
Возбуждения мембраны нервного волокна
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
Нервные волокна (отростки нервных клеток) выполняют специализированную функцию - проведение нервных импульсов. По морфологическому признаку нервные волокна делятся на миелиновые (покрытые миелиновой оболочкой) и безмиелиновые. Нервные волокна формируют нерв или нервный ствол, состоящий из большого числа нервных волокон, заключенных в общую соединительноткан-ную оболочку. В состав нерва входят миелиновые и безмиелиновые волокна.
Нервные волокна, проводящие возбуждение от рецепторов в ЦНС называются афферентными, а волокна, проводящие возбуждение от ЦНС к исполнительным органам, называются эфферентными. Нервы состоят из афферентных и эфферентных волокон.
Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью.
Проведение возбуждения по нервным волокнам осуществляется по определенным законам.
Закон двустороннего проведения возбуждения по нервному волокну. Возбуждение по нервному волокну распространяется в обе стороны от места его возникновения, т. е. центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения.
Закон анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е. к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, т. е. не переходя с одного волокна на другое. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна.
Изолированное проведение возбуждения имеет важное значение. Нервсодержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки, ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно.
Нервные волокна по скорости проведения возбуждения, диаметру волок- на, длительности различных фаз потенциала действия и строению принято подразделять на три типа: А, В, С. Волокна типа А в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта.
Волокна типа А покрыты миелиновой оболочкой. Наиболее толстые из них А - альфа, они имеют диаметр 12-22 мкм и высокую скорость проведения возбуждения - 70-120 м/с. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам.
Три другие группы волокон тина А (бета, гамма, дельта) имеют меньший диаметр от 8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна этих групп относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых, рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам.
К волокнам типа В относятся миелинизированные преганглио-нарные волокна вегетативной нервной системы. Их диаметр - 1-3,5 мкм, а скорость проведения возбуждения - 3-18 м/с.
К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра - 0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более 3 м/с (0,5-3,0 м/с) . Большинство волокон типа С - это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
Механизм проведения возбуждения по нервным волокнам. Механизм распространения возбуждения у различных нервных волокон неодинаков. По современным представлениям распространение возбуждения по нервным волокнам осуществляется на основе ионных механизмов генерации потенциала действия.
При распространении возбуждения по безмиелиновому нервному волокну местные электрические токи,, которые возникают между его возбужденным участком, заряженным отрицательно, и невозбужденным, заряженным положительно, вызывают деполяризацию мембраны до критического уровня с последующей генерацией ПД в ближайшей точке невозбужденного участка мембраны. Этот процесс повторяется многократно. На всем протяжении нервного волокна происходит процесс репродукции нового ПД в каждой точке мембраны волокна. Такое проведение возбуждения называете и непрерывным.
Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки (перехватов Ранвье) создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Местные электрические токи возникают между соседними перехватами Ранвье, т. к. мембрана возбужденного перехвата становится заряженной отрицательно по отношению к поверхности соседнего невозбужденного перехвата. Эти местные токи деполярязуют мембрану невозбужденного перехвата до критического уровня и в нем возникает ПД (рис. 4). Следовательно, возбуждение как бы "перепрыгивает" через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Такой механизм распространения возбуждения называется сальтаторным или скачкообразным. Скорость такого способа проведения возбуждения значительно выше и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов.
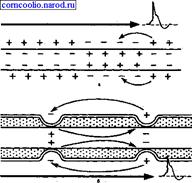
Рис. 4. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах.
"Перепрыгивание" потенциала действия через участок между перехватами возможно потому, что амплитуда ПД в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата. ПД может "перепрыгивать" не только через один, но и через два межперехватных промежутка. Это явление может наблюдаться при снижении возбудимости соседнего перехвата под действием какого-либо фармакологического вещества, например, новокаина, кокаина и др.
Нервные волокна обладают лабильностью - способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в единицу времени без трансформации ритма раздражения. Лабильность определяется длительностью пика потенциала действия, т. е. фазой абсолютной рефрактерности. Так как длительность абсолютной ре-фрактерности у спайкового потенциала нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000 импульсов в секунду.
Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности было названо Н. Е. Введенским парабиозом. В развитии состояния парабиоза можно отметить три, последовательно сменяющих друг друга, фазы: уравнительную, парадоксальную, тормозную.
В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители - больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100) , то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) - происходит уравнивание ответной реакции.
В парадоксальную-фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс - на редкие раздражители ответная реакция больше, чем на частые.
В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние.
Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ также связано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
Парабиоз - явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности.
Механизм развития парабиотического состояния сводится к следующему. При воздействии на нервное волокно парабиотического фактора нарушается способность мембраны увеличивать натриевую проницаемость в ответ на раздражение. В участке альтерации инак-тивация натриевых каналов, вызванная повреждающим агентом, суммируется с инактивацией, вызываемой нервным импульсом, и возбудимость снижается настолько, что проведение следующего импульса блокируется.
Теория по нормальной физиологии на тему: Синапс. Физиология мышечных волокон. В данной статье рассматривается механизм синаптической передачи, НМС…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Синапс — это специфическое место контакта двух возбудимых систем (клеток) для передачи возбуждения.
По способу передачи сигналов:
- механические,
- химические,
- электрические.
По виду медиатора: холинэргические и др.
Нервно-мышечный синапс (НМС) — химический, передача с помощью медиатора ацетилхолина.
Синонимы к слову НМС:
- Нервно-мышечное соединение;
- Моторная концевая пластинка.

Аксоны нервных клеток на своих окончаниях теряют миелиновую оболочку , ветвятся, и концевые веточки аксона утолщаются. Это пресинаптическая терминаль или бляшка или пуговка , которая погружается в углубление на поверхности мышечного волокна.
Покрывающая концевую веточку аксона поверхностная мембрана называется пресинаптической мембраной , т.е. это мембрана, покрывающая поверхность синаптической бляшки (терминали аксона).
Мембрана, покрывающая мышечное волокно в области синапса, называется постсинаптической мембраной , или концевой пластинкой. Она имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна, за счет чего увеличивается площадь контакта.

На постсинаптической мембране находятся белковые структуры — рецепторы , способные связывать медиатор . В одном синапсе количество рецепторов достигает 10-20 млн.
Между пре- и постсинаптическими мембранами находится синаптическая щель , размеры ее в среднем 50 нм, она открывается в межклеточное пространство и заполнена межклеточной жидкостью .
В синаптической щели находится мукополисахаридное плотное вещество в виде полосок, мостиков и содержится фермент ацетилхолинэстераза .
В пресинаптической терминали находится большое количество пузырьков или везикул , заполненных медиатором — химическим веществом посредником, осуществляющим передачу возбуждения.
В нервно-мышечном синапсе медиатор — ацетилхолин (АХ).

АХ синтезируется из холина и уксусной кислоты (ацетил-коэнзима А) с помощью фермента холинэстеразы. Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование АХ.
3 фракции медиатора:
- Первая фракция — доступная — располагается рядом с пресинаптической мембраной.
- Вторая фракция — депонированная — располагается над первой фракцией.
- Третья фракция — диффузно рассеянная — наиболее удаленная от пресинаптической мембраны.
Механизм синаптической передачи



Ионы Ca вызывают образование специального белкового комплекса , который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома .



Излитию содержимого пузырька в щель способствует белок синаптопорин , формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.





На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает ! Он формируется на соседней мембране мышечного волокна.
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе :
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП) .
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Физиология мышечных волокон
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл , расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин .
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации . Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
Сравнительная характеристика скелетных и гладких мышц:
Читайте также:
