэпителиальных синапсов осуществляется нервная регуляция деятельности железистого аппарата. Мионевральные синапсы представляют собой функциональную связь между аксоном мотонейрона и мышечными волокнами. На основании результатов электронно-микроскопических исследований в мионевральном синапсе различают три основные структуры: пресинаптическую мембрану, постсинаптическую мембрану и синаптическую щель (рис. 61).
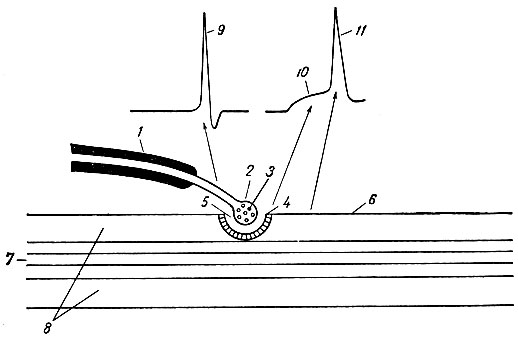
Рис. 61. Взаимоотношение между нервным волокном, нервным окончанием и скелетным мышечным волокном (схема). 1 - миелинизированное нервное волокно; 2 - нервное окончание с пузырьками медиатора; 3 - пресинаптическая мембрана; 4 - постсинаптическая мембрана мышечного волокна; 5 - синаптическая щель; 6 - внесинаптическая мембрана мышечного волокна; 7 - миофибриллы; 8 - саркоплазма; 9 - потенциал действия нервного волокна; 10 - потенциал концевой пластинки (постсинаптический потенциал); 11 - потенциал действия мышечного волокна
Пресинаптическая мембрана представляет собой электрогенную мембрану нервного окончания, аксоплазма которого включает большое количество гранул, или пузырьков, содержащих ацетилхолин.
Постсинаптическая мембрана, или концевая пластинка, - это электрогенная мембрана мышечного волокна, имеющая большое количество складок, что приводит к увеличению ее площади. Постсинаптическая мембрана содержит холинорецепторы, представляющие собой специальные белки мембраны. Кроме того, на постсинаптической мембране обнаружен фермент холинэстераза, который разрушает ацетилхолин,
Пресинаптическая и постсинаптическая мембраны разделяются узкой синаптической щелью - 20-50 нм (200-500 А), что обеспечивает быструю диффузию ацетилхолина. Синаптическая щель открывается во внеклеточное пространство и заполнена межклеточной жидкостью.
Передача возбуждения через мионевральный синапс осуществляется в результате выделения нервными окончаниями высокоактивных химических соединений, называемых медиаторами (посредниками, передатчиками нервного импульса). Предположение о химической передаче возбуждения через мионевральный синапс было высказано в 1924 г. русским физиологом А. Ф. Самойловым. Впоследствии было показано, что этим веществом является ацетилхолин.
Проведение возбуждения через мионевральный синапс осуществляется следующим образом. Кратковременная деполяризация пресинаптической мембраны, вызванная потенциалом действия нервного волокна, обеспечивает освобождение ацетилхолина из пузырьков терминали. Освобожденный ацетилхолин проникает через синаптическую щель к постсинаптической мембране и связывается с холинорецепторами. Указанное взаимодействие сопровождается повышением проницаемости постсинаптической мембраны для ионов натрия и калия, что обусловливает деполяризацию, которая проявляется в виде электроотрицательного возбуждающего постсинаптического потенциала (ВПСП). ВПСП имеет характер нераспространяющейся волны возбуждения, или локального ответа.
По мере развития ВПСП между деполяризованной постсинаптической мембраной и недеполяризованной мембраной мышечного волокна, примыкающей к ней, возникает разность потенциалов и появляются местные или круговые токи. При достижении ими порогового уровня в мембране мышечного волокна возникает распространяющийся мышечный потенциал действия.
Следует отметить, что связь ацетилхолина с холинорецептором непрочная. Молекулы ацетилхолина могут вступать в контакт с холинэстеразой, что приводит к распаду медиатора. В результате этого восстанавливается готовность синапса к проведению следующего нервного импульса.
Сильный и частый раздражитель приводит к выделению избыточных количеств ацетилхолина на пресинаптической мембране. Проникающий на постсинаптическую мембрану медиатор не успевает разрушаться холинэстеразой, что является причиной суммации постсинаптических потенциалов, возникновения стойкой деполяризации постсинаптической мембраны и блока проведения возбуждения.
Таким образом, в основе передачи возбуждения через мионевральный синапс лежат потенциал действия и сложные взаимодействия ацетилхолина с постсинаптической мембраной.
Синапсы обладают следующими основными физиологическими свойствами: односторонностью проведения возбуждения (в отличие от двустороннего проведения возбуждения в нервном волокне) и синаптической задержкой.
Одностороннее проведение возбуждения в синапсе связано с морфологическими его особенностями. Ацетилхолин вырабатывается в пресинаптической мембране, диффундирует через синаптическую щель и взаимодействует с холинорецептором, локализованным на постсинаптической мембране. Кроме того, синаптическая щель препятствует электрическому распространению возбуждения с нервного волокна на мышцу и с мышцы на нервное волокно.
Синаптическая задержка обусловливается следующими факторами: временем, необходимым для освобождения и диффузии медиатора через синаптическую щель, временем взаимодействия ацетилхолина с холинорецептором. В мионевральном синапсе задержка равна примерно 0,2-0,5 мс.
Н. Е. Введенский показал, что ответная реакция мышцы при увеличении силы или частоты раздражителя не может беспредельно возрастать.
На классическом объекте - нервно-мышечном препарате лягушки - было установлено, что при нарастании частоты раздражителя от 10 до 50 имп/с наблюдается увеличение амплитуды мышечного сокращения. Максимальная ответная реакция обнаруживалась при частоте 40-50 имп/с. Дальнейшее увеличение частоты раздражителя приводило к снижению амплитуды мышечного сокращения или к отсутствию механической реакции мышцы. Уменьшение частоты раздражителя сопровождалось восстановлением амплитуды мышечного сокращения.
Таким образом, Н. Е. Введенским было открыто и изучено совершенно новое явление в физиологии, которое свидетельствовало о том, что отсутствие ответной реакции мышцы на раздражение не связано с наступлением утомления в ней. Аналогичная закономерность в сократительном эффекте мышцы была установлена H. Е. Введенским и при увеличении силы раздражения, наносимого на двигательный нерв нервно-мышечного препарата лягушки.
Изменение реакции мышцы в зависимости от силы и частоты раздражителя позволило Н. Е. Введенскому дать определение оптимума и пессимума.
Оптимум (наилучший) - такая сила и частота раздражителя, которая обусловливает максимальную амплитуду мышечного сокращения.
Пессимум (наихудший) - такая чрезмерная (большая) сила и частота раздражителя, которая вызывает резкое уменьшение амплитуды мышечного сокращения или даже отсутствие механической реакции мышцы.
Снижение ответной реакции мышцы или ее отсутствие при пессимальном раздражении Н Е. Введенский связывал с понижением лабильности ткани. В результате снижения лабильности падает возбудимость, уменьшается скорость проведения возбуждения, удлиняется рефрактерный период. В итоге снижение лабильности приводит к блоку проведения возбуждения.
Пессимальная реакция, по Н. Е. Введенскому, развивается в синапсе как наиболее низколабильном образовании.
Современные представления о механизмах синаптической передачи позволяют трактовать механизм оптимума и пессимума следующим образом. При оптимальной частоте раздражителя каждый последующий нервный импульс попадает в фазу убывания постсинаптического потенциала, вызванного предыдущим нервным импульсом.
Нервные волокна (отростки нервных клеток) выполняют специализированную функцию - проведение нервных импульсов. По морфологическому признаку нервные волокна делятся на миелиновые (покрытые миелиновой оболочкой) и безмиелиновые. Нервные волокна формируют нерв или нервный ствол, состоящий из большого числа нервных волокон, заключенных в общую соединительноткан-ную оболочку. В состав нерва входят миелиновые и безмиелиновые волокна.
Нервные волокна, проводящие возбуждение от рецепторов в ЦНС называются афферентными, а волокна, проводящие возбуждение от ЦНС к исполнительным органам, называются эфферентными. Нервы состоят из афферентных и эфферентных волокон.
Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью.
Проведение возбуждения по нервным волокнам осуществляется по определенным законам.
Закон двустороннего проведения возбуждения по нервному волокну. Возбуждение по нервному волокну распространяется в обе стороны от места его возникновения, т. е. центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения.
Закон анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е. к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, т. е. не переходя с одного волокна на другое. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна.
Изолированное проведение возбуждения имеет важное значение. Нервсодержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки, ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно.
Нервные волокна по скорости проведения возбуждения, диаметру волок- на, длительности различных фаз потенциала действия и строению принято подразделять на три типа: А, В, С. Волокна типа А в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта.
Волокна типа А покрыты миелиновой оболочкой. Наиболее толстые из них А - альфа, они имеют диаметр 12-22 мкм и высокую скорость проведения возбуждения - 70-120 м/с. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам.
Три другие группы волокон тина А (бета, гамма, дельта) имеют меньший диаметр от 8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна этих групп относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых, рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам.
К волокнам типа В относятся миелинизированные преганглио-нарные волокна вегетативной нервной системы. Их диаметр - 1-3,5 мкм, а скорость проведения возбуждения - 3-18 м/с.
К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра - 0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более 3 м/с (0,5-3,0 м/с) . Большинство волокон типа С - это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
Механизм проведения возбуждения по нервным волокнам. Механизм распространения возбуждения у различных нервных волокон неодинаков. По современным представлениям распространение возбуждения по нервным волокнам осуществляется на основе ионных механизмов генерации потенциала действия.
При распространении возбуждения по безмиелиновому нервному волокну местные электрические токи,, которые возникают между его возбужденным участком, заряженным отрицательно, и невозбужденным, заряженным положительно, вызывают деполяризацию мембраны до критического уровня с последующей генерацией ПД в ближайшей точке невозбужденного участка мембраны. Этот процесс повторяется многократно. На всем протяжении нервного волокна происходит процесс репродукции нового ПД в каждой точке мембраны волокна. Такое проведение возбуждения называете и непрерывным.
Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки (перехватов Ранвье) создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Местные электрические токи возникают между соседними перехватами Ранвье, т. к. мембрана возбужденного перехвата становится заряженной отрицательно по отношению к поверхности соседнего невозбужденного перехвата. Эти местные токи деполярязуют мембрану невозбужденного перехвата до критического уровня и в нем возникает ПД (рис. 4). Следовательно, возбуждение как бы "перепрыгивает" через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Такой механизм распространения возбуждения называется сальтаторным или скачкообразным. Скорость такого способа проведения возбуждения значительно выше и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов.
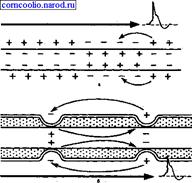
Рис. 4. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах.
"Перепрыгивание" потенциала действия через участок между перехватами возможно потому, что амплитуда ПД в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата. ПД может "перепрыгивать" не только через один, но и через два межперехватных промежутка. Это явление может наблюдаться при снижении возбудимости соседнего перехвата под действием какого-либо фармакологического вещества, например, новокаина, кокаина и др.
Нервные волокна обладают лабильностью - способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в единицу времени без трансформации ритма раздражения. Лабильность определяется длительностью пика потенциала действия, т. е. фазой абсолютной рефрактерности. Так как длительность абсолютной ре-фрактерности у спайкового потенциала нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000 импульсов в секунду.
Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности было названо Н. Е. Введенским парабиозом. В развитии состояния парабиоза можно отметить три, последовательно сменяющих друг друга, фазы: уравнительную, парадоксальную, тормозную.
В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители - больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100) , то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) - происходит уравнивание ответной реакции.
В парадоксальную-фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс - на редкие раздражители ответная реакция больше, чем на частые.
В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние.
Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ также связано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
Парабиоз - явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности.
Механизм развития парабиотического состояния сводится к следующему. При воздействии на нервное волокно парабиотического фактора нарушается способность мембраны увеличивать натриевую проницаемость в ответ на раздражение. В участке альтерации инак-тивация натриевых каналов, вызванная повреждающим агентом, суммируется с инактивацией, вызываемой нервным импульсом, и возбудимость снижается настолько, что проведение следующего импульса блокируется.
Читайте также:
