
Зародыш ланцетника нервная трубка
Индивидуальное развитие ланцетника представляет собой простейшую исходную схему эмбриогенеза, путем постепенного усложнения которой в ходе эволюции возникли более сложные системы развития хордовых животных, включая человека.
СТРОЕНИЕ ЯЙЦЕКЛЕТКИ. ОПЛОДОТВОРЕНИЕ
Яйца ланцетника бедны желтком и микроскопически малы (100-120мкм), относятся к изолецитальному типу. Желточные зерна невелики и распределены в цитоплазме почти равномерно. Тем не менее, в яйцеклетке выделяют анимальный и вегетативный полюс. В области анимального полюса при созревании яйцеклетки происходит отделение редукционных телец. Ядро в оплодотворенной яйцеклетке находится ближе к анимальному полюсу в силу не совсем равномерного распределения желтка, располагаясь в свободной от желточных включений части клетки. Созревание яйцеклетки происходит в воде. Первое редукционное тельце отделяется на анимальном полюсе ооцита еще до оплодотворения. Оно смывается водой и погибает.
Самки ланцетника выметывают яйца в воду, сюда же самцы выпускают сперматозоиды – оплодотворение наружное, моноспермное. После проникновения сперматозоида вокруг яйцеклетки образуется оболочка оплодотворения, которая препятствует проникновению в яйцо других
избыточных спермиев. Вслед за этим происходит отделение второго редукционного тельца, которое располагается между желточной оболочкой и яйцеклеткой.
Все дальнейшее развитие проходит также в воде. Через 4-5 дней из яйцевой оболочки вылупливается микроскопическая личинка, которая переходит к самостоятельному питанию. Сначала она плавает, а затем оседает на дно, растет и совершает метаморфоз
ДРОБЛЕНИЕ. БЛАСТУЛА
Малое количество желтка объясняет простоту дробления и гаструляции. Дробление полное, почти равномерное, радиального типа, в результате образуется целобластула (рис. 1).

Рис. 1. Дробление яйца ланцетника (по Алмазову, Сутулову, 1978):
А – зигота; Б, В, Г – образование бластомеров (показано расположение веретена деления)
Анимальный полюс приблизительно соответствует будущему переднему концу тела личинки. Оплодотворенная яйцеклетка (зигота) целиком дробится на бластомеры в правильной геометрической прогрессии. Бластомеры почти одинаковой величины, анимальные лишь не-
сколько мельче вегетативных. Первая борозда дробления – меридиональная, проходит через анимальный и вегетативный полюс. Она разделяет шаровидное яйцо на две совершенно симметричные половины, но бластомеры округляются. Они шаровидные, имеют малую площадь со-
прикосновения. Вторая борозда дробления также меридиональная, перпендикулярна первой, а третья – широтная.
По мере увеличения количества бластомеров они все больше расходятся от центра зародыша, образуя посередине большую полость. В конце концов, зародыш принимает форму типичной целобластулы – пузырька со стенкой, образованной одним слоем клеток – бластодермой и с полостью, заполненной жидкостью – бластоцелем (рис. 2).
Клетки бластулы, вначале округлые и потому не плотно сомкнутые, затем приобретают форму призм и плотно смыкаются. Поэтому позднюю бластулу, в противоположность ранней, называют эпителиальной.
Стадия поздней бластулы завершает период дробления. К концу этого периода размеры клеток достигают минимума, а общая масса зародыша не увеличивается по сравнению с массой оплодотворенной яйцеклетки.


Рис. 2. Бластула ланцетника (по Алмазову, Сутулову, 1978):
А – внешний вид; Б – поперечный разрез (стрелка показывает задне-переднее направление тела будущего зародыша); В – расположение материалов будущих органов на сагитальном разрезе бластулы
Гаструляция происходит путем инвагинации – впячивания вегетативного полушария бластулы внутрь, по направлению к анимальному полюсу (рис. 3). Процесс протекает постепенно и заканчивается тем, что все вегетативное полушарие бластулы уходит внутрь и становится внутренним зародышевым листком – первичная энтодерма зародыша. Фактор, обуславливающий инвагинацию – разница темпов деления клеток в краевой зоне и в вегетативной части бластулы, приводящей к активному перемещению клеточного материала. Анимальное полушарие становится
.jpg)
Рис. 3. Начальные стадии гаструляции ланцетника (по Мануиловой, 1973):
наружным зародышевым листком – первичная эктодерма. Зародыш приобретает вид двуслойной чаши с широко зияющим отверстием – первичным ртом или бластопором. Полость, в которую ведет бластопор, называют гастроцель (полость первичного кишечника). Бластоцель в результате впячивания низводится до узкой щели между наружным и внутренним зародышевыми листками. На данной стадии зародыш носит название гаструлы (рис. 4 А, Б).
Первичный кишечник (архентерон), представленный внутренним зародышевым листком, окружающим полость гаструлы, является зачатком не только пищеварительной системы, но и других органов и тканей личинки. Бластула, как и яйцеклетка, плавает анимальным полюсом кверху в силу большего веса вегетативного полушария.
В результате инвагинации центр тяжести зародыша перемещается и гаструла поворачивается бластопором кверху.
Бластопор окружен дорсальной, вентральной и боковыми губами. Далее происходит концентрическое смыкание краев бластопора и удлинение зародыша. У ланцетника, представителя вторичноротых, бластопор соответствует не ротовому, а заднепроходному отверстию, обозначая
задний конец зародыша. В результате смыкания краев бластопора и выпячивания тела в передне-заднем направлении, зародыш удлиняется. При этом поперечник гаструлы уменьшается – общая масса составляющих зародыш клеток не может увеличиваться, пока развитие идет под покровом яйцевых оболочек. Зародыш приобретает билатеральную симметрию.
Расположение зачатков в поздней гаструле лучше всего видно на поперечном разрезе зародыша (рис. 4 В, Г).
Наружную стенку его образует эктодерма, неоднородная в своем составе. В дорcальной части эктодерма утолщена и состоит из высоких цилиндрических клеток. Это зачаток нервной системы, которая остается


Гаструла ланцетника (по Мануиловой, 1973):
А – ранняя стадия; Б – поздняя стадия; В – поперечный разрез через позднюю гаструлу; Г – гаструла, переходящая в нейрулу (поперечный разрез)
еще на поверхности и образует так называемую медуллярную или нервную пластинку. Остальная эктодерма состоит из мелких клеток и является зачатком покрова животного. Под нервной пластинкой во внутреннем зародышевом листке располагается зачаток хорды, по обеим сторонам которого в виде двух тяжей находится материал мезодермы. В брюшной части располагается энтодерма, образующая основание первичной кишки, крышу которой составляют зачатки хорды и мезодермы .
Материал будущих внутренних органов, находясь в бластуле снаружи, в процессе гаструляции перемещается внутрь зародыша и располагается на местах, развивающихся из них органов. Только зачаток нервной системы остается еще на поверхности. Он погружается внутрь зародыша на стадии, следующей за гаструлой.
НЕЙРУЛЯЦИЯ И ОБРАЗОВАНИЕ ОСЕВЫХ ОРГАНОВ
По окончании гаструляции начинается следующий этап в развитии зародыша – дифференцировка зародышевых листков и закладка органов. Наличие комплекса спинных органов: нервной трубки, хорды и осевой мускулатуры, известных также под именем осевых, является одной их
характерных черт типа хордовых.
Стадия, на которой происходит закладка осевых органов, называется нейрулой. Внешне она характеризуется изменениями, происходящими с зачатком нервной системы.
Они начинаются с нарастания эктодермы по краям нервной пластинки. Образующиеся нервные валики растут навстречу друг другу и затем смыкаются. Пластинка же погружается внутрь и сильно прогибается (рис. 5).

Рис. 5. Нейрула ланцетника (по Мануиловой, 1973):
А – ранняя стадия (поперечный разрез); Б – поздняя стадия (поперечный
разрез), буквой “ Ц ” обозначена вторичная полость тела (целом)
Это приводит к образованию желобка, а затем нервной трубки, которая в передней и задней части зародыша некоторое время остается открытой (указанные изменения удобнее всего проследить на поперечном разрезе зародыша). Вскоре, в задней части тела эктодерма нарастает на бластопор и отверстие нервной трубки, закрывая их таким образом, что нервная трубка остается сообщенной с кишечной полостью – образуется нервно-кишечный канал.
Одновременно с формированием нервной трубки существенные изменения происходят и во внутреннем зародышевом листке. Из него постепенно обособляются материалы будущих внутренних органов. Зачаток хорды начинает выгибаться, выделяется из общей пластинки и превращается в обособленный тяж в виде сплошного цилиндра. Одновременно происходит обособление мезодермы. Этот процесс начинается с появления небольших карманообразных выростов по двум сторонам
внутреннего листка. По мере роста они отделяются от энтодермы и в ви-де двух тяжей с полостью внутри располагаются по всей длине зародыша. Кроме продольных желобков от переднего конца первичного кишечника последовательно отчленяются еще две пары целомических мешков.
Таким образом, в развитии ланцетника имеется стадия, характеризующаяся наличием трех пар сегментов и свидетельствующая об эволюционном родстве ланцетника с трехсегментарными личинками полухордовых и иглокожих. У ланцетника ярко выражен энтероцельный способ
образования целома – его отшнуровка от первичного кишечника. Этот способ является исходным для всех вторичноротых животных, но почти ни у кого из вышестоящих позвоночных, за исключением круглоротых, с такой ясностью не представлен. После отделения хорды и мезодермы
края энтодермы постепенно сближаются в спинной части и в конце концов смыкаются, образуя замкнутую кишечную трубку.
В ходе дальнейшего развития мезодерма сегментируется: тяжи разделяются поперечно на первичные сегменты или сомиты. Из них образуются три основные закладки:
• дерматом формируется из наружной, обращенной к эктодерме стенке сомита, – из его клеток впоследствие возникает соединительная часть кожи, представленная преимущественно фибробластами;
• склеротом образуется из внутренней части сомита, примыкающей к хорде (низшие позвоночные) или к хорде и нервной трубке (высшие позвоночные) – представляет зачаток осевого скелета;
• миотом представляет часть сомита, расположенную между дерматомом и склеротомом – является зачатком всей поперечно-полосатой мускулатуры.
Дифференцировка сомитов у ланцетника протекает иначе, чем у позвоночных. Это различие выражается в том, что у позвоночных сегментируется только спинная часть мезодермальных тяжей, тогда как у ланцетника они полностью распадаются на сегменты. Последние вскоре разделяются на спинную часть – сомиты, и брюшную – спланхнотом.
Сомиты, из которых развивается туловищная мускулатура, остаются обособленными друг от друга, спланхнотомы же сливаются на каждой стороне, образуя левую и правую полости, которые затем объединяются под кишечной трубкой в общую вторичную полость тела (целом).
В развитии ланцетника, с одной стороны, ясно представлены черты типичных позвоночных (характерное расположение зачатков при гаструляции, формирование хорды из дорсальной стенки первичной кишки и нервной пластинки из дорсальной эктодермы), а с другой – черты беспозвоночных вторичноротых животных (целобластула, инвагинационная гаструла, трехсегментная стадия, энтероцельная закладка мезодермы и образования целома).
В дальнейшем, в связи с образованием хвоста нервно-кишечный канал исчезает. В головной части кишечной трубки прорывается ротовое отверстие, а на заднем конце, под хвостом, образуется анальное – путем вторичного прорыва стенки тела животного на месте закрывшегося бластопора. Зародыш переходит в стадию свободноплавающей личинки.

Половые железы. Половые органы по внешнему виду построены одинаково у самцов и у самок. Половые железы в количестве около 26 пар расположены метамерно. Содержимое половых желез выводится в атриум, а в наружную среду продукты половых органов поступают через атриопор. В воде происходит оплодотворение яйца.
Нервная система ланцетника
Нервная система в виде продольного стержня расположена над хордой. Нервный ствол начинается несколько отступя от переднего конца спинной струны, а задний конец его едва заметно заходит за тыльный отдел хорды. Внутри нервного стержня тянется канал—невроцель, расширяющийся в переднем отделе мозга в небольшую полость, которую обычно сравнивают с мозговым желудочком (рис. , 2). Передний конец мозга образует непарный выступ, тесно связанный с обонятельной ямкой посредством короткого нерва.
Подобно центральной нервной системе каждого позвоночного животного, мозг ланцетника слагается из нервных клеток, группирующихся около центрального канала, и нервных волокон, идущих от мозга к периферии (см. ниже в общем очерке строения тела позвоночных).
Рис. Передняя часть нервной трубки молодого ланцетника.
I , II , III — поперечные разрезы через переднюю часть нервной трубки; I —в области невропора; II —в области спинного расширения; III —в промежутке между этими областями; 1—хорда; 2—переднее расширение нервной трубки; 3—спинное расширение; 4—пигментное пятно; 5— невропор; 6—обонятельная ямка; 7—8 — две передние пары нервов.
В области переднего расширения нервной системы отходят две пары нервов, называемых головными (рис., 7 и 8). Первая пара начинается от брюшной стороны мозга, вторая—от его спиной части. Они содержат чувствительные волокна, иннервирующие передний конец туловища. От остальных частей нервного стержня соответственно каждому сегменту берут начало спинные нервы. Необходимо отметить, что нервы правой и левой сторон чередуются по сегментам, а не отходят друг против друга. Каждый нерв отходит двумя ветвями: спинная начинается одним корешком на верхней стороне мозга, а брюшная образуется многочисленными волокнами. Спинные нервы направляются к поперечным мышцам и коже; по своей функции они смешанного типа—чувствительные и двигательные; брюшные—исключительно двигательные.

Органы чувств ланцетника
По всему эпидермису кожного слоя ланцетника рассеяны отдельные чувствительные клетки, местами образующие скопления. Чувствительных клеток особенно много по краям ротового отверстия и на щупальцах. Слева на спинной стороне передней части тела помещается выстланная мерцательным эпителием ямка, которую считают органом обоняния (ямка Келликера). У личинки эта ямка сообщается с полостью переднего отдела мозга посредством отверстия (невропора).
На верхней стороне ротового воротника помещается небольшая ямка, выстланная мерцательным эпителием. Эту ямку Гатчека обычно считают за орган.
Рис. 2. Развитие ланцетника, образование бластулы (1, 2) и гаструлы (3, 4).
осязания. Между передними нервами лежит непарное пигментное пятно, которое считают глазом (рис., 4). Оно лишено хрусталика и специфических нервных элементов, характерных даже для примитивно устроенных глаз. Интересно указать, что скопления пигмента распределяются вдоль всего центрального мозгового канала, образуя так называемые глазки Гессе. Обычно каждый глазок слагается из чувствительной клетки, снабженной тонким палочковидным придатком; к подобной клетке прилегает другая со скоплением пигмента. Нет сомнения, что светочувствительность всего мозга ланцетника обусловлена прозрачностью его тела. Органы слуха совершенно отсутствуют.
Размножение ланцетника
Развитие ланцетника было прослежено с большой тщательностью целым рядом исследователей (Ковалевским, Гатчеком, Рей-Ланкестером и Конклином). Оказалось, что содержимые в неволе ланцетники легко мечут икру. Процесс икрометания происходит час спустя после солнечного захода. Интересно весьма быстрое развитие яйца. Дробление протекает в тот же вечер; к 11 часам ночи образуется путем впячивания одного из боков шаровидной бластулы двуслойный зародыш или гаструла. К 1 часу ночи подобная гаструла совершенно готова. В 3 часа ночи можно видеть, что зародыш при помощи ресничек вращается в оболочке яйца. В 5 часов утра у зародыша появляются выпячивания мезодермы, он прорывает оболочку и выходит наружу. Приблизительно через 36 часов после начала процесса развития появляется рот, почти одновременно с ним первая жаберная щель. Вскоре открывается порошица. Этот момент можно считать завершением собственно зародышевой жизни и переходом к личиночному существованию. Молодые личинки живут на глубине 30—40 м.
Дробление оплодотворенного яйца ланцетника полное и почти равномерное. В результате последовательных делений бластомер развивается шаровидная бластула, клетки нижнего полюса которой несколько крупнее верхних (рис. 2, 1, 2). Верхняя половина бластулы, где сосредоточены более мелкие клетки, называется анимальным полюсом ее. Относительно крупные клетки нижней части составляют растительный, или вегетативный, полюс бластулы. В дополнение к сказанному отметим, что в бластуле образуется внутренняя полость—б ластоцель. Далее наступает впячивание—и н в а г и н а ц и я—нижних клеток бластулы по направлению к слою верхних клеток анимального полюса. В результате этого процесса образуется двуслойная первоначально чашковидная гаструла с очень широким отверстием—бластопо-ром или гастропором (рис. 2, 3, 4). Вскоре полость гаструлы углубляется, а бластопор сильно суживается (рис. 3, 1). Как было указано, стенки гаструлы
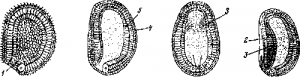
Рис. 3. Гаструла ланцетника и первые стадии образования личинки.
1 —бластопор; 2—закладка нервной системы; 3 —мезодерма; 4 —эктодерма; 5 —энтодерма.
двойные: наружный слой составляют более мелкие клетки эктодермы, образующие мерцательные жгутики,—благодаря их действию зародыш двигается еще в своей яйцевой оболочке; из внутренних, более крупных клеток диферен-цируется слой энтодермы (рис. 3, 5).
На этой ранней стадии развития намечается более плоская спинная сторона зародыша, тогда как брюшная часть заметно выпуклее. Далее наступает стадия, когда клетки эктодермы, образующие срединную часть плоской спинной стороны, обособляются и углубляются, давая начало медуллярной пластинке (рис. 4, А, 2). По бокам пластинки эктодермические клетки подымаются с каждой стороны в виде двух продольных валиков — медуллярных складок. Валики растут навстречу друг другу и смыкаются; сама медуллярная пластинка становится желобовидной (из плоской) (рис. 4, Б). В результате описанных процессов образуется нервная трубка с центральным внутренним каналом, или н е в р о ц е л е м. Тело личинки явно удлиняется в продольном направлении, медуллярные складки при своей закладке продолжаются за бластопор, который посредством особого нервно-кишечного канала сообщается с невроцелем. Нервная трубка открывается наружу на переднем конце туловища отверстием—невропором. В дальнейшем на этом месте развивается обонятельная ямка. Существенные картины диференцировки наблюдаются теперь и в энтодерме. На спинной стороне энтодермальной трубки, на так называемой первичной кишке, обособляются метамерные выступы— целомные карманы (рис.4, А, 6). Вскоре они смыкаются и отшнуровываются от первичной кишки и превращаются в целомные мешки (рис. 4, Б). Стенки этих целомных мешков состоят теперь из обособившегося слоя мезодермы, которая берет свое начало от энтодермы. В полостях этих мешков возникает вторичная полость тела—целом (энтероцель). От срединной, дорзальной, стенки первичной кишки обособляется борозда, стенки которой смыкаются, и борозда превращается в трубку хорды, или спинной струны, имеющей энтодермическое происхождение (рис. 4, 3).

Рис. 4. Развитие хорды, нервной системы и мезодермы у личинки ланцетника.
А —более ранняя стадия; Б—более поздняя стадия. 1—вырост эктодермы над медуллярной пластинкой; 2 —медуллярная пластинка; 3 — хорда; 4 — энтодерма; 5 —эктодерма; 6 —развитие мезодермы (образование целомных выростов).
Отдельные мезодермические целомные мешки начинают разрастаться по бокам тела зародыша, разделяясь каждый на спинной и брюшной отделы. Спинной отдел называется сомитом (рис. 5, 2), брюшной—боковой пластинкой. От нижнего внутреннего угла сомита обособляется особая складка ткани, окружающая плотным слоем хорду и нервную трубку. Эта складка называется склеротомом. Вся остальная часть сомита, называемая миотомом, внутренней полостью — миоцелем — разделяется на два листка: наружный, кожный, и внутренний, мускульный. Из наружного слоя в дальнейшем у личинки ланцетника развивается соединительнотканный клеточный слой, расположенный под эпидермисом. Клетки внутреннего листка дают начало миомерам, или мускульным сегментам.
В боковой пластинке, или в спланхнот оме, особая внутренняя полость (спланхноцель) отделяет прилегающий к кишечнику висцеральный листок от лежащего кнаружи париетального. Брюшной отдел мезодермы в спланхнотомах утрачивает метамерию: полости боковых пластинок (спланхноцели) соединяются между собой, образуя полость тела, или целом (см. выше).

Рис. 5. Зародыш ланцетника.
А—более ранняя стадия; Б —более поздняя стадия. 1—нервно-кишечный канал; 2—меводермические сомиты; 3—бластопор; 4 — нервная трубка;
Рис. 5. Зародыш ланцетника, окруженный зоной мерцательных ресничек.
1 —нервно-кишечный канал; 2—мезодермические сомиты; 3— бластопор; 4—нервная трубка;5—целомный мешок переднего отдела тела (rostrum).
Рис. 5. Личинка ланцетника.
1—невропор; 2— целомный мешок в отделе rosrtum; 3— хорда; 4—предротовой мешочек; 5 — железа; 6 — кишечник; 7 — пигментныепятна.
Рис. 5. Дальнейшая стадия развития личинки (обозначения, как на предыдущем рисунке).
У взрослого ланцетника, как мы могли видеть из предшествующего описания, полость тела очень сильно сокращена. Висце ральная внутренняя мускулатура возникает из глубокого, висцерального, листка боковой пластинки; париетальная же мускулатура развивается из миотомов.
Личинка быстро растет (рис. 5), становится длинной и тонкой; ее передний головной отдел заостряется; задний охватывается широким эмбриональным хвостовым плавником (рис. 5). На брюшной стороне в виде впячивания эктодермы, слева, появляется ротовое отверстие, которое сначала мало, а затем заметно увеличивается. Также на брюшной стороне показывается другое незначительное отверстие—ж аберная щель (рис. 6, 7).
В дальнейшем эта жаберная щель перемещается на правую сторону тела, существует недолго и вскоре зарастает. На правой стороне головного отдела появляется особая колбовидная железа, которая сначала имеет вид щели (видоизмененная жаберная щель), а затем превращается в заметный замкну тый мешок (рис. 5, 5). Колбовидная железа—личиночный орган, позднее совершенно исчезающий. По мере роста личинки в длину на брюшной стороне тела последовательно развиваются 14 жаберных щелей, постепенно перемещающихся на правую сторону. На той же правой стороне появляется новый ряд из 8 жаберных щелей, расположенных выше (дорзально) от упомянутых четырнадцати. Затем нижний ряд из 14 щелей снова передвигается на брюхо и далее на левую сторону, при этом количество их сокращается до восьми (исчезают первая и пять задних). Тогда расположение и число (8) жаберных щелей на обеих сторонах туловища становятся равными и симметричными.
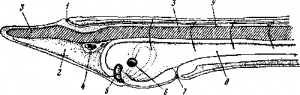
Рис. 6. Личинка ланцетника.
1—невропор; 2—целомный мешок в отделе rostrum; 3—хорда; 4—предротовой мешочек; 5—железа; 6— рот; 7 —закладка первой жаберной щели; 8—кишечник; 9—канал нервной системы.
Количество жаберных щелей в дальнейшем увеличивается; далее со спинной стороны каждой щели вырастает вниз в виде язычка перегородка, делящая щель надвое (вдоль). Количество щелей, вследствие этого процесса, удваивается. Перегородка, развившаяся из язычковидных выростов, представляет собой вторичные перегородки, характерные для взрослого ланцетника.
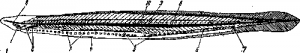
Рис. 7. Amphioxides pelagicus.
1—предротовая ямка; 2—эндостиль; 3—жаберные дужки; 4—метаплевральные складки; 5—гонады; 6— кишка; 7—анальное отверстие; 8—хорда; 9—нервная система; 10—пигментные пятна вдоль боков нервной трубки.
Одновременно с образованием жаберных щелей на брюшной стороне тела (соответственно расположению жаберных щелей) появляются продольные выросты—метаплевральные складки. Вскоре можно видеть, что на внутренней стороне каждой складки показываются продольные субатриальные выросты. Они растут навстречу друг другу, сходятся и образуют канал—зачаток атриальной полости. Первоначально эта полость узка, но потом начинает разрастаться вверх и в бока, охватывая собой жаберный отдел кишечной трубки и вытесняя полость целома. Атриальная полость, образующаяся из смыкания субатриальных выростов, первоначально открыта как спереди, так и сзади, но вскоре ее переднее отверстие замыкается, и остается открытым только заднее, превращающееся в атриопор.
От стенок полости тела в виде слепых выростов появляются нефридиальные канальцы, которые позднее получают отверстие в атриальную полость. Из нижних отделов миотомов развиваются в виде мешочков-гонотомов будущие половые железы, или гонады.
История развития ланцетника весьма интересна, так как представляет как бы упрощенную, схематическую картину эмбрионального развития позвоночных; кроме того, как было указано выше, в развитии ланцетника много общих этапов с развитием оболочников. Последнее явление имеет большой филогенетический интерес.
Статья на тему Размножение ланцетника
Гаструляция — результат активного деления клеток, роста и направленных перемещений (миграций) клеточных потоков с формированием многослойного зародыша, или гаструлы, (возникновением послойно расположенных, отделенных друг от друга отчетливой щелью, зародышевых листков: наружного — эктодермы, среднего — мезодермы, внутреннего — энтодермы). Перемещение клеток происходит в строго определенной области зародыша — в области серого серпа. Последний был описан В. Ру в 1888 г. В оплодотворенном яйце амфибии серый серп выявляется как окрашенная область на стороне, противоположной проникновению спермия. В этом месте, как полагают, локализуются факторы, необходимые для гаструляции.
У разных представителей позвоночных гаструляция совершается несколькими основными способами: путем инвагинации (впячивания), иммиграции (перемещения части клеток внутрь зародыша), эпиболии (обрастания), деламинации (расщепления). Способы гаструляции зависят от типа яйцеклетки. При любом способе гаструляции в качестве ведущих сил выступают неравномерная пролиферация клеток в разных частях зародыша, уровень обменных процессов в клетках, расположенных в разных частях зародыша, активность амебоидных движений клеток, а также индуктивные факторы (белки, нуклеопротеиды, стероиды и др.). В результате гаструляции обособляются основные зачатки органов и тканей.
Следующим периодом эмбриогенеза является гисто- и органогенез — дифференцировка различных тканей и органов организма из материала зародышевых листков и эмбриональных зачатков.
В результате гаструляции возникает многослойный зародыш. Несмотря на различные способы гаструляции после выделения материала зародышевых листков по оси зародыша находится материал хорды, который подстилает нервную пластинку, слева и справа от хорды располагается материал мезодермы. Все это характеризует осевой комплекс зачатков. В дальнейшем происходит формирование зачатков органов, представляющих собой пространственно локализованные группы стволовых клеток — источников развития тканей. Закономерности дифференцировки клеточного материала зачатков можно проследить в эмбриогенезе наиболее изученных животных.
Классическим объектом эмбриологических исследований ланцентник, подробно изученный А.О. Ковалевским. Ланцетник является представителем класса хордовых животных подтипа бесчерепных, размером до 8 см и обитает на песчаном дне в теплых морях. Свое название получил из-за формы, напоминающей ланцет (хирургический инструмент с обоюдоострым лезвием, современный скальпель).
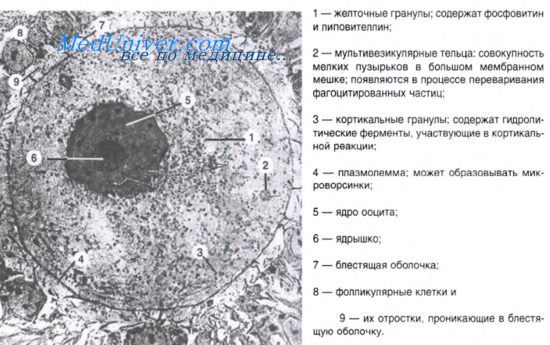
Яйцеклетка ланцетника олиго- и изолецитальная, размером 110 мкм, ядро располагается ближе к анимальному полюсу. Оплодотворение наружное. Дробление зиготы полное, почти равномерное, синхронное и заканчивается образованием бластулы. В результате чередования меридианальных и широтных борозд дробления формируется однослойная бластула с полостью, заполненной жидкостью — бластоцелем. Бластула сохраняет полярность, ее дно представляет собой вегетативную, а крыша — анимальную части; между ними находится краевая зона.
При гаструляции происходит впячивание вегетативной части бластулы в анимальную. Впячивание постепенно углубляется и, наконец, формируется двустенная чаша с широко зияющим отверстием, ведущим в новообразованную полость зародыша. Такой способ гаструляции называется инвагинацией. Так бластула превращается в гаструлу. В ней материал зародыша оказывается дифференцированным на наружный листок — эктодерму, и внутренний — энтодерму. Полость чаши именуется гастроцелем, или полостью первичной кишки, которая сообщается с внешней средой посредством бластопора, что соответствует заднепроходному отверстию. В бластопоре различают дорсальную, вентральную и две латеральных губы. В результате инвагинации центр тяжести зародыша смещается, и зародыш поворачивается бластопором кверху. Постепенно происходит смыкание краев бластопора и удлинение зародыша. Топография клеток в составе губ бластопора определяет развитие разных частей зародыша. При гаструляции из внутреннего листка гаструлы обособляются хорда и мезодерма, которые располагаются между экто- и энтодермой. Завершается гаструляция образованием осевого комплекса зачатков и дальше — обособлением зачатков органов. Хорда индуцирует развитие нервной трубки из материала дорсальной эктодермы. Эта часть эктодермы утолщается, образуется нервная пластинка (нейроэктодерма), которая прогибается по средней линии и превращается в желобок.
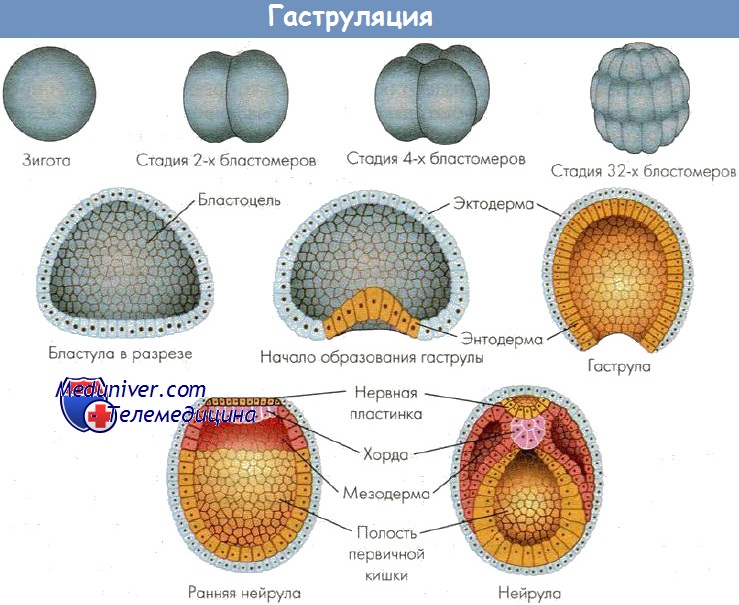
Края желобка постепенно смыкаются в нервную трубку. Оставшаяся часть эктодермы — кожная, срастается над нервной трубкой. Однако на самом переднем и заднем концах зародыша нервная трубка некоторое время сообщается с внешней средой с помощью двух отверстий — нейропоров. В последующем происходит разделение мезодермы на спинные сегменты — сомиты, количество которых нарастает от 15 пар до 60-65 пар у взрослого ланцетника. Часть латерально расположенной мезодермы не сегментируется и расщепляется на наружный (париетальный) и внутренний (висцеральный) листки спланхнотома. Эти листки растут между экто- и энтодермой и, достигнув середины на вентральной стороне зародыша под кишечной трубкой, срастаются, формируя единую вторичную полость — целом. На переднем конце зародыша возникает углубление (ротовая бухта), растущее по направлению к переднему отделу кишечной трубки. При контакте эктодермы ротовой бухты и слепого конца кишечной трубки происходит апоптоз клеток и возникает сообщение кишки с внешней средой. Аналогичный процесс протекает на заднем конце зародыша. По бокам головного отдела зародыша также возникает контакт кожной эктодермы и кишечной энтодермы. В месте этого контакта происходит прорыв. Так полость передней кишки сообщается с внешней средой (формируется жаберный аппарат). После этого зародыш выходит из яйцевой оболочки во внешнюю среду в виде личинки.
Методы маркирования для изучения процессов миграции бластомеров позволили выделить определенные области зародыша на ранних стадиях развития (зиготы — бластулы), которые позднее развиваются в зародышевые листки и эмбриональные зачатки органов и тканей. Эти области были названы презумптивными (предполагаемыми) участками, или зачатками.
Таким образом, на примере развития ланцетника можно наблюдать закономерное перемещение клеток и формирование зародышевых листков, осевого комплекса зачатков и обособление зачатков органов и тканей.
Читайте также:
