
Возбуждение дыхательного центра углекислым газом крови нервная
Дыхательный центр не только обеспечивает ритмическое чередование вдоха и выдоха, но и способен изменять глубину и частоту дыхательных движений, приспосабливая тем самым легочную вентиляцию к текущим потребностям организма. Факторы внешней среды, например состав и давление атмосферного воздуха, окружающая температура, и изменения состояния организма, например при мышечной работе, эмоциональном возбуждении и др., влияя на интенсивность обмена веществ, а, следовательно, потребление кислорода и выделение углекислого газа, действуют на функциональное состояние дыхательного центра. В результате меняется объем легочной вентиляции.
Как и все другие процессы автоматической регуляции физиологических функций, регуляция дыхания осуществляется в организме на основе принципа обратной связи. Это значит, что деятельность дыхательного центра, регулирующего снабжение организма кислородом и удаление образующегося в нем углекислого газа, определяется состоянием регулируемого им процесса. Накопление в крови углекислоты, а также недостаток кислорода являются факторами, вызывающими возбуждение дыхательного центра.
Значение газового состава крови в регуляции дыхания было показано Фредериком путем опыта с перекрестным кровообращением. Для этого у двух собак, находившихся под наркозом, перерезали и соединяли перекрестно их сонные артерии и отдельно яремные вены (рисунок 2) После такого соединения этих и зажатия других сосудов шеи голова первой собаки снабжалась кровью не от собственного туловища, а от туловища второй собаки, голова же второй собаки - от туловища первой.
Если у одной из этих собак зажать трахею и таким образом производить удушение организма, то через некоторое время у нее происходит остановка дыхания (апноэ), у второй же собаки возникает резкая одышка (диспноэ). Это объясняется тем, что зажатие трахеи у первой собаки вызывает накопление СО2 в крови ее туловища (гиперкапния) и уменьшение содержания кислорода (гипоксемия). Кровь из туловища первой собаки поступает в голову второй собаки и стимулирует ее дыхательный центр. В результате возникает усиленное дыхание - гипервентиляция - у второй собаки, что приводит к снижению напряжения СО2 и повышению напряжения О2 в крови сосудов туловища второй собаки. Богатая кислородом и бедная углекислым газом кровь из туловища этой собаки поступает в голову первой и вызывает у нее апноэ.
Рисунок 2 - Схема опыта Фредерика с перекрестным кровообращением
Опыт Фредерика показывает, что деятельность дыхательного центра изменяется при изменении напряжения СО2 и О2 в крови. Рассмотрим влияние на дыхание каждого из этих газов в отдельности.
Значение напряжения углекислого газа в крови в регуляции дыхания. Повышение напряжения углекислого газа в крови вызывает возбуждение дыхательного центра, приводящее к увеличению вентиляции легких, а понижение напряжения углекислого газа в крови угнетает деятельность дыхательного центра, что приводит к уменьшению вентиляции легких. Роль углекислого газа в регуляции дыхания доказана Холденом в опытах, в которых человек находился в замкнутом пространстве небольшого объема. По мере того как во вдыхаемом воздухе уменьшается содержание кислорода и увеличивается содержание углекислого газа, начинает развиваться диспноэ. Если же поглощать выделяющийся углекислый газ натронной известью, содержание кислорода во вдыхаемом воздухе может снизиться до 12%, причем заметного увеличения легочной вентиляции не наступает. Таким образом, увеличение объема вентиляции легких в этом опыте обусловлено повышением содержания во вдыхаемом воздухе углекислого газа.
Результаты экспериментов дали убедительное доказательство того, что состояние дыхательного центра зависит от содержания углекислого газа в альвеолярном воздухе. Выявлено, что увеличение содержания СО2 в альвеолах на 0,2% вызывает увеличение вентиляции легких на 100%.
Уменьшение содержания углекислого газа в альвеолярном воздухе (и, следовательно, уменьшение напряжения его в крови) понижает деятельность дыхательного центра. Это происходит, например, в результате искусственной гипервентиляции, т. е. усиленного глубокого и частого дыхания, которое приводит к снижению парциального давления СО2 в альвеолярном воздухе и напряжения СО2 в крови. В результате наступает остановка дыхания. Пользуясь таким способом, т. е. производя предварительную гипервентиляцию, можно значительно увеличить время произвольной задержки дыхания. Так поступают ныряльщики, когда им нужно провести под водой 2…3 минуты (обычная длительность произвольной задержки дыхания составляет 40…60 секунд).
На дыхательный центр оказывает влияниеповышение концентрации водородных ионов.Винтерштейн в 1911 г. высказал точку зрения, что возбуждение дыхательного центра вызывает не сама угольная кислота, а, повышение концентрации водородных ионов вследствие увеличения ее содержания в клетках дыхательного центра.
Стимулирующее влияние углекислого газа на дыхательный центр является основанием одного мероприятия, нашедшего применение в клинической практике. При ослаблении функции дыхательного центра и возникающем при этом недостаточном снабжении организма кислородом больного заставляют дышать через маску смесью кислорода с 6% углекислого газа. Такая газовая смесь носит название карбогена.
Значение хеморецепторов продолговатого мозга видно из следующих фактов. При воздействии на эти хеморецепторы углекислого газа или растворов с повышенной концентрацией Н+-ионов наблюдается стимуляция дыхания. Охлаждение одного из хеморецепторных телец продолговатого мозга влечет за собой, согласно опытам Лешке, прекращение дыхательных движений на противоположной стороне тела. Если хеморецепторные тельца разрушены или отравлены новокаином, дыхание прекращается.
Наряду схеморецепторами продолговатого мозга в регуляции дыхания важная роль принадлежит хеморецепторам, находящимся в каротидном и аортальном тельцах. Это было доказано Геймансом в методически сложных опытах, в которых сосуды двух животных соединялись так, что каротидный синус и каротидное тельце или дуга аорты и аортальное тельце одного животного снабжались кровью другого животного. Оказалось, что увеличение концентрации Н + -ионов в крови и повышение напряжения СО2 вызывают возбуждение каротидных и аортальных хеморецепторов и рефлекторное усиление дыхательных движений.
Рассмотрим влияние недостатка кислорода на дыхание.Возбуждение инспираторных нейронов дыхательного центра возникает не только при повышении напряжения углекислого газа в крови, но и при понижении напряжения кислорода.
Характер изменения дыхания при избытке углекислоты и понижении напряжения кислорода в крови различен. При небольшом понижении напряжения кислорода в крови наблюдается рефлекторное учащение ритма дыхания, а при незначительном повышении напряжения углекислоты в крови происходит рефлекторное углубление дыхательных движений.
Таким образом, деятельность дыхательного центра регулируется воздействием повышенной концентрации Н+-ионов и увеличенного напряжения СО2 на хеморецепторы продолговатого мозга и на хеморецепторы каротидного и аортального телец, а также действием на хеморецепторы указанных
Значение механорецепторов в регуляции дыхания.Дыхательный центр получает афферентные импульсы не только от хеморецепторов, но и от прессорецепторов сосудистых рефлексогенных зон, а также от механорецепторов легких, дыхательных путей и дыхательных мышц.
Влияние прессорецепторов сосудистых рефлексогенных зон обнаруживается в том, что повышение давления в изолированном каротидном синусе, связанном с организмом только нервными волокнами, приводит к угнетению дыхательных движений. Это происходит и в организме при повышении артериального давления. Наоборот, при понижении артериального давления дыхание учащается и углубляется.
Важное значение в регуляции дыхания имеют импульсы, поступающие к дыхательному центру по блуждающим нервам от рецепторов легких. От них в значительной степени зависит глубина вдоха и выдоха. Наличие рефлекторных влияний с легких было описано в 1868 г. Герингом и Брейером и легло в основу представления о рефлекторной саморегуляции дыхания. Она проявляется в том, что при вдохе в рецепторах, находящихся в стенках альвеол, возникают импульсы, рефлекторно тормозящие вдох, и стимулирующих выдох, а при очень резком выдохе, при крайней степени уменьшения объема легких возникают импульсы, поступающие к дыхательному центру и рефлекторно стимулирующие вдох. О наличии такой рефлекторной регуляции свидетельствуют следующие факты:
- в легочной ткани в стенках альвеол, т. е. в наиболее растяжимой части легкого, имеются интерорецепторы, представляющие собой воспринимающие раздражения окончания афферентных волокон блуждающего нерва;
- после перерезки блуждающих нервов дыхание становится резко замедленным и глубоким;
- при раздувании легкого индифферентным газом, например азотом, при обязательном условии целости блуждающих нервов, мускулатура диафрагмы и межреберий внезапно перестает сокращаться, вдох останавливается, не достигнув обычной глубины; наоборот, при искусственном отсасывании воздуха из легкого наступает сокращение диафрагмы.
Помимо механорецепторов легких, в регуляции дыхания принимают участие механорецепторы межреберных мышц и диафрагмы. Они возбуждаются растяжением при выдохе и рефлекторно стимулируют вдох (С. И. Франштейн).
Соотношения между инспираторными и экспираторными нейронами дыхательного центра. Между инспираторными и экспираторными нейронами существуют сложные реципрокные (сопряженные) соотношения. Это означает, что возбуждение инспираторных нейронов тормозит экспираторные, а возбуждение экспираторных нейронов тормозит инспиряторные. Такие явления частично обусловлены наличием прямых связей, существующих между нейронами дыхательного центра, но в основном они зависят от рефлекторных влияний и от функционирования центра пневмотаксиса.
Взаимодействие между нейронами дыхательного центра в настоящее время представляют следующим образом. Вследствие рефлекторного (через хеморецепторы) действия углекислоты на дыхательный центр возникает возбуждение инспираторных нейронов, которое передается на мотонейроны, иннервирующие дыхательные мышцы, вызывая акт вдоха. Одновременно импульсы от инспираторных нейронов поступают к центру пневмотаксиса, расположенному в варолиевом мосту, а от него по отросткам его нейронов импульсы приходят к экспираторным нейронам дыхательного центра продолговатого мозга, вызывая возбуждение этих нейронов, прекращение вдоха и стимуляцию выдоха. Кроме того, возбуждение экспираторных нейронов во время вдоха осуществляется и рефлекторно посредством рефлекса Геринга - Брейера. После перерезки блуждающих нервов приток импульсов от механорецепторов легких прекращается и экспираторные нейроны могут возбуждаться лишь посредством импульсов, приходящих из центра пневмотаксиса. Импульсация, возбуждающая центр выдоха, значительно уменьшается и возбуждение его несколько запаздывает. Поэтому после перерезки блуждающих нервов вдох продолжается значительно дольше и сменяется выдохом позднее, чем до перерезки нервов. Дыхание становится редким и глубоким.
Таким образом, жизненно важная функция дыхания, возможная лишь при ритмическом чередовании вдоха и выдоха, регулируется сложным нервным механизмом. При его изучении обращает на себя внимание множественное обеспечение работы этого механизма. Возбуждение центра вдоха возникает как под влиянием увеличения концентрации водородных ионов (повышения напряжения СО2) в крови, вызывающего возбуждение хеморецепторов продолговатого мозга и хеморецепторов сосудистых рефлексогенных зон, так и в результате влияния пониженного напряжения кислорода на аортальные и каротидные хеморецепторы. Возбуждение центра выдоха обусловлено как рефлекторными импульсами, приходящими к нему по афферентным волокнам блуждающих нервов, так и влиянием центра вдоха, осуществляемым через центр пневмотаксиса.
Возбудимость дыхательного центра изменяется при действии нервных импульсов, поступающих по шейному симпатическому нерву. Раздражение этого нерва повышает возбудимость центра дыхания, что усиливает и учащает дыхание.
Влиянием симпатических нервов на дыхательный центр отчасти объясняются изменения дыхания при эмоциях.
Установите правильную последовательность процессов нормальных вдоха и выдоха у человека, начиная с повышения концентрации СО2 в крови.
Запишите в таблицу соответствующую последовательность цифр.
1) сокращение диафрагмы
2) повышение концентрации кислорода
3) повышение концентрации СО2
4) возбуждение хеморецепторов продолговатого мозга
6) расслабление диафрагмы
Последовательность процессов нормальных вдоха и выдоха у человека, начиная с повышения концентрации СО2 в крови:
3) повышение концентрации СО2→4) возбуждение хеморецепторов продолговатого мозга→6) расслабление диафрагмы→1) сокращение диафрагмы→2) повышение концентрации кислорода→5) выдох
Дыхательный центр находится в продолговатом мозге. Под действием углекислого газа крови в нем возникает возбуждение, оно передается к дыхательным мышцам, происходит вдох. При этом возбуждаются рецепторы растяжения в стенках легких, они посылают тормозящий сигнал в дыхательный центр, он перестает посылать сигналы к дыхательным мышцам, происходит выдох.
Если задержать дыхание надолго, то углекислый газ будет все сильнее возбуждать дыхательный центр, в конце концов дыхание возобновится непроизвольно.
Кислород не влияет на дыхательный центр. При избытке кислорода (при гипервентиляции) происходит спазм сосудов мозга, что приводит к головокружению или обмороку.
Т.к. данное задание вызывает много споров, о том, что последовательность в ответе не корректная - принят решение отправить данное задание в неиспользуемые.
Кто хочет подробнее узнать о механизмах регуляции дыхания можно почитать статью "Физиология системы дыхания". О хеморецепторах в самом конце статьи.
Дыхательный центр
Под дыхательным центром следует понимать совокупность нейронов специфических (дыхательных) ядер продолговатого мозга, способных генерировать дыхательный ритм.
В нормальных (физиологических) условиях дыхательный центр получает афферентные сигналы от периферических и центральных хеморецепторов, сигнализирующих соответственно о парциальном давлении О2 в крови и концентрации Н + во внеклеточной жидкости мозга. В период бодрствования деятельность дыхательного центра регулируется дополнительными сигналами, исходящими из различных структур ЦНС. У человека это, например, структуры, обеспечивающие речь. Речь (пение) может в значительной степени отклонить от нормального уровень газов крови, даже снизить реакцию дыхательного центра на гипоксию или гиперкапнию. Афферентные сигналы от хеморецепторов тесно взаимодействуют с другими афферентными стимулами дыхательного центра, но, в конечном счете, химический, или гуморальный, контроль дыхания всегда доминирует над нейрогенным. Например, человек произвольно не может бесконечно долго задерживать дыхание из-за нарастающих во время остановки дыхания гипоксии и гиперкапнии.
Ритмическая последовательность вдоха и выдоха, а также изменение характера дыхательных движений в зависимости от состояния организма регулируются дыхательным центром, расположенным в продолговатом мозге.
В дыхательном центре имеются две группы нейронов: инспираторные и экспираторные. При возбуждении инспираторных нейронов, обеспечивающих вдох, деятельность экспираторных нервных клеток заторможена, и наоборот.
В верхней части моста головного мозга (варолиев мост) находится пневмотаксический центр, который контролирует деятельность расположенных ниже центров вдоха и выдоха и обеспечивает правильное чередование циклов дыхательных движений.
Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровне III—IV шейных сегментов спинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах (III—XII) грудных сегментов спинного мозга.
Дыхательный центр выполняет две основные функции в системе дыхания: моторную, или двигательную, которая проявляется в виде сокращения дыхательных мышц, и гомеостатическую, связанную с изменением характера дыхания при сдвигах содержания О2 и СО2 во внутренней среде организма.
Диафрагмальные мотонейроны. Образуют диафрагмальный нерв. Нейроны расположены узким столбом в медиальной части вентральных рогов от СIII до CV. Диафрагмальный нерв состоит из 700—800 миелинизированных и более 1500 немиелинизированных волокон. Подавляющее количество волокон является аксонами α-мотонейронов, а меньшая часть представлена афферентными волокнами мышечных и сухожильных веретен, локализованных в диафрагме, а также рецепторов плевры, брюшины и свободных нервных окончаний самой диафрагмы.
Мотонейроны сегментов спинного мозга, иннервирующие дыхательные мышцы. На уровне CI—СII вблизи латерального края промежуточной зоны серого вещества находятся инспираторные нейроны, которые участвуют в регуляции активности межреберных и диафрагмальных мотонейронов.
Мотонейроны, иннервирующие межреберные мышцы, локализованы в сером веществе передних рогов на уровне от TIV до ТX. Причем одни нейроны регулируют преимущественно дыхательную, а другие — преимущественно позно-тоническую активность межреберных мышц. Мотонейроны, иннервирующие мышцы брюшной стенки, локализованы в пределах вентральных рогов спинного мозга на уровне TIV—LIII.
Генерация дыхательного ритма.
Спонтанная активность нейронов дыхательного центра начинает появляться к концу периода внутриутробного развития. Об этом судят по периодически возникающим ритмическим сокращениям мышц вдоха у плода. В настоящее время доказано, что возбуждение дыхательного центра у плода появляется благодаря пейсмекерным свойствам сети дыхательных нейронов продолговатого мозга. Иными словами, первоначально дыхательные нейроны способны самовозбуждаться. Этот же механизм поддерживает вентиляцию легких у новорожденных в первые дни после рождения. С момента рождения по мере формирования синаптических связей дыхательного центра с различными отделами ЦНС пейсмекерный механизм дыхательной активности быстро теряет свое физиологическое значение. У взрослых ритм активности в нейронах дыхательного центра возникает и изменяется только под влиянием различных синаптических воздействий на дыхательные нейроны.
Дыхательный цикл подразделяют на фазу вдоха и фазу выдоха относительно движения воздуха из атмосферы в сторону альвеол (вдох) и обратно (выдох).
Двум фазам внешнего дыхания соответствуют три фазы активности нейронов дыхательного центра продолговатого мозга: инспираторная, которая соответствует вдоху; постинспираторная, которая соответствует первой половине выдоха и называется пассивной контролируемой экспирацией; экспираторная, которая соответствует второй половине фазы выдоха и называется фазой активной экспирации .
Активность дыхательных мышц в течение трех фаз нейронной активности дыхательного центра изменяется следующим образом. В инспирацию мышечные волокна диафрагмы и наружных межреберных мышц постепенно увеличивают силу сокращения. В этот же период активируются мышцы гортани, которые расширяют голосовую щель, что снижает сопротивление воздушному потоку на вдохе. Работа инспираторных мышц во время вдоха создает достаточный запас энергии, которая высвобождается в постинспираторную фазу, или в фазу пассивной контролируемой экспирации. В постинспираторную фазу дыхания объем выдыхаемого из легких воздуха контролируется медленным расслаблением диафрагмы и одновременным сокращением мышц гортани. Сужение голосовой щели в постинспираторную фазу увеличивает сопротивление воздушному потоку на выдохе. Это является очень важным физиологическим механизмом, который препятствует спадению воздухоносных путей легких при резком увеличении скорости воздушного потока на выдохе, например при форсированном дыхании или защитных рефлексах кашля и чиханья.
Во вторую фазу выдоха, или фазу активной экспирации, экспираторный поток воздуха усиливается за счет сокращения внутренних межреберных мышц и мышц брюшной стенки. В эту фазу отсутствует электрическая активность диафрагмы и наружных межреберных мышц.
Регуляция деятельности дыхательного центра.
Регуляция деятельности дыхательного центра осуществляется с помощью гуморальных, рефлекторных механизмов и нервных импульсов, поступающих из вышележащих отделов головного мозга.
Гуморальные механизмы. Специфическим регулятором активности нейронов дыхательного центра является углекислый газ, который действует на дыхательные нейроны непосредственно и опосредованно. В ретикулярной формации продолговатого мозга, вблизи дыхательного центра, а также в области сонных синусов и дуги аорты обнаружены хеморецепторы, чувствительные к углекислому газу. При увеличении напряжения углекислого газа в крови хеморецепторы возбуждаются, и нервные импульсы поступают к инспираторным нейронам, что приводит к повышению их активности.
Дыхание регулируется дыхательным центром, который находится в спинном, продолговатом мозге, мосту, гипоталамусе, коре больших полушарий.
Ведущая роль в организации дыхания принадлежит центру продолговатого мозга, который состоит из центров вдоха (инспираторные нейроны) и выдоха (экспираторные нейроны). Разрушение этой области ведет к остановке дыхания. Здесь находятся нейроны, обеспечивающие ритмичность вдоха и выдоха. Это связано с тем, что дыхательный центр обладает свойством автоматии, т.е. его нейроны способны ритмически самовозбуждаться. Автоматия сохраняется, даже если к дыхательному центру не поступают нервные импульсы по центростремительным нейронам. Автоматия может изменяться в зависимости от гуморальных факторов, нервных импульсов, поступающих по центростремительным нейронам и под влиянием вышележащих отделов мозга. От дыхательного центра нервные импульсы по центробежным нейронам подходят к межреберным мышцам, диафрагме и др. мышцам.
Регуляция дыхания осуществляется с помощью гуморальных, рефлекторных механизмов и нервных импульсов, поступающих из вышележащих отделов головного мозга.
Специфическим регулятором активности нейронов дыхательного центра является углекислый газ, который действует на дыхательные нейроны непосредственно и опосредованно. Углекислый газ непосредственно возбуждает инспираторные клетки дыхательного центра. В механизме стимулирующего влияния углекислого газа на дыхательный центр важное место принадлежит хеморецепторам сосудистого русла. В области сонных синусов и дуги аорты обнаружены хеморецепторы, чувствительные к изменениям напряжения углекислого газа в крови. Кстати, первый вдох новорожденного объясняется действием накопившейся в его тканях углекислого газа на дыхательный центр (после перерезки пуповины и отделения от организма матери). Это действие является как прямым, так и опосредованным, рефлекторным – через хеморецепторы сонного синуса и дуги аорты. Избыток углекислого газа в крови вызывает одышку. Недостаток кислорода в крови углубляет дыхание. Установлено, что повышение напряжения кислорода в крови тормозит активность дыхательного центра.
Рефлекторные механизмы. Различают постоянные и непостоянные рефлекторные влияния на функциональное состояние дыхательного центра. Постоянные рефлекторные влияния возникают в результате раздражения рецепторов альвеол (рефлекс Э. Геринга – И. Брейера), корня легкого и плевры (плевропульмональный рефлекс), хеморецепторов дуги аорты и сонных синусов (рефлекс К. Гейманса), проприорецепторов дыхательных мышц.
Рефлекс Э. Геринга и И. Брейера называют рефлексом торможения вдоха при растяжении легких. При вдохе возникают импульсы, тормозящие вдох и стимулирующие выдох, а при выдохе – импульсы, рефлекторно стимулирующие вдох. Регуляция дыхательных движений осуществляется по принципу обратной связи. При перерезке блуждающих нервов рефлекс выключается, дыхание становится редким и глубоким.
Непостоянные рефлекторные влияния на активность дыхательных нейронов связаны с возбуждением разнообразных экстерорецепторов и интерорецепторов. Так, например, при внезапном вдыхании паров аммиака, хлора, табачного дыма и некоторых других веществ происходит раздражение рецепторов слизистой оболочки носа, глотки, гортани, что приводит к рефлекторному спазму голосовой щели (иногда даже мускулатуры бронхов) и рефлекторной задержке дыхания. Сильные температурные воздействия на кожу возбуждают дыхательный центр, увеличивают вентиляцию легких. Резкое охлаждение угнетает дыхательный центр. На дыхание влияют боль, импульсы от сосудистых барорецепторов: так, повышение артериального давления угнетает дыхательный центр, что проявляется уменьшением глубины и частоты дыхания.
При раздражении эпителия дыхательных путей накопившейся пылью, слизью, химическими раздражителями и инородными телами возникают чиханье и кашель (защитные врожденные рефлексы). Чиханье возникает при раздражении рецепторов слизистой оболочки носа, кашель – при возбуждении рецепторов гортани, трахеи, бронхов.
Первый уровень регуляции – спинной мозг. Здесь располагаются центры диафрагмальных и межреберных нервов, обусловливающие сокращение дыхательных мышц. Однако этот уровень регуляции дыхания не может обеспечить ритмичную смену фаз дыхательного аппарата.
Второй уровень регуляции – продолговатый мозг. Здесь находится дыхательный центр, который перерабатывает разнообразные афферентные импульсы, идущие от дыхательного аппарата, а также от основных рефлексогенных сосудистых зон. Этот уровень регуляции обеспечивает ритмичную смену фаз дыхания и активность спинномозговых мотонейронов, аксоны которых иннервируют дыхательную мускулатуру.
Третий уровень регуляции – верхние отделы головного мозга, включающие и корковые нейроны. Только при участии коры большого мозга возможно адекватное приспособление реакций системы органов дыхания к изменяющимся условиям окружающей среды.
ЗАДАНИЯ ДЛЯ САМОКОНТРОЛЯ ЗНАНИЙ
ВОПРОСЫ ДЛЯ САМОПОДГОТОВКИ:
1. Общий план строения и значение дыхательной системы.
2. Строение полости носа.
3. Строение гортани.
4. Строение трахеи и главных бронхов.
5. Строение легких.
6. Плевральные полости и синусы.
7. Границы легких и плевры
8. Понятие о средостении. Органы средостения.
9. Этапы процесса дыхания.
10. Аппарат внешнего дыхания.
11. Дыхательный цикл.
12. Механизмы вдоха и выдоха.
13. Понятие о легочных объемах и легочной вентиляции.
14. Состав атмосферного, альвеолярного и выдыхаемого воздуха.
15. Газообмен в легких.
16. Транспорт газов кровью.
17. Газообмен между кровью и тканями.
18. Дыхательный центр. Регуляция дыхания.
Задание № 1. Выберите одно правильное утверждение:
1. В дыхательную систему не входит:
2. Вход в гортань при глотании закрывается:
A) голосовыми связками
B) щитовидным хрящом
D) язычком мягкого неба
3. Наибольшая частота дыхательных движений будет при концентрации углекислого газа в крови:
4. Дыхательный объем легких равен:
A) 1500 – 2000 мл
C) 1000 – 1500 мл
D) 3000 – 3500 мл
5. Обонятельной областью является:
A) верхний носовой ход
B) средний носовой ход
C) нижний носовой ход
D) преддверие носа
6. Самый крупный хрящ гортани:
7. Бифуркация трахеи находится на уровне:
A) VIII грудного позвонка
B) III грудного позвонка
C) V грудного позвонка
D) II грудного позвонка
8. Количество сегментов верхней доли правого легкого:
9. Каждая легочная долька содержит:
10. Увеличение глубины дыхания называется:
Задание №2. Ответьте на вопросы ситуационных задач
№1. Жизненная емкость обследуемого 3600 мл, резервный объем выдоха 1500 мл, резервный объем вдоха 1600 мл, частота дыхания 16 в минуту. Каков минутный объем дыхания обследуемого?
№2. Два спортсмена, близких по возрасту и физическому развитию, участвуют в забеге на 1000 метров. В конце дистанции минутный объем дыхания первого - 120 литров, частота дыхательных движений – 80 в минуту, у второго минутный объем дыхания - 120 литров, при частоте дыхательных движений – 40 в минуту. Кто из обследуемых спортсменов более тренирован? Вычислить дыхательный объем обоих спортсменов.
№3. При исследовании функционального состояния аппарата внешнего дыхания у студента выявлено: дыхательный объем – 600 мл, резервный объем вдоха – 1800 мл, резервный объем выдоха – 1900 мл. Какова жизненная емкость легких студента?
Задание №1: 1-A, 2-C, 3-D, 4-B, 5-A, 6-B, 7-C, 8-D, 9-A, 10-B.
Задание №2. №1: Минутный объем обследуемого 8000 мл, если дыхательный объем – 500 мл.
№2: Второй спортсмен тренирован в большей степени: он имеет больший дыхательный объем, поэтому частота дыхательных движений у него вдвое реже. Дыхательный объем первого спортсмена – 1,5 литра; второго спортсмена – 3 литра. №3: Жизненная емкость легких – 4300 мл.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Теория по нормальной физиологии. Тема: Регуляция дыхания. Дыхательный центр, опыты Фредерика и Холдена, действие карбогена, хеморецепторы, механорецепторы..
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Регуляция дыхания осуществляется ЦНС.
Спокойное дыхание взрослого человека характеризуется последовательной сменой актов вдоха и выдоха с частотой 14-16 дыханий в минуту.
Чтобы произошел вдох, необходимо сокращение дыхательных мышц.
Импульсы к ним поступают от мотонейронов передних рогов спинного мозга.
К диафрагме – от III – IV шейных сегментов, к межреберным мышцам – от грудных сегментов спинного мозга.
Мотонейроны получают импульсы от дыхательного центра , расположенного в продолговатом мозге.
Если перерезать спинной мозг под продолговатым мозгом , то дыхание – прекращается .
Дыхательный центр
Был открыт в 1812 г. Легаллуа и в 1842 г. Флурансом, которые своими опытами доказали его локализацию в продолговатом мозге .
Н.А.Миславский в 1885 г. уточнил местоположение дыхательного центра – в области РФ (ретикулярной формации) продолговатого мозга , т.к. перерезка между грудными и шейными сегментами спинного мозга – сохраняет диафрагмальное дыхание:
- продолговатым и спинным – полностью прекращает дыхание ,
- выше продолговатого мозга – дыхание сохраняется .
Н.А.Миславским также было показано, что дыхательный центр состоит из двух отделов:
- инспираторного (отвечает на вдох),
- экспираторного (отвечает за выдох).
Они находятся в реципрокных (то есть в противоположных) отношениях.
В настоящее время установлено:
- в РФ продолговатого мозга находятся инспираторные и экспираторные нейроны ,
- эти нейроны рассеяны диффузно , среди других нейронов РФ и относительно друг друга,
- инспираторных нейронов примерно вдвое больше , чем экспираторных.
В продолговатом мозге есть еще два скопления дыхательных нейронов – дорсальные и вентральные дыхательные ядра.
Дорсальное ядро содержит, в основном, инспираторные нейроны, контролирующие сокращение диафрагмы .
В вентральном ядре содержатся как инспираторные , так и экспираторные нейроны, которые включаются при форсированном дыхании.
Все дыхательные нейроны делят на 6 групп:
- ранние инспираторные – активны в начале фазы вдоха (инспирации),
- поздние инспираторные – активны в конце вдоха,
- полные инспираторные – активны в течение всего вдоха,
- постинспираторные – максимальный разряд в начале выдоха,
- экспираторные – активны во вторую фазу выдоха,
- преинспираторные – активны перед вдохом, они включают активную экспирацию (выдох.
Значение дыхательных нейронов:
- Преинспираторные и ранние инспираторные влияют на момент начала вдоха.
- Инспираторные нейроны ДЦ генерируют дыхательный ритм (частоту и глубину дыхания), иннервируют мотонейроны.
- Постинспираторные нейроны контролируют процесс пассивного выдоха.
- Экспираторные нейроны отвечают за активный выдох, так как иннервируют мотонейроны внутренних межреберных мышц и мышц передней брюшной стенки.
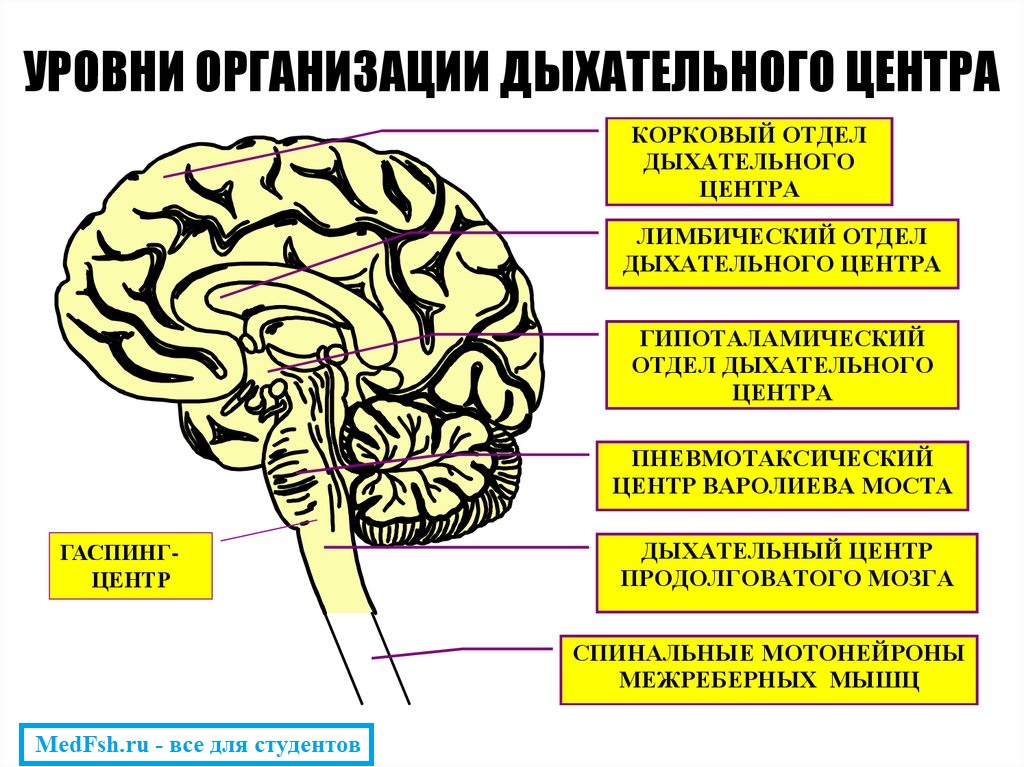
Дыхательный центр – это совокупность нервных клеток, расположенных в различных отделах ЦНС и принимающих участие в регуляции дыхания .
У двух собак, находящихся под наркозом, перекрестно соединяли сонные артерии и яремные вены, латерально же расположенные сосуды пережимали. При этом голова 1-й собаки снабжались кровью из туловища 2-й и наоборот.
У первой собаки кратковременно пережимали трахею, и у нее в крови уменьшалось содержание кислорода (гипоксемия) и увеличивалось содержание углекислого газа (гиперкапния). Эта кровь поступала в голову 2-й собаки, и у нее наступала одышка (диспноэ).
В результате у нее в крови увеличивалось содержание кислорода (гипероксимия) и уменьшалось содержание углекислого газа (гипокапния), и эта кровь поступала в голову 1-й собаки, и у нее наступало апноэ – остановка дыхания. (Нормальное дыхание — эйпноэ).
На состояние ДЦ влияет газовый состав крови:
При увеличении в крови напряжения углекислого газа и уменьшении кислорода, ДЦ – возбуждается и, наоборот, уменьшается , если в крови уменьшается напряжение углекислого газа и увеличивается напряжение кислорода.
При дыхании в герметичной камере в воздухе увеличивалось содержание CO2 и уменьшалось O2 – наступала одышка . Когда CO2 поглощается натронной известью, одышка наступала намного позже, хотя содержание O2 в воздухе значительно снижалось .
Одышка наступает:
- При снижении O2 в атмосферном воздухе с 20,94% до 12%, т.е. на 9%.
- При повышении содержания CO2 в альвеолярном воздухе на 0,17% вентиляция удваивается.
Главный активатор ДЦ – CO2
Действие карбогена
Карбоген – газовая смесь, состоящая из 96% — O2, 4% — CO2.
В сравнении с воздухом в карбогене в 4,8 раза больше O2 и в 130 раз – CO2.
Карбоген применяют при расстройствах дыхания.
Эффект карбогена связан с эффектами содержащегося в нем CO2:
- стимуляция ДЦ,
- расширение бронхов и кровеносных сосудов,
- сдвиг кривой диссоциации HbO2 вправо –> увеличение диффузии O2 из крови в ткани.
Карбоген применяется в медицинских целях для лечения горной болезни, отравления угарным газом, глаукомы, стресса, при восстановлении слуха после воздействия шума и в ряде случаев для улучшения кровоснабжения опухолей при химио- и лекарственной терапии.
В 1911 г. Винтерштейн доказал, что возбудителем ДЦ также являются ионы Н .
Ацидоз – усиливает легочную вентиляцию.
Он показал, что возбуждают ДЦ нелетучие кислоты – молочная, никотиновая и другие.
Самым сильным стимулятором дыхания являются:
- pCO2 (гиперкапния),
- pH (ацидоз),
- pO2 (гипоксемия).
Механизм действия гуморальных факторов (CO2, O2, H) на ДЦ :
- Через хеморецепторы (периферические) сосудистых рефлексогенных зон.
- Через хеморецепторы, находящиеся в продолговатом мозге (медуллярные).
- расположены в каротидных и в аортальных тельцах,
- реагируют на (в артериальной крови):
- увеличение pCO2,
- уменьшение pO2,
- увеличение H (от есть уменьшение pH).
Хеморецепторы возбуждаются постоянно CO2 и O2 , растворенными в крови, а также H, т.к. порог для pCO2 равен 20-30 мм.рт.ст. В норме pCO2 = 40 мм.рт.ст.
Порог для pO2 равен 130-140 мм.рт.ст. В норме pO2 = 100 мм.рт.ст. Одышка же наступает при pO2 ниже 50-60 мм.рт.ст.
Таким образом, хеморецепторы постоянно посылают импульсы в ДЦ, возбуждая инспираторные нейроны, причем большую роль играют хеморецепторы каротидного синуса.
- Находятся на вентролатеральной поверхности продолговатого мозга .
- Реагируют только на H и изменение напряжения CO2.
- Эти рецепторы возбуждаются позднее, поскольку требуется время для проникновения CO2 через гематоэнцефалический барьер.
- Импульсы, поступающие с медуллярных хеморецепторов в ДЦ, увеличивают прирост вентиляции на 60-80% .
Находятся в:
- легких,
- дыхательных путях,
- дыхательных мышцах (проприорецепторы)
Это барорецепторы рефлексогенных зон.
Механорецепторы легких (РРЛ)
Рецепторы растяжения легких (РРЛ) – являются наиболее значимым среди всех механорецепторов.
В 1868 г. Геринг и Брейер доказали наличие в легких рецепторов, которые возбуждаются при их растяжении, то есть при вдохе. Они являются чувствительными окончаниями блуждающих нервов, которые направляют свои импульсы в ДЦ.
Геринг и Брейтер раздували легкие и наблюдали прекражение вдоха (инспираторно-тормозной рефлекс).
Таким образом, этот рефлекс способствует смене вдоха на выдох. Он называется рефлексом Геринга-Брейера и является рефлексом саморегуляции дыхания.
При перерезке блуждающих нервов, дыхание становится редким и глубоким, альвеолы расширяются до максимального предела, т.к. вдох не тормозится. В этом случае, смене вдоха на выдох будет способствовать пневмотаксический центр (ПТЦ).
В настоящее время известно, что в легких существует 3 разновидности механорецепторов :
- РРЛ медленноадаптирующиеся,
- РРЛ быстроадаптирующиеся или ирритантные,
- Юкстаальвеолярные рецепторы капилляров.
РРЛ медленноадаптирующиеся:
- Они расположены в ГМК (гладкомышечных клетках) дыхательных путей.
- Возбуждаются при вдохе.
- С них осуществляется рефлекс Геринга-Брейера.
Ирритантные рецепторы (быстроадаптирующиеся):
- Расположены в слизистой дыхательных путей.
- Реагируют на механические и химические стимулы.
- Быстро адаптирующиеся.
- Длительное раздражение этих рецепторов приводит к хроническому бронхиту.
Физиологическое значение при вдыхании токсических веществ: Сужение бронхов -> вентиляция альвеол -> поступление этих веществ в альвеолы и кровь.
- расположены в паренхиме легких, в альвеолярных перегородках, прилегающих к капиллярам,
- стимулируются, главным образом, растяжением легочных сосудов,
- быстро реагируют на введение химических веществ в легочные сосуды,
- стимуляция может вызвать апноэ, затем учащение дыхания, уменьшение давления, брадикардию и бронхоспазм.
Возбуждение механорецепторов верхних ДП вызывает возникновение дыхательных рефлексов.
Защитные дыхательные рефлексы:
- Чихание – с рецепторов слизистой носа.
- Кашель – с ирритантных рецепторов слизистой гортани, трахеи, бронхов.
- Рефлекс ныряльщиков – остановка дыхания при действии воды на носовые ходы.
- Остановка дыхания во время акта глотания .
- Рефлексогенная задержка дыхания – сужение голосовой щели, бронхоконстрикция при вдыхании дыма, газов, едких веществ.
Механорецепторы межреберных мышц и диафрагмы (проприорецепторы) – они осуществляют обратную связь дыхательных мышц с ДЦ.
Барорецепторы рефлексогенных зон – возбуждаются они при увеличении АД и дыхание при этом угнетается. При уменьшении АД – дыхание усиливается.
Ритмическая смена дыхательных фаз
Хеморецепторы возбуждаются постоянно и постоянно посылают импульсы в продолговатый мозг, возбуждая α-инспираторные нейроны. Они возбуждаются и посылают импульсы к мотонейронам спинного мозга.
Мотонейроны возбуждаются и посылают импульсы к мышцам. Они сокращаются и наступает вдох .
При вдохе объем легких увеличивается и возбуждаются РРЛ, которые посылают возбуждающие импульсы по чувствительным волокнам блуждающего нерва к β-инспираторным нейронам. В результате суммации импульсов от α-инспираторных нейронов и рецепторов растяжения легких достигается порог и возбуждаются и β-инспираторные нейроны благодаря влиянию вышележащих отделов ЦНС.
Инспираторно-тормозные нейроны посылают тормозные импульсы к α-инспираторным нейронам. В результате α-инспираторные нейроны тормозятся и не посылают импульсы к мотонейронам. Мышцы расслабляются, происходит выдох .
К β-инспираторным нейронам не поступают импульсы, и они тормозятся (не возбуждаются).
β-инспираторные нейроны не возбуждают инспираторно-тормозные нейроны и поэтому они не посылают импульсы к α-инспираторным нейронам. α-инспираторные нейроны вновь возбуждаются импульсами от хеморецепторов и наступает вдох .
Читайте также:
