
Возбуждение от нервного волокна к мышечному волокну
Передача возбуждения с нервного волокна на мышечное осуществляется через посредство нервно-мышечного синапса (морфо-функциональный контакт между пресинаптическим окончанием аксона эфферентного нейрона и определенным участком мембраны мышечного волокна). По способу передачи информации мионевральный синапс относится к синапсам химического типа.
В нервно-мышечном синапсе, как и в любом другом, выделяют:
Ø пресинаптическую мембрану (часть мембраны пресинаптического окончания аксона двигательного нейрона, вступающая в контакт с мембраной мышечного волокна),
Ø постсинаптическую мембрану (часть мембраны мышечного волокна, с которой вступает в контакт мембрана нервного волокна),
Ø синаптическую щель (узкое пространство между пре- и постсинаптической мембранами, заполненное межклеточной жидкостью; ширина этого пространства в нервно-мышечном синапсе составляет 80-100 нм).
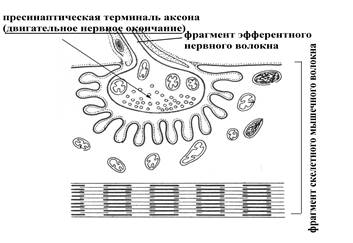

Рис. 8. Мотонейрон (двигательный нейрон) и иннервируемое им скелетное мышечное волокно (А); прямоугольник указывает на нервно-мышечный синапс (Б – его схематическое представление), в образовании которого принимает участие пресинаптическая терминаль аксона мотонейрона (по Е.А. Шубниковой, 1987).
Нервно-мышечный синапс является химическим: передача возбуждения с нервного волокна на мышечное (будь-то скелетное или гладкое мышечное волокно) осуществляется с участием специальных химических посредников – медиаторов (химических веществ, чаще всего, органической природы). Так, медиатором в синапсе между нервным и скелетным мышечным волокнами является ацетилхолин (Ах), а в синапсах между нервными и гладкими мышечными волокнами – могут выступать как ацетилхолин, так и норадреналин (НА).
Рассмотрим механизм передачи возбуждения с нервного волокна на скелетное мышечное волокно (этот механизм является в определенном плане универсальным для всех химических синапсов, но в зависимости от того, какие структуры принимают участие в образовании синапса, имеет свои особенности).
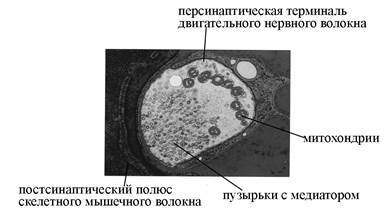
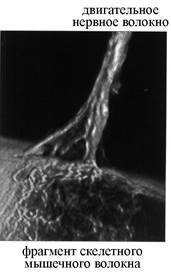
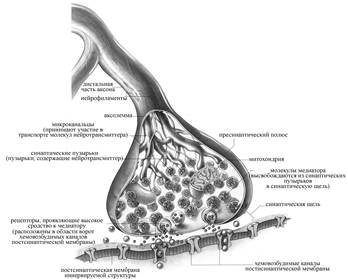
Рис. 6. Ультраструктурная организация нервно-мышечного синапса (по T. Smit, 2002). А – СЭМ терминали двигательного нервного волокна и фрагмента скелетного мышечного волокна, принимающего участие в образовании синаптического контакта. Б – ТЭМ нервно-мышечного синапса. В – схема строения химического синапса.
Возбуждение в виде нервного импульса распространяется от тела нервной клетки вдоль ее аксона к пресинаптическому окончанию аксона. При этом на пресинаптической мембрана аксона возникает не потенциал действия, а локальная деполяризация. В пресинаптической мембране, наряду с электровозбудимыми Na + - и К + -каналами, имеются и электровозбудимые (потенциалзависимые) Са 2+ -каналы, срабатывающие на деполяризацию открытием своих единственных ворот. Как следствие этого, проницаемость пресинаптической мембраны для ионов кальция повышается, и они начинают входить по концентрационному градиенту из синаптической щели в пресинаптическое окончание аксона, в результате чего концентрация кальция внутри пресинаптического окончания возрастает. В пресинаптическом окончании аксона имеются актиновые филаменты, которые ассоциированы с мембранными пузырьками, содержащими медиатор (ацетилхолин), и миозиновые филаменты, которые одним своим концом прикреплены к внутренней поверхности пресинаптической мембраны, а другой их конец свободен и направлен в сторону пузырьков с медиатором. Повышение концентрации кальция в пресинаптическом окончании, в свою очередь, делает возможным взаимодействие актиновых и миозиновых филаментов между собой, что сопровождается продвижением пузырьков с медиатором в направлении внутренней поверхности пресинаптической мембраны.
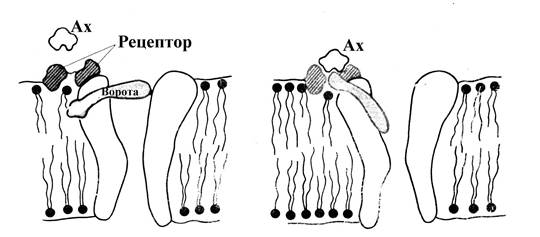
Рис. 7. Схематическое изображение хемовозбудимого канала . А – закрытое состояние, в отсутствии связи ацетилхолина (Ах) с рецептором, Б – открытое состояние канала, возникающее после присоединения ацетилхолина к холинорецептору.
В момент, когда пузырьки, содержащие ацетилхолин, приближаются к пресинаптической мембране на определенное расстояние, их мембрана встраивается в пресинаптическую, а содержимое (медиатор – ацетилхолин) оказывается в синаптической щели. Количество молекул ацетилхолина во всех пузырьках примерно одинаково; содержимое одного пузырька получило название кванта медиатора. Таким образом, деполяризация пресинаптической мембраны сопровождается выделением в синаптическую щель определенного количества квантов медиатора ацетилхолина.
Ацетилхолин диффундирует через синаптическую щель и достигает постсинаптической мембраны. Особенностью постсинаптической мембраны является отсутствие потенциалзависимых каналов (имеющихся во всей остальной мембране мышечного волокна, называемой внесинаптической), но наличие хемовозбудимых каналов. Состояние хемовозбудимых каналов не зависит от величины мембранного потенциала, но может изменяться под влиянием определенных химических веществ (в частности, молекул медиатора).
В хемовозбудимом канале, подобно электровозбудимому, различают отверстие (пору), в области которого расположены ворота (белковой природы); но состояние этих ворот (их пространственная структура, определяющая открытое или закрытое состояние канала) зависит не от величины заряда на мембране, а от состояния, расположенного вблизи этих ворот белка-рецептора, обладающего высоким сродством к медиатору (и возможно некоторым другим веществам). В частности, в хемовозбудимых каналах постсинаптической мембраны скелетного мышечного волокна имеются белковые холинорецептор и ворота. В отсутствии ацетилхолина в области холинорецептора канал закрыт. Присоединение ацетилхолина к холинорецептору сопровождается изменением его конформации, что приводит и к изменению конформации расположенных поблизости от холинорецептора ворот таким образом, что они отодвигаются от поры канала, и канал открывается. Размер пор в хемовозбудимых каналах (
0,65 нм) несколько больше такового в электровозбудимых натриевых и калиевых каналах, в связи с чем они потенциально способны пропускать любые ионы (натрия, калия, кальция), т.е. характеризуются сравнительно низкой селективностью. Вместе с тем преобладающими в силу распределения электрического заряда на мембране является входящий натриевый ток, который деполяризует постсинаптическую мембрану мышечного волокна (возникает т.н. постсинаптический потенциал). Но такая деполяризация не может приобретать самообновляющийся характер, поскольку в постсинаптической мембране отсутствуют потенциалзависимые каналы. Доля открытых хемовозбудимых каналов определяется количеством высвободившихся в синаптическую щель молекул медиатора. В связи с этим постсинаптический потенциал, в отличие от потенциала действия, является градуальным. При некоторой определенной амплитуде постсинаптического потенциала в ближайшем к синапсу участке внесинаптической мембраны мышечного волокнаактивная деполяризация, вызванная этим постсинаптическим потенциалом, приобретает самообновляющийся характер, что приводит к генерации нервного импульса, который проводится вдоль скелетного мышечного волокна, инициируя его сокращение.
Важную роль в функционировании нервно-мышечного синапса играет фермент холинэстераза, который локализован в синаптической щели и расщепляет ацетилхолин. Расщепление ацетилхолина обеспечивает не постоянное, а временное его действие на постсинаптическую мембрану, благодаря чему синаптическая передача носит дискретный характер. При ритмическом слишком частом следовании нервных импульсов к пресинаптическому окончанию по нервному волокну холинэстераза не успевает расщепить ацетилхолин, выделившейся при каждом предыдущем возбуждении пресинаптической мембраны нервного волокна, и все бóльшая доля хемовозбудимых каналов постсинаптической мембраны скелетного мышечного волокна оказывается открытой, что приводит к росту постсинаптического потенциала. Данное обстоятельство является причиной длительной и стойкой деполяризации внесинаптической мембраны мышечного волокна, сопровождающейся постепенной натриевой инактивацией и снижением возбудимости внесинаптической мембраны вплоть до полной временной утраты способности генерировать нервные импульсы; наступает блок синаптической передачи (пессимальное торможение нервно-мышечного синапса). Пессимальное торможение нервно-мышечной передачи наступает при частоте стимуляции нервного волокна 100-150 имп./с. Таким образом, лабильность нервно-мышечного синапса в связи с химическим механизмом передачи гораздо меньше таковой нервного волокна (до 2000 имп./с) и скелетных мышечных волокон (до 500 имп./c).
Химический механизм передачи возбуждения через синапс определяет следующие ее особенности:
Ø одностороннее проведение возбуждения (только в направлении от нервного волокна на мышечное волокно). Связано с тем, что синаптическая щель в химических синапсах довольно широкая (до 80-100 нм), и ионные токи, возникающие при деполяризации постсинаптической мембраны, шунтируются в этой щели и не способны вызвать возвратную деполяризацию пресинаптической мембраны
Ø задержанное проведение возбуждения (латентный период передачи информации через синапс составляет 0,3-0,5 мс). Обусловлено тем, что для осуществления процессов выделения медиатора в синаптическую щель под действием приходящего к пресинапсу нервного импульса, диффузии медиатора к постсинаптической мембране и проявления его эффектов на постсинаптическую мембрану требуется определенное время
Ø относительно низкая лабильность синапсов (по сравнению с нервными и мышечными волокнами) в связи с зависимостью процесса нервно-мышечной передачи от медиатора и необходимостью для нормальной работы синапса постоянного расщепления медиатора после каждого проводимого возбуждения
Ø относительно высокая утомляемость синапсов (поскольку максимальная продолжительность их постоянного функционирования лимитируется запасом "готового" к высвобождению медиатора; кроме того, при длительном проведении возбуждения через синапс уменьшается чувствительность постсинаптической мембраны к ацетилхолину, что также является одной из причин развития утомления в синапсе).
Химический способ передачи информации через нервно-мышечный синапс обуславливает, наряду с отмеченным, и высокую степень зависимости этого процесса от ряда химических веществ (т.н. модуляторов синаптической передачи). Так, временно заблокировать нервно-мышечную передачу можно несколькими путями:
Ø блокируя секрецию ацетилхолина в синаптическую щель (таким действием обладают столбнячный и ботулинический токсины),
Ø блокируя холинорецепторы постсинаптической мембраны (таким действием обладают кураре и его производные),
Ø инактивируя холинэстеразу (например, под действием прозерина).
Механизм распространение возбуждения по нервным и мышечным волокнам осуществляется при помощи местных токов, возникающих между возбужденными и невозбужденными участками мембраны. Теория проведения возбуждения при помощи местных токов была сформулирована Германом в 1899 году и получила в настоящее время экспериментальное подтверждение.
На рис.15 стрелками показано направление тока, возникающего между возбужденным (А) и соседним невозбужденным (Б) участками.
Распространение местных токов по длине волокна определяется ее кабельными свойствами. Направление же местного тока таково, что он деполяризует соседний с активным (А) покоящийся (Б) участок мембраны. В итоге в участке Б уменьшается мембранный потенциал, т.е. происходит деполяризация, которая при достижении критического уровня вызывает возникновение ПД, и теперь уже возбужденный участок Б становится способным активировать соседний с ним покоящийся участок.
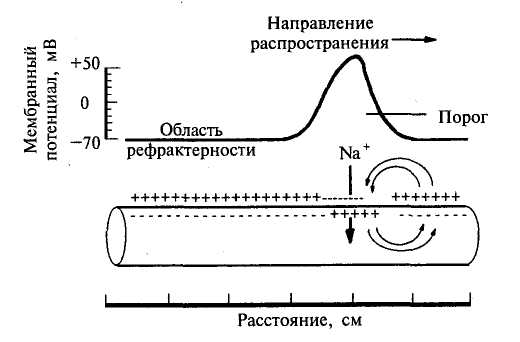
Рис. 15. Механизм распространения возбуждения с помощью местных токов после прохождения потенциала действия в нервном волокне возникает зона невозбудимости, или реф-рактерности.
В нервных мякотных волокнах, покрытых миелиновой оболочкой, ПД распространяется очень быстро, вследствие того, что возбуждение возникает только в перехватах Ранвье.
К миелинизированным волокнам относятся чувствительные и двигательные волокна, а также часть волокон вегетативной нервной системы.
На рис.16 показано строение миелинизированного нервного волокна. Миелиновая оболочка создается Шваннновской клеткой, многократно обертывающей осевой цилиндр волокна, поверхность которого образована плазматической мембраной. Свободные от миелиновой оболочки участки мембраны шириной, примерно, 1мкм называют перехватами Ранвье. В нервных волокнах, диаметром 10-20 мкм, длина межперехватных участков составляет 1-2 мм. Миелиновая оболочка участвует в регуляции обмена веществ и росте осевого цилиндра, выполняет функцию электрического изолятора, благодаря высокому сопротивлению и обеспечивает более экономное и быстрое проведение возбуждения.


Рис. 4.1. Непрерывное распространение ПД в нервном волокне:
1 — возникновение ПД в нервном волокне и распространение ПД в обе
стороны от места возникновения. В области деполяризации мембраны (1)
преобладает входящий в клетку натриевый ток; 2 — соседняя область,
в которой локальный ток от области ПД вызывает деполяризацию
до критического уровня
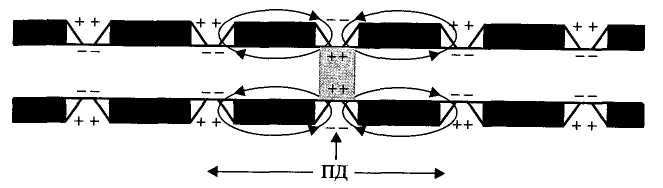
Рис. 4.2. Сальтаторное распространение ПД в миелиновых нервных волокнах. Возникновение ПД в перехвате Ранвье среднего участка волокна и распространение ПД в обе стороны от места возникновения. Стрелками показаны токи на электротоническом этапе распространения ПД. В области перехватов Na+ движется в клетку, К+ — из клетки

Рис. 3.17. Схемы распространения ПД по нервным клеткам: а — общая схема; б — распространение ПД по неми-елинизированному (безмякотному) волокну (1, 2, 3, 4 — участки мембраны нейрона); в — распространение ПД по миелинизированному волокну
Рис. 16. Строение и распространение возбуждения по миелинизированному волокну.
А. Образование миелинового чехла на аксоне.
1 — наматывание слоев миелина; 2 — аксон; 3 — олигодендроцит; 4 — перехват Ранвье; 5 — слой миелиновой оболочки.
Б. Миелиновое нервное волокно.
В. Распространение возбуждения
Участки между перехватами, покрытые миелином являются практически невозбудимыми. Перепрыгивание нервного импульса с одного перехвата на другой называют сальтаторное проведение возбуждения. Оно связано с высокой плотностью натриевых каналов в перехватах (до 1000 на 1 мкм мембраны). Сальтаторное проведение возбуждения от одного перехвата до другого оказывается возможным только потому, что амплитуда ПД в каждом перехвате в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата Ранвье.
Предположение о скачкообразном распространении возбуждения в нервных волокнах впервые было высказано Вериго в 1899 году. Возбуждение по мякотному нервному волокну проводится с большой скоростью без затухания (декремента), а интенсивность его одинакова в любом участке волокна. Расход же энергии при проведении возбуждения по миелинизированным волокнам невелик
Рис.17. Проведенеие возбуждения по миелинизированным нервным волокнам.
В мышечных и безмякотных волокнах возбуждение осуществляется непрерывно от точки к точке. Такое распространение возбуждения происходит с затуханием.
Безмякотные волокна у позвоночных животных принадлежат в основном симпатической нервной системе. Они не имеют миелиновой оболочки, однако изолированы друг от друга Шванновскими клетками.
Нервы обычно состоят из миелинизированных и немиелинизированных волокон в различном соотношении. В нервах иннервирующих скелетные мышцы преобладают миелинизированные волокна, а в нервах вегетативной нервной системы, например, в блуждающем нерве количество безмякотных волокон достигает 80-95%.
Современная техника исследования позволяет точно определить скорость проведения возбуждения путем регистрации с помощью осциллографа латентного периода возникновения ПД при раздражении нерва с разных участков. Скорость проведения возбуждения зависит от диаметра волокна и структуры его мембраны - чем толще волокно, тем больше скорость проведения возбуждения в нем. В настоящее время нервные волокна по скорости проведения возбуждения, длительности различных фаз потенциала действия и строению принято подразделять на три основных типа, обозначаемых буквами А, Б и С.
Таблица 2. Свойства различных нервных волокон теплокровных
Рис. 30. Сокращения икроножной мышцы при разной частоте раздражения (по Е. Е. Жукову).
Одиночное сокращение показано пунктиром. Цифры над кривыми — частоты стимулов. При частотах 20—35 в секунду — зубчатый тетанус, при частоте 1 15 в секунду — гладкий тетанус.
Таблица 3 Свойства различных нервных волокон теплокровных
| Длитель- | Длитель- | Длитель- | ||||
| Тип | Диаметр | Скорость | ность пика | ность | следовой | |
| волокон | волокна, | проведе- | потенциала | следовой | гиперпо- | - Функция |
| мкм | ния, м/с | действия, ме | деполяризации, мс | ляризации, мс | ||
| А а | 12—22 | 70—120 | 0,4—0,5 | 15—20 | 40—60 | Моторные волокна скелет- |
| ных мышц, афферентные | ||||||
| i | волокна от мышечных ре- | |||||
| цепторов | ||||||
| 8—12 | 40—70 | 0,4—0,6 | — | — | Афферентные волокна от ре- | |
| цепторов прикосновения | ||||||
| А7 | 4—8 | 15—40 | 0,5—0,7 | — | — | Афферентные волокна от ре- |
| цепторов прикосновения и | ||||||
| ■ | Давления, эфферентные | |||||
| волокна к мышечным ве- | ||||||
| Ретенам | ||||||
| Аб | 1—4 ' | 5—15 | 0,6—1,0 | — | — | Афферентные волокна от не- |
| которых рецепторов тепла, | ||||||
| Давления, боли | ||||||
| В | 1—3,5 | 3—18 | 1—2 | Отсутст- | 100—300 | Преганглионарные вегетати- |
| вует | вные волокна | |||||
| С | 0,5—2,0 | 0,5—3 | 2,0 | 50—80 | 300-^1000 | Постганглионарные вегета- |
| тивные волокна, афферен- | ||||||
| тные волокна от некоторых | ||||||
| рецепторов те,пла, давле- | ||||||
| * | ния, боли ( |
Для сохранения функции нервного волокна необходима его связь с телом. При перерезке нервного волокна его периферический конец, отделенный от тела клетки перерождается и дегенерирует (у теплокровных животных через 2 - 3 дня), миелиновая оболочка скапливается в виде капель жира и рассасывается. При этом нарушается функция иннервируемого органа. Центральная же часть нервного волокна способна к регенерации. Регенерация нерва происходит очень медленно.
Лабильность. Нервные волокна обладают разной лабильностью (функциональной подвижностью). Показателем лабильности является максимальное число потенциалов действия, которое способно возбудимое образование генерировать в 1с в соответствии с ритмом раздражения. Она определяется скоростями процессов изменений ионной проводимости, лежащих в основе абсолютной и относительной рефрактерности. При увеличении частоты ритмического раздражения лабильность ткани повышается, т.е. ткань отвечает более высокой частотой возбуждения по сравнению с исходной частотой. Это явление было открыто в 1923 году А.А. Ухтомским и получило название усвоение ритма раздражения.
Лабильность нервных волокон колеблется от нескольких десятков до 500 импульсов в секунду, а в ряде случаев до 1000 и более. Наиболее высокой лабильностью обладают толстые миелинизированные волокна. Лабильность мышцы - около 200имп/сек, нервно-мышечного синапса- порядка 100 имп/сек.
Существуют агенты, например, местные анестетики, способные замедлять реактивацию натриевых каналов и потому увеличивающие длительность фазы относительной рефрактерности. Так, если участок нервного волокна подвергнуть воздействию малых концентраций новокаина (или какого – либо другого местного анестетика), то волокно утрачивает способность проводить высокочастотные разряды импульсов, тогда как низкочастотные разряды еще продолжают проходить.
Сходным образом влияет на процесс реактивации повышение концентрации ионов K+в окружающей нервные волокна жидкости.
Обнаружены агенты, снижающие возбудимость (в результате блокады натриевых каналов), но не влияющие на длительность рефрактерной фазы. К числу таких агентов относится специфический блокатор натриевых каналов - тетродотоксин.
Таким образом, различные по характеру своего воздействия агенты способны вызывать внешне сходные изменения возбудимости и проведения нервных импульсов.
Обмен веществ в нервном волокне относительно невелик. О динамике обмена веществ нерва в покое и при возбуждении можно судить по его теплопродукции, которую впервые удалось зарегистрировать Хиллу в 1926 г.
Деятельное состояние нерва характеризуется увеличением выделения тепла - повышением теплопродукции. Различают фазу начального теплообразования, длящуюся при одиночном раздражении в течение десятых долей секунды и фазу запаздывающего теплообразования, продолжающуюся в течение минуты. Исследования при помощи высокочувствительных приборов показали, что начальное теплообразование сопровождает процесс возбуждения (генерацию ПД), а запаздывающее теплообразование связано с восстановительными процессами. При возбуждении нервных волокон выделяется значительно меньше тепла по сравнению с количеством энергии, выделяемой мышцей. Деятельное состояние нерва характеризуется увеличением потребления аденозинтрифосфата (АТФ) и креатинфосфата (КФ), выделением молочной кислоты, усилением белкового обмена, обмена нуклеиновых кислот и фосфолипидов.
Нервные волокна характеризуются относительной неутомляемостью, которая связана, прежде всего, с уровнем обмена веществ. Энергетические траты при возбуждении нервных волокон связаны главным образом с работой натрий - калиевого насоса.
Теория по нормальной физиологии на тему: Синапс. Физиология мышечных волокон. В данной статье рассматривается механизм синаптической передачи, НМС…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Синапс — это специфическое место контакта двух возбудимых систем (клеток) для передачи возбуждения.
По способу передачи сигналов:
- механические,
- химические,
- электрические.
По виду медиатора: холинэргические и др.
Нервно-мышечный синапс (НМС) — химический, передача с помощью медиатора ацетилхолина.
Синонимы к слову НМС:
- Нервно-мышечное соединение;
- Моторная концевая пластинка.

Аксоны нервных клеток на своих окончаниях теряют миелиновую оболочку , ветвятся, и концевые веточки аксона утолщаются. Это пресинаптическая терминаль или бляшка или пуговка , которая погружается в углубление на поверхности мышечного волокна.
Покрывающая концевую веточку аксона поверхностная мембрана называется пресинаптической мембраной , т.е. это мембрана, покрывающая поверхность синаптической бляшки (терминали аксона).
Мембрана, покрывающая мышечное волокно в области синапса, называется постсинаптической мембраной , или концевой пластинкой. Она имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна, за счет чего увеличивается площадь контакта.

На постсинаптической мембране находятся белковые структуры — рецепторы , способные связывать медиатор . В одном синапсе количество рецепторов достигает 10-20 млн.
Между пре- и постсинаптическими мембранами находится синаптическая щель , размеры ее в среднем 50 нм, она открывается в межклеточное пространство и заполнена межклеточной жидкостью .
В синаптической щели находится мукополисахаридное плотное вещество в виде полосок, мостиков и содержится фермент ацетилхолинэстераза .
В пресинаптической терминали находится большое количество пузырьков или везикул , заполненных медиатором — химическим веществом посредником, осуществляющим передачу возбуждения.
В нервно-мышечном синапсе медиатор — ацетилхолин (АХ).

АХ синтезируется из холина и уксусной кислоты (ацетил-коэнзима А) с помощью фермента холинэстеразы. Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование АХ.
3 фракции медиатора:
- Первая фракция — доступная — располагается рядом с пресинаптической мембраной.
- Вторая фракция — депонированная — располагается над первой фракцией.
- Третья фракция — диффузно рассеянная — наиболее удаленная от пресинаптической мембраны.
Механизм синаптической передачи



Ионы Ca вызывают образование специального белкового комплекса , который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома .



Излитию содержимого пузырька в щель способствует белок синаптопорин , формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.





На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает ! Он формируется на соседней мембране мышечного волокна.
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе :
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП) .
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Физиология мышечных волокон
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл , расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин .
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации . Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
Сравнительная характеристика скелетных и гладких мышц:
Читайте также:
