
Возбуждающие и тормозящие синапсы цнс
Взаимодействие нейронов между собой (и с эффекторными органами) происходит через специальные образования — синапсы (греч. — контакт). Они образуются концевыми разветвлениями нейрона на теле или отростках другого нейрона. Чем больше синапсов на нервной клетке, тем больше она воспринимает различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия в разнообразных реакциях организма. Особенно много синапсов в высших отделах нервной системы и именно у нейронов с наиболее сложными функциями.
В структуре синапса различаюттри элемента (рис. 2):
1)пресинаптическую мембрану, образованную утолщением мембраны конечной веточки аксона;
2)синаптическую щель между нейронами;
3)постсинаптическую мембрану — утолщение прилегающей поверхности следующего нейрона.
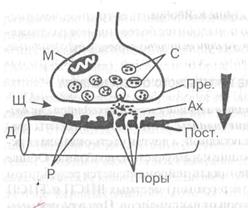
Рис. 2. Схема синапса
Пре. — пресинаптическая
мембрана, Пост. — постсинаптическая
С — синоптические пузырьки,
Щ—синоптическая щель,,
М — митохондрий, ;
Ах — ацетилхолин
Р — рецепторы и поры (Поры)
дендрита (Д) следующего
Стрелка — одностороннее проведение возбуждения.
В большинстве случаев передача влияния одного нейрона на другой осуществляется химическим путем. В пресинаптической части контакта имеются синоптические пузырьки, которые содержат специальные вещества — медиаторы или посредники. Ими могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных узлах), норадреналин (в окончаниях симпатических нервных волокон, в гипоталамусе), некоторые аминокислоты и др. Приходящие в окончания аксона нервные импульсы вызывают опорожнение синаптических пузырьков и выведение медиатора в синаптическую щель.
По характеру воздействия на последующую нервную клетку различают возбуждающие и тормозящие синапсы.
Ввозбуждающих синапсах медиаторы (например, ацетилхолин) связываются со специфическими макромолекулами постсинаптической мембраны и вызывают ее деполяризацию. При этом регистрируется небольшое и кратковременное (около 1мс) колебание мембранного потенциала в сторону делоляризации ил и возбуждающий постсинаптический потенциал (ВПСП). Для возбуждения нейрона необходимо, чтобы ВПСП достиг порогового уровня. Для этого величина деполяризационного сдвига мембранного потенциала должна составлять не менее 10 мВ. Действие медиатора очень кратковременно (1 -2 мс), после чего он расщепляется на неэффективные компоненты (например, ацетилхолин расщепляется ферментом холинэстеразой на холин и уксусную кислоту) ил и поглощается обратно пресинаптическими окончаниями (например, норадреналин).
В тормозящих синапсах содержатся тормозные медиаторы (например, гамма-аминомасляная кислота). Их действие на постсинаптическую мембрану вызывает усиление выхода ионов калия из клетки и увеличение поляризации мембраны. При этом регистрируется кратковременное колебание мембранного потенциала в сторону гиперполяризации — тормозящий постсинаптический потенциал (ТПСП). В результате нервная
клетка оказывается заторможенной. Возбудить ее труднее, чем в исходном состоянии. Для этого понадобится более сильное раздражение, чтобы достичь критического уровня деполяризации.
Функции нейронов и их типы. Физиология нервной ткани
Нейроны (нервные клетки) – основные структурные единицы нервной системы, через которые передается информация от одного участка НС к другому, обмен информацией между НС и различными участками тела.
1. Рецепторная – воспринимают внешние раздражения
2. Интегративная – осуществляют переработку информации о внешних раздражениях
3. Эффекторная – передают нервные влияния на другие нейроны или различные рабочие органы
1. Тело нейрона (сома) – в ней происходит переработка информации
2. Многочисленные древовидно разветвленные отростки дендриты – входы нейрона, через которые сигналы поступают в нервную клетку
3. Отходящий от тела клетки один отросток аксон – выход, который передает нервные импульсы дальше – к другой нервной клетке или рабочему органу (мышце, железе). Особо высокой возбудимостью обладает начальная часть аксона и расширение в месте его выхода из тела клетки – аксонный холмик нейрона, именно здесь и возникает нервный импульс.
1. Афферентные (чувствительные или центростремительные) – передают информацию от рецепторов в ЦНС. Тела этих нейронов расположены вне ЦНС – в спинномозговых узлах и в узлах черепных нервов. Дендрит у этих нейронов один и контактирует на периферии с воспринимающим сигнал образованием (рецептором) или сам образует рецептор, а аксон у него входит через задние рога в спинной мозг.
2. Эфферентные (центробежные) – передают нисходящие влияния от вышележащих этажей НС к нижележащим или из ЦНС к рабочим органам. У этих нейронов много коротких дендритов.
3. Промежуточные нейроны (интернейроны или вставочные) – более мелкие нервные клетки, осуществляющие связь между различными (в частности между афферентными и эфферентными) нейронами. Благодаря многогисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов.
Взаимодействие нейронов между собой (и с эффекторными органами) происходит через специальные образования – синапсы.
Рис. 6. Схема строения нейрона.
Обозначения:
I — сенсорный нейрон: 1 — окончания нейрона; 2 — аксон; 3 — ядро; 4 — тело клетки; 5— дендрит; 6 — миелиновая оболочка; 7 — рецептор; 8 — орган; 9 — неврилемма;
II — двигательный нейрон: 1 — дендриты; 2 — аксон; 3 — концевая бляшка; 4 — перехват Ранвье; 5 — ядро шванновской клетки; 6 — шванновская клетка;
III — вставочный нейрон: 1 — аксон; 2 — дендриты; 3 — ядро; 4 — тело клетки; 5 — дендрон
Синапсы - это специальные контакты, образованные концевыми разветвлениями нейрона на теле или отростках другого нейрона. Чем больше синапсов на нервной клетке, тем больше она воспринимает различных раздражений и тем шире сфера влияний на ее деятельность и возможность участия в разнообразных реакциях организма. Особенно много синапсов на нейронах, выполняющих наиболее сложные функции.
1. Пресинаптическая мембрана, образованная утолщением конечной веточки аксона.
2. Синаптическая щель между нейронами.
3. Постсинаптическая мембрана – утолщение прилегающей поверхности следующего нейрона.
Рис. 7. Структура синаптического контакта
В большинстве случаев передача влияния одного нейрона на другой осуществляется химическим путем. В пресинаптической части имеются синаптические пузырьки, которые содержат специальные вещества – медиаторы (ацетилхолин, аминокислоты, норадреналин и др). Приходящие в окончание аксона нервные импульсы вызывают сближение синаптических пузырьков с пресинаптической мембраной и выведение медиатора в синаптическую щель.
По характеру воздействия на последующую нервную клетку различают возбуждающие и тормозящие синапсы.
В возбуждающих синапсах – медиаторы (ацетилхолин) связываются со специфическими макромолекулами на постисинаптической мембране (рецепторами) и вызывают ее деполяризацию. При этом регистрируется небольшое кратковременное (около 1 мс) колебание мембранного потенциала в сторону деполяризации – возбуждающий постсинаптический потенциал (ВПСП). Для возбуждения нейрона необходимо, чтобы ВПСП достиг порогового уровня, а величина деполяризационного сдвига мембранного потенциала должна составлять не менее 10 мВ. Действие медиатора кратковременно (1-2 мс), после чего он расщепляется на неактивные компоненты или поглощается обратно пресинаптическими окончаниями.
В тормозящих синапсах содержатся тормозные медиаторы (ГАМК) и их действие на постсинаптическую мембрану вызывает усиление выхода ионов калия из клетки и увеличение поляризации мембраны. При этом регистрируется кратковременное колебание мембранного потенциала в сторону гиперполяризации – тормозный постсинаптический потенциал (ТПСП). В результате нервная клетка оказывается заторможенной и возбудить ее труднее, чем в исходном состоянии, так как необходимо более сильное раздражение, чтобы достичь критического уровня деполяризации.
Взаимодействие нейронов между собой (и с эффекторными органами) происходит через специальные образования - синапсы (греч. - контакт). Они образуются, в основном, концевыми разветвлениями нейрона на теле или отростках другого нейрона. Чем больше синапсов на нервной клетке, тем больше она воспринимает различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия в разнообразных реакциях организма. Особенно много синапсов в высших отделах нервной системы и именно у нейронов с наиболее сложными функциями.
Синапс- морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эф-фекторную клетку (мышечное волокно, секреторную клетку).
■ по конечному эффекту (по характеру воздействия на последующую нервную клетку) - тормозные и возбуждающие;
■ по механизму передачи сигнала - электрические(посредством ионов), химические(посредством медиаторов), смешанные.
В структуре синапса различают три элемента (рис. 5):
■ пресинаптическую мембрану, образованную утолщением мембраны конечной веточки аксона;
■ синаптическую щель между нейронами;
■ постсинаптическую мембрану - утолщение прилегающей поверхности следующего нейрона.
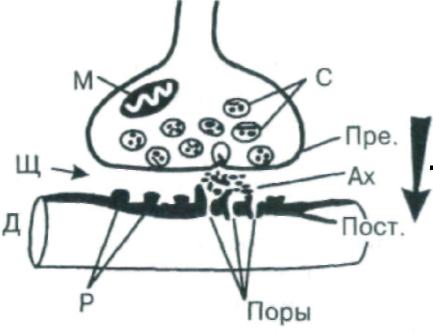
Рис. 5. Схема синапса: Пре. – пресинаптическая мембрана, Пост. – постсинаптическая мембрана, С – синаптические пузырьки, Щ – синаптическая щель, М – митохондрии, Ах – ацетилхолин, Р – рецепторы и поры (Поры) дендрита (Д) следующего нейрона; стрелка – одностороннее проведение возбуждения
В большинстве случаев передача влияния одного нейрона на другой осуществляется химическим путем. В пресинаптической части контакта имеются синаптические пузырьки, которые содержат специальные вещества – медиаторы,или посредники. Ими могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных узлах), норадреналин (в окончаниях симпатических нервных волокон, в гипоталамусе), некоторые аминокислоты и другие вещества. Приходящие в окончания аксона нервные импульсы вызывают опорожнение синаптических пузырьков и выведение медиатора в синаптическую щель.
В возбуждающих синапсах медиаторы (например, ацетилхолин) связываются со специфическими макромолекулами постсинаптической мембраны и вызывают ее деполяризацию. Эта деполяризация получила определенное название: возбуждающий постсинаптический потенциал (ВПСП).Для возбуждения нейрона необходимо, чтобы ВПСП достиг КУД. Для этого величина деполяризационного сдвига мембранного потенциала должна составлять не менее 10 мВ. Действие медиатора очень кратковременно (1 – 2 мс), после чего он или расщепляется на неэффективные компоненты (например, ацетилхолин расщепляется ферментом холинэстеразой на холин и уксусную кислоту), или поглощается обратно пресинап-тическими окончаниями (например, норадреналин).
В тормозящих синапсах содержатся тормозные медиаторы (например, гаммааминомасляная кислота). Их действие на постсинаптическую мембрану вызывает усиление выхода ионов калия из клетки, что приводит к гиперполяризации мембраны – регистрируется тормозящий постсинаптический потенциал (ТПСП).В результате нервная клетка оказывается заторможенной. Возбудить ее труднее, чем в исходном состоянии. Для этого понадобится более сильное раздражение, чтобы достичь критического уровня деполяризации.
На мембране тела и дендритов нервной клетки находятся как возбуждающие, так и тормозящие синапсы.
При одновременном влиянии как возбуждающих, так и тормозящих синапсов происходит алгебраическое суммирование (т.е. взаимное вычитание) их эффектов. При этом возбуждение нейрона возникнет лишь в том случае,если сумма возбуждающих постсинаптических потенциалов окажется больше суммы тормозящих.Это превышение должно составлять определенную пороговую величину (около 10 мВ). Только в этом случае появляется потенциал действия клетки. Следует отметить, что, в целом, возбудимость нейрона зависит от его размеров:чем меньше клетка,тем выше ее возбудимость.
С появлением потенциала действия начинается процесс проведения нервного импульса по аксону и передача его на следующий нейрон или рабочий орган, т.е. осуществляется эффекторная функция нейрона. Нервный импульс является основным средством связи между
Таким образом, передача информации в нервной системе происходит с помощью двух механизмов – электрического (ВПСП; ТПСП; потенциал действия) и химического (медиаторы).
2.Особенности проведения возбуждения в ЦНС (через нервные центры)
Свойства нервных центров в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, связывающие различные нервные клетки.
Нервным центром, как было сказано выше, называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния). В целостном организме имеется строгое согласование – координация их деятельности.
Проведение возбуждения в ЦНС имеет некоторые особенности.
1. Одностороннее проведение возбуждения.Проведение волны возбуждения от одного нейрона к другому через синапс происходит в большинстве нервных клеток химическим путем – с помощью медиатора, а медиатор содержится лишь в пресинаптической части синапса и отсутст-вует в постсинаптической мембране. Поэтому проведение нервных влияний возможно лишь от пресинаптической мембраны к постсинап-тической и невозможно в обратном направлении.В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление – от афферентных нейронов к вставочным и затем к эфферентным – мото-нейронам или вегетативным нейронам.
2. Большое значение в деятельности нервной системы имеет другая особенность проведения возбуждения через синапсы – замедленное прове-дение.Затрата времени на процессы, происходящие от момента подхода нервного импульса к пресинаптической мембране до появления в постси-наптической мембране потенциалов, называется синаптической задерж-кой.В большинстве центральных нейронов она составляет около 0,3 мс. После этого требуется еще время на развитие возбуждающего постсинапти-ческого потенциала (ВПСП) и потенциала действия. Весь процесс передачи нервного импульса (от потенциала действия одной клетки до потенциала действия следующей клетки) через один синапс занимает примерно 1,5 мс. При утомлении, охлаждении и ряде других воздействий длительность си-наптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам может составить десятые доли секунды и даже целые секунды.
При рефлекторной деятельности общее время от момента нанесения внешнего раздражения до появления ответной реакции организма — так называемое скрытое, или латентное, время рефлекса определяется в основ-ном длительностью проведения через синапсы. Величина латентного вре-мени рефлекса служит важным показателем функционального состояния нервных центров. Измерение латентного времени простой двигательной реакции человека на внешний сигнал широко используется в практике для оценки функционального состояния ЦНС.
3.Суммация возбуждения.В ответ на одиночную афферентную
волну, идущую от рецепторов к нейронам, в пресинаптической части синап-
са освобождается небольшое количество медиатора. При этом в постсин тической мембране нейрона обычно возникает ВПСП – небольшая местная деполяризация. Для того чтобы общая по всей мембране нейрона величина ВПСП достигала критического уровня возникновения потенциала действия, требуется суммация на мембране клетки многих подпороговых ВПСП. Лишь в результате такой суммации возбуждения возникает ответ нейрона. Различают пространственную и временную суммацию (рис. 6).

Рис. 6. Временная и пространственная суммация на уровне нейрона: 1 – приход импульсов к нервной клетке; 2 – формирование биопотенциала
Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным нервным волокнам. Возбуждения, приходящие в точку В, А, С нейрона (даже если они подпороговые) при одновременном появлении у данного нейрона могут привести к его возбуждению при условии, что суммирован-ный ВПСП достигает КУД.
Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражений. Если с определенным интервалом к нейрону в точку А приходят импульсы, они вызывают в этой области генерацию ВПСП. Если эти ВПСП не достигают критического уровня деполяризации, то ПД не возникает. Если же частота следования импульсов достаточно большая и ВПСП нейрона от предыдущих раздражений не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны нейрона не достигнет критического уровня для возникновения потенциала действия. Т.е. в этом месте происходит суммация ВПСП,при достижении ВПСП КУД возникает потенциал действия, нейрон возбуждается.
Таким способом даже слабые раздражения через некоторое время могут вызывать ответные реакции организма, например, чихания и кашля в ответ на слабые раздражения слизистой оболочки дыхательных путей.
4.Трансформация и усвоение ритма.Характер ответного разряда
нейрона зависит не только от свойств раздражителя, но и от функционального состояния самого нейрона (его мембранного заряда, возбудимости, лабильности). Нервные клетки обладают свойством изменять частоту
передающихся импульсов,т.е.свойством трансформации ритма.
При высокой возбудимости нейрона (например, после приема кофеина) может возникать учащение импульсации (мультипликация ритма), а при низкой возбудимости (например, при утомлении) происходит урежение ритма,т.к. несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения потенциала действия. Эти изменения частоты импульсации могут усиливать или ослаблять ответные реакции организма на внешние раздражения.
При ритмических раздражениях активность нейрона может настроиться на ритм приходящих импульсов, т.е. наблюдается явление усвоения ритма (А. А. Ухтомский, 1928). Развитие усвоения ритма обеспечивает сонастройку активности многих нервных центров при управлении сложными двигательными актами, особенно это важно для поддержания темпа циклических упражнений.
5. Следовые процессы.После окончания действия раздражителя активное состояние нервной клетки или нервного центра обычно продолжается еще некоторое время. Длительность следовых процессов различна:
небольшая в спинном мозге (несколько секунд или минут), значительно больше в центрах головного мозга (десятки минут, часы или даже дни) и очень большая в коре больших полушарий (до нескольких десятков лет).
Поддерживать явное и кратковременное состояние возбуждения в нервном центре могут импульсы, циркулирующие по замкнутым цепям нейронов. Значительно сложнее по природе длительно сохраняющиеся скрытые следы. Предполагают, что длительное сохранение в нервной клетке следов со всеми характерными свойствами раздражителя основано на изменении структуры составляющих клетку белков и на перестройке синаптических контактов.
Непродолжительные импульсные последействия (длительностью до 1 ч) лежат в основе так называемой кратковременной памяти, а длительные следы, связанные со структурными и биохимическими перестройками в клетках, – в основе формирования долговременной памяти.
Лекция 6КООРДИНАЦИЯ ДЕЯТЕЛЬНОСТИ ЦНС
Координация (дословно) – упорядочение, взаимосвязь, согласование. Координация – это объединение действий в единое целое. Управление!
Процессы координации деятельности ЦНС основаны на согласовании двух главных нервных процессов – возбуждения и торможения. Торможение является активным нервным процессом, который предупреждает или угнетает возбуждение.
Последнее изменение этой страницы: 2016-08-06; Нарушение авторского права страницы
Вопросы к зачету
2. Строение и основные свойства мембраны нейрона
Клеточная мембрана отделяет содержимое любой клетки от внешней среды, обеспечивая ее целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты илиорганеллы, в которых поддерживаются определенные условия среды.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой(если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
§ 1)барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
§ 2)транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального pH и концентрации ионов, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине неспособные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза.
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии по градиенту концентрации путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивает в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+).
§ 3)матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
§ 4)механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
§ 5)энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
§ 6)рецепторная — некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
§ 7)ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клетоккишечника содержат пищеварительные ферменты.
§ 8)осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты,глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс — одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних — активные процессы, связанные с потреблением энергии.
3.Морфофункциональная организация нейроглии
Среди глиоцитов, по морфологической организации, выделяют ряд типов:
- эпендимоциты
- астроциты
Эпендимоциты образуют плотный слой клеток, элементов, выстилающих спинномозговой канал и желудочки мозга. В процессе онтогенезе, эпендимоциты образовывались из спонгиобластов. Эпендимоциты представляют собой слегка вытянутые клетки с ветвящимися отростками. Некоторые эпендимоциты выполняют секреторную функцию, выделяя биологически активные вещества в кровь и в желудочки мозга. Эпендимоциты образуют скопления на капиллярной цепи желудочков мозга; при введении в кровь красителя, он накапливается эпендимоцитах, это свидетельствует о том, что последние выполняют функцию гематоэнцефалического барьера.
Астроциты выполняют опорную функцию. Это огромное количество глиальных клеток, имеющих множество коротких отростков. Среди астроцитов выделяют 2 группы:
- плазматические клетки
- волокнистые астроциты
Олигодендроциты – крупные глиальные клетки, часто сконцентрированы вокруг нервной клетки и поэтому называются сатиллитными глиацитами. Их функция очень важна для трофики нервной клетки. При функциональных перенапряжениях нервной клетки, глиоциты способны прореферировать вещества поступающие путем пиноцитоза в нервную клетку. При функциональных нагрузках, вначале происходит истощение синтетического аппарата глиальных клеток, а затем нервных. При восстановлении (репарации), вначале восстанавливаются функции нейронов, а затем – глиальных клеток. Таким образом, глиоциты принимают участи1е в обеспечении функций нейронов. Глиальные клетки существенным образом способны влиять на трофику мозга, а также на функциональный статус нервной клетки.
5.Синаптическая передача информации в ЦНС
1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся в пресинаптической терминали и концентрирующиеся в активных зонах пресинаптической мембраны.
2. Приходящий по аксону ПД деполяризует пресинаптическую мембрану.
3. Вследствие деполяризации открываются потенциалозависимые Са 2+ ‑каналы, и Са 2+ поступает в терминаль.
4. Увеличение внутриклеточного [Са 2+ ] запускает слияние синаптических пузырьков с пресинаптической мембраной и выброс нейромедиатора в синаптическую щель (экзоцитоз).
5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть молекул нейромедиатора связывается со специфичными для них рецепторами постсинаптической мембраны.
6. Связавшие нейромедиатор рецепторы активированы, что приводит к изменению поляризации постсинаптической мембраны либо прямо (поступление ионов через ионотропные рецепторы) либо опосредованно — активация ионных каналов через систему G‑белка (метаботропные рецепторы).
7. Инактивация нейромедиаторов происходит либо путём их ферментной деградации, либо молекулы нейромедиатора захватываются клетками.
6.Функции ионных каналов возбудимых мембран
Ионные каналы (ИК) - это мембранные молекулярные структуры, образованные интегральными (трансмембранными) белками, пронизывающими клеточную мембрану поперёк в виде нескольких петель и образующими в мембране сквозной канал (пору).ИК состоят из белков сложной структуры (белков-каналоформеров)
Практически все ИК имеют в составе своих субъединиц регуляторные домены, способные связываться с различными управляющими веществами (регуляторными молекулами) и за счёт этого менять состояние или свойства канала. В потенциал-активируемых ИК один из трансмембранных сегментов содержит специальный набор аминокислот с положительными зарядами и работает как сенсор электрического потенциала мембраны. При изменении потенциала такой сенсор меняет состояние канала с открытого на закрытое или наоборот. Таким образом, ИК могут управляться определёнными воздействиями извне, это важное их свойство.
ИК в своём составе могут иметь также вспомогательные субъединицы, выполняющие модуляторные, структурные или стабилизирующие функции. Один класс таких субъединиц - внутриклеточные, расположенные полностью в цитоплазме, а второй - мембранные, т.к. они имеют трансмембранные домены, прошивающие мембрану.
Свойства ИК:
1.Селективность -избирательная повышенная проницаемость ИК для определённых ионов
2.Управляемая проницаемость - это способность ИК открываться или закрываться при определённых управляющих воздействиях на канал.
3.Инактивация - это способность ИК через некоторое время после своего открытия автоматически понижать свою проницаемость
4.Блокировка - это способность ИК под действием веществ-блокаторов фиксировать какое-то одно своё состояние и не реагировать на обычные управляющие воздействия
5.Пластичность - это способность ИК изменять свои свойства, свои характеристики.
Функции ИК:
1. Регуляция водного обмена клетки: объём и тургор.
2. Регуляция pH: закисление и защелачивание.
3. Регуляция ионного обмена (обмен солей): изменение внутриклеточного ионного состава и концентрации.
4. Создание и изменение мембранных потенциалов: потенциал покоя; в возбудимых клетках - локальные потенциалы, потенциал действия.
5. Проведение возбуждения в возбудимых клетках: обеспечение движения нервных импульсов.
6. Трансдукция в сенсорных рецепторах: преобразование раздражения (стимула) в возбуждение.
7. Управление активностью клетки: за счёт обеспечения потоков вторичного мессенджера - Са2+.
Главная функция ИК - обеспечивать управляемое перемещение ионов через мембрану.
Функциональные состояния ИК:
1. Открытое. Канал открыт и через него происходит перемещение ионов.
2. Закрытое. Канал закрыт и ионы не проходят через него.
3. Активированное. Канал может выполнять свои функции, т.е. открываться и закрываться под действием его регуляторов (управляющих веществ или электрических потенциалов).
4. Инактивированное. Канал не может выполнять свои функции, т.е. открываться и закрываться, он "фиксируется" в каком-то одном состоянии.
5. Блокированное. Канал перекрыт, инактивирован веществом-антагонистом (блокатором), занявшем место управляющего вещества.
6. Модулированное (фосфорилированное). Канал изменяет свои обычные свойства под действием фосфорилирования - присоединения к какому-то его участку фосфатного остатка.
Возбуждающие и тормозные синапсы. Электрический синапс.
Синапс возбуждающий —синапс, в котором в результате деполяризации постсинаптической мембраны генерируется возбуждающий постсинаптический потенциал.
Синапс тормозной —
1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала;
2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение.
Возбуждение
1. Открытие натриевых каналов позволяет большому числу положительных электрических зарядов войти в постсинаптическую клетку. Это сдвигает внутриклеточный мембранный потенциал в положительном направлении, приближая его к пороговому для возбуждения уровню. Это наиболее широко используемый способ для вызова возбуждения.
2. Снижение проводимости через хлорные или калиевые каналы, или через те и другие уменьшает диффузию отрицательно заряженных ионов Сl- внутрь постсинаптического нейрона или снижает диффузию положительно заряженных ионов К+ наружу. В любом случае результатом будет поддержание более положительного, чем в норме, мембранного потенциала, что способствует возбуждению.
3. Различные изменения внутриклеточного метаболизма постсинаптического нейрона ведут к возбуждению клеточной активности или в некоторых случаях — к увеличению числа возбуждающих или уменьшению числа тормозных мембранных рецепторов.
Торможение
1. Открытие каналов для ионов хлора в постсинаптической мембране нейрона позволяет отрицательно заряженным ионам быстро диффундировать снаружи внутрь постсинаптического нейрона, увеличивая таким образом отрицательность внутри нейрона. Это тормозной эффект.
2. Увеличение проводимости мембраны для ионов калия позволяет положительным ионам диффундировать наружу, что ведет к увеличению отрицательности внутри нейрона. Это также является тормозным эффектом.
3. Активация ферментов, отвечающих за клеточные метаболические функции, которые увеличивают число тормозных рецепторов или уменьшают количество возбуждающих синаптических рецепторов.
Синапс электрический - в нем возбуждение от пре- к постсинаптической мембране передается электрическим путем, т.е. совершается эфаптическая передача возбуждения - потенциал действия достигает пресинаптического окончания и далее распространяется по межклеточным каналам, вызывая деполяризацию постсинаптической мембраны. В электрическом синапсе медиатор не вырабатывается, синаптическая щель мала (2 - 4 нм) и в ней имеются белковые мостики-каналы, шириной 1 - 2 нм, по которым движутся ионы и небольшие молекулы.
10. ТИПЫ НЕРВНЫХ ВОЛОКОН И СКОРОСТИ ПРОВЕДЕНИЯ ПО НИМ ВОЗБУЖДЕНИЯ.
По особенностям строения и функциям нервные волокна подразделяются на два вида: безмиелиновые и миелиновые. В зависимости от скорости проведения возбуждения нервные волокна делятся на три типа: А, В, С.
А – миелинизированные, наибольшего диаметра, скорость 120-70 м в сек, длительность ПД – минимальная, делятся на подгруппы: альфа, бета, гамма, дельта. Миелиновая оболочка:α- Очень толстая,β- Толстая,γ -Умеренная, δ-Тонкая.Пример – аксоны мотонейронов.
В – миелинизированные волокна, меньшего диаметра, скорость 3-18 м в сек, ПД более длителен. Миелиновая оболочка: Прерывистая.Пример – преганглионарные волокна симпатической нервной системы.
С – немиелизированные нервные волокна, скорость менее 2 м в сек, длительность ПД наибольшая. Миелиновая оболочка: Отсутствует. Пример - постганглионарные волокна парасимпатической нервной системы.
11. Функции вегетативной нервной системы
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА - автономная нервная система.
Вегетативная нервная система контролирует деятельность важнейших органов и систем человека. Она регулирует все функции сердца и кровеносных сосудов, например, при занятиях спортом отдельными мышцам необходимо больше крови, поэтому при воздействии нервных импульсов увеличивается количество сокращений сердца и расширяются кровеносные сосуды. В это же время нервная система усиливает и дыхание, чтобы кровь могла отнести больше кислорода к мышцам, на которых приходится большая нагрузка. Подобным образом вегетативная нервная система регулирует и температуру тела. Избыток тепла удаляется интенсивным кровообращением кожи.
12. Рефлекторный принцип функционирования спинного мозга.
Принцип работы сегментарного аппарата спинного мозга — рефлекторные дуги.
Основная схема рефлекторной дуги спинного мозга: информация от рецептора идет по чувствительному нейрону, тот переключается на вставочный нейрон, тот в свою очередь на мотонейрон, который несет информацию к эффекторному органу. Для рефлекторной дуги характерен сенсорный вход, непроизвольность, межсегментарность, моторный выход.
Примерами спинномозговых рефлексов могут служить: Сгибательный (флексорный) рефлекс — рефлекс защитного типа направленный на удаление повреждающего раздражителя (отдергивание руки от горячего). Рефлекс на растяжения (проприоцептивный) — предотвращающий чрезмерное растяжение мышцы. Особенностью этого рефлекса является, то что рефлекторная дуга содержит минимум элементов — мышечные веретена генерируют импульсы которые проходят в спинной мозг и вызывают моносинаптическое возбуждение в α-мотонейронах той же мышцы.Сухожильный, разнообразные тонические и ритмические рефлексы.У четвероногих животных можно наблюдать экстензорный толчок.
Дата добавления: 2018-04-05 ; просмотров: 1009 ;
Читайте также:
