
Животная клетка нервная система
ГЛАВА 13. ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ
СИСТЕМЫ (ЦНС)
Центральная нервная система представителей животного мира включает множество различных групп нервных клеток и их многочисленных отростков, которые вступают в связь друг с другом и обеспечивают контакт организма с постоянно меняющейся окружающей средой, регуляцию, координацию и интеграцию всех процессов внутри организма.
В процессе эволюции нервная система претерпела значительные изменения, она все более и более совершенствовалась.
У сельскохозяйственных животных центральная нервная система представлена скоплением нервных клеток (нейронов), образующих головной и спинной мозг.
Основные функции, которые осуществляет центральная нервная система следующие:
- анализ поступающих раздражений из внешней и внутренней
среды,формирование ответных приспособительных реакций;
- обеспечение точной регуляции всех процессов внутри органи-
зма, их координацию и интеграцию, благодаря чему все орга-
ны и системы действуют согласованно;
- является материальным субстратом психических процессов -
ощущений, эмоций, памяти и других процессов, лежащих в
основе сложных форм поведения животных.
НЕЙРОННОЕ СТРОЕНИЕ И РЕФЛЕКТОРНАЯ
ДЕЯТЕЛЬНОСТЬ ЦНС. РЕФЛЕКСЫ И ИХ
КЛАССИФИКАЦИЯ
Центральная нервная система в морфологическом отношении состоит из двух компонентов - нервных клеток (нейронов) и нейроглии (совокупность всех других клеточных элементов нервной ткани).
Главной функциональной единицей (элементом ) ЦНС является нейрон. Количество нейронов в организме животных огромно (примерно 50 млрд). Нейроны составляют 10 - 15% общего количества клеточных элементов в нервной системе, а основную часть ее занимают клетки нейроглии. Считается, что глиальных клеток в 8-9 раз больше, чем нейронов.
Между глиальными клетки и нейронами имеются щели шириной 15-20 нм, которые сообщаются друг с другом, образуя интерстициальное пространство, заполненное жидкостью. Через это пространство происходит обмен веществ между нейроном и глиальными клетками, а также снабжение нейронов кислородом и питательными веществами путем диффузии.
Клетки нейроглии выполняют функции защиты и опоры нейронов.
Нейроны существенно различаются по форме (пирамидные, круглые, звездчатые, овальные), размером (от 5 до 150 мкм), по количеству отростков. В процессе постнатального онтогенеза нейроны не делятся.
Нейрон состоит из тела (сомы, перикариона) и отростков разного типа - дендритов (греч. dendron - дерево) и аксона (греч. axon - ось) (рис. 84).
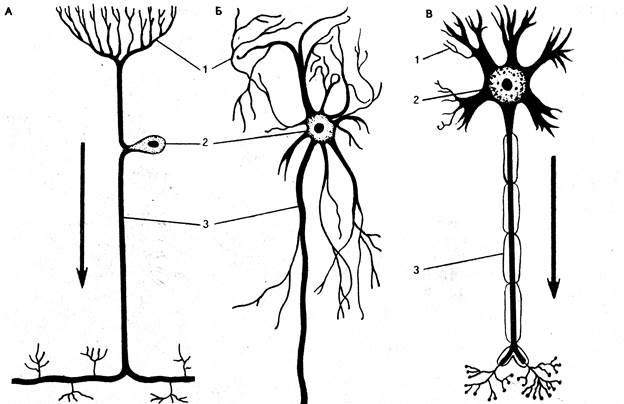
Рис. 84. Типы нейронов (схема):
А- сенсорный; Б- вставочный; В- эффекторный; 1- дендриты; 2- тело (сома) нейронов; 3- аксоны.
Для ЦНС позвоночных типичны биполярные (двухотростковые) и мультиполярные (многоотростковые) нейроны. Дендритов может быть много, иногда они сильно ветвятся, различной толщины и снабжены выступами, по которым нервные импульсы передаются к телу нейрона. Суммарная поверхность дендритов значительно превосходит поверхность тела нейрона. На дендритах размещено большое количество синапсов. Дендритам принадлежит ведущая роль в восприятии нейроном информации.
Аксон (нейрит) - удлиненный вырост цитоплазмы нейрона. Окруженный клетками олигодендроглии, он занимает центральное положение и поэтому называется осевым цилиндром. Все зрелые нейроны имеют один аксон. Он может ветвиться, образуя коллатерали и терминали. Структурно и функционально приспособлен для проведения возбуждения. Из аксонов формируются нервные стволы и проводящие пути нервной системы. Скорость проведения возбуждения по аксону возрастает как с увеличением его диаметра, так и при образовании вокруг него миелиновой оболочки.
Главная функция нейронов заключается в приеме сигналов от рецепторов или других нервных клеток, хранении и переработке информации и передача нервных импульсов к другим возбудимым клеткам - нервным, мышечным или секреторным.
В функциональном отношении нейроны подразделяют на 3 основных вида (класса, группы):
- чувствительные (сенсорные, афферентные) нейроны, которые воспринимают сигналы из внешней или внутренней среды;
- вставочные (промежуточные, ассоциативные) нейроны, которые связывают разные нервные клетки друг с другом;
- двигательные (эффекторные) нейроны, которые передают нисходящие влияния от вышерасположенных отделов ЦНС к нижерасположенным или из ЦНС к рабочим органам (эффекторам).
Чувствительные (афферентные) нейроны проводят возбуждение от рецепторов в центральную нервную систему. Тела этих нейронов расположены вне ЦНС и находятся в спинномозговых или черепно-мозговых ганглиях. Данные нейроны отличаются от других наличием двух длинных отростков: собственно аксона, по которому возбуждение передается от тела нервной клетки в центры спинного мозга или мозгового ствола, и аксоноподобного дендрита, уходящего на периферию в виде афферентного волокна и ветвящегося там на чувствительные нервные окончания -рецепторы.
К рецепторным нейронам относятся также некоторые нейроны в центральной нервной системе, которые получают возбуждение не непосредственно от рецепторов, а через другие, истинно рецепторные нейроны (например, нейроны зрительных бугров).
Вставочные (промежуточные, контактные нейроны, ассоциативные, интернейроны) образуют самую многочисленную группу нейронов. Они имеют более мелкий размер, звездчатую форму и аксоны с многочисленными разветвлениями и расположены в сером веществе мозга. Эти нейроны осуществляют связь между разными нейронами, например, афферентным и двигательным в пределах одного сегмента мозга или между соседними сегментами. Их отростки не выходят за пределы ЦНС.
По характеру вызываемого ими эффекта вставочные нейроны подразделяют на возбуждающие и тормозящие.
Двигательные (эфферентные, эффекторные) нейроны расположены в ЦНС. Их аксоны участвуют в передаче нисходящих влияний от вышерасположенных участков мозга к нижерасположенным или из ЦНС к рабочим органам - эффекторам. Те эфферентные нейроны, которые посылают импульсы к скелетным мышцам, называются мотонейронами. Их тела лежат в вентральных рогах спинного мозга, в продолговатом и среднем мозге. Имеются двигательные нейроны и в вегетативной нервной системе. Их тела лежат вне ЦНС - в периферических ганглиях. Особенностями этих нейронов являются разветвленная сеть дендритов и один длинный аксон.
Воспринимающей частью нейрона служат в основном дендриты, на которых имеется рецепторная мембрана. Возбуждение проходит по нейрону в одном направлении - от дендритов к соме и аксону.
Сложность и многообразие нервной системы зависит от взаимодействия между нейронами, которые, в свою очередь, представляют собой набор различных сигналов, передаваемых в рамках взаимодействия нейронов с другими нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов, генерирующих электрический заряд, который движется вдоль нейрона.
Строение
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Аксон обычно — длинный отросток, приспособленный для проведения возбуждения от тела нейрона. Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Си́напс — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона, другие — гиперполяризацию; первые являются возбуждающими, вторые — тормозящими. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Классификация
На основании числа и расположения дейндритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны - небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны - нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге.
Биполярные нейроны - нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах - сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
Мультиполярные нейроны - Нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе
Псевдоуниполярные нейроны - являются уникальными в своём роде. От тела отходит один остросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (т. е. находится вне тела клетки).
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятся конечные нейроны - ультиматные и предпоследние – неультиматные.
Ассоциативные нейроны (вставочные или интернейроны) - эта группа нейронов осуществляет связь между эфферентными и афферентными, их делят на комисуральные и проекционные (головной мозг).
Нервные клетки бывают звездчатые и веретенообразные, пирамидальные, зернистые, грушевидные и т.д.
Развитие и рост нейрона

Нейрон развивается из небольшой клетки — предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным. [1] (рус.) ) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — это область быстрого экзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
См. также
- Нервные волокна
- Аксон
- Дендрит
- Синапс
- Компьютер
- Искусственный нейрон
- Нейронная сеть
Сома · Аксон (Аксонный холмик, Терминаль аксона, Аксоплазма, Аксолемма, Нейрофиламенты)
3. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ФИЛОГЕНЕЗЕ
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. У одного и того же типа животных нервные клетки могут одновременно и независимо происходить из трех разных зародышевых листков. Полигенез нервных клеток беспозвоночных является основой разнообразия медиаторных механизмов их нервной системы.
Нервная система впервые появляется у кишечнополостных животных. Кишечнополостные — это двухслойные животные. Их тело представляет собой полый мешок, внутренняя полость которого является пищеварительной полостью. Нервная система кишечнополостных принадлежит к диффузному типу. Каждая нервная клетка в ней длинными отростками соединена с несколькими соседними, образуя нервную сеть. Нервные клетки кишечнополостных не имеют специализированных поляризованных отростков. Их отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками диффузной нервной системы бывают нескольких типов. Существуют плазматические контакты, обеспечивающие непрерывность сети (анастомозы). Появляются и щелевидные контакты между отростками нервных клеток, подобные синапсам. Причем среди них существуют контакты, в которых синаптические пузырьки располагаются по обе стороны контакта — так называемые симметричные синапсы, а есть и несимметричные синапсы: в них везикулы располагаются только с одной стороны щели.
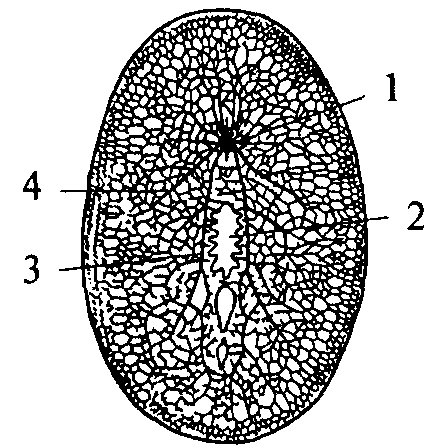
Нервные клетки типичного кишечнополостного животного гидры равномерно распределены по поверхности тела, образуя некоторые скопления в районе ротового отверстия и подошвы (рис. 8). Диффузная нервная сеть проводит возбуждение во всех направлениях. При этом волну распространяющегося возбуждения сопровождает волна мышечного сокращения.
Рис. 8. Схема строения диффузной нервной системы кишечнополостного животного:
1 — ротовое отверстие; 2 — щупальце; 3 — подошва
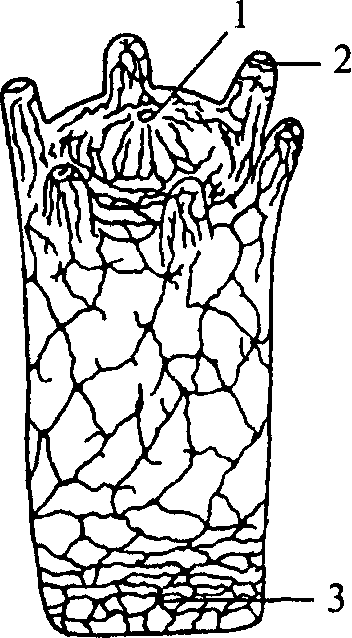
Рис. 9. Схема строения диффузностволовой нервной системы турбеллярии:
1 — нервный узел; 2 — глотка; 3 — брюшной продольный ствол; 4 — боковой нервный ствол
Следующим этапом развития беспозвоночных является появление трехслойных животных — плоских червей. Подобно кишечнополостным они имеют кишечную полость, сообщающуюся с внешней средой ротовым отверстием. Однако у них появляется третий зародышевый слой — мезодерма и двусторонний тип симметрии. Нервная система низших плоских червей принадлежит диффузному типу. Однако из диффузной сети уже обособляются несколько нервных стволов (рис. 9, 3, 4).
У свободно живущих плоских червей нервный аппарат приобретает черты централизации. Нервные элементы собираются в несколько продольных стволов (рис. 10, 4, 5) (для самых высокоорганизованных животных характерно наличие двух стволов), которые соединяются между собой поперечными волокнами (комиссурами) (рис. 10, 6). Упорядоченная таким образом нервная система называется ортогоном. Стволы ортогона представляют собой совокупность нервных клеток и их отростков (рис. 10).
1 — щупальцевидный вырост; 2 — нерв, иннервирующий вырост; 3 — мозговой ганглий; 4 — боковой продольный нервный ствол; 5 — брюшной продольный нервный ствол; 6 — комиссура
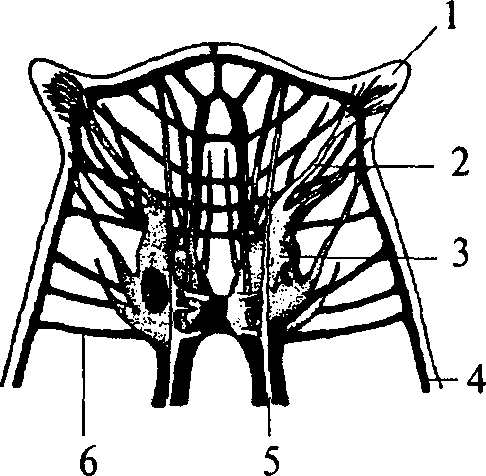
ганглия появляются длинные отростки, идущие в продольные стволы ортогона (рис. 10, 4, 5).
Таким образом, ортогон представляет собой первый шаг к централизации нервного аппарата и его цефализации (появлению мозга). Централизация и цефализация являются результатом развития сенсорных (чувствительных) структур.
Следующим этапом развития беспозвоночных животных является появление сегментированных животных — кольчатых червей. Их тело метамерно, т.е. состоит из сегментов. Структурной основой нервной системы кольчатых червей является ганглий — парное скопление нервных клеток, расположенных по одному в каждом сегменте. Нервные клетки в ганглии размещаются по периферии. Центральную его часть занимает нейропиль — переплетение отростков нервных клеток и глиальные клетки. Ганглий расположен на брюшной стороне сегмента под кишечной трубкой. Он посылает свои чувствительные и двигательные волокна в свой сегмент и в два соседних. Таким образом, каждый ганглий имеет три пары боковых нервов, каждый из которых является смешанным и иннервирует свой сегмент. Приходящие с периферии чувствительные волокна попадают в ганглий через вентральные корешки нервов. Двигательные волокна выходят из ганглия по дорсальным корешкам нервов. Соответственно этому чувствительные нейроны расположены в вентральной части ганглия, а двигательные — в дорсальной. Кроме того, в ганглии есть мелкие клетки, иннервирующие внутренние органы (вегетативные элементы), они расположены латерально — между чувствительными и двигательными нейронами. Среди нейронов чувствительной, двигательной или ассоциативной зон ганглиев кольчатых червей не обнаружено группирования элементов, нейроны распределены диффузно, т.е. не образуют центров.
Ганглии кольчатых червей соединены между собой в цепочку. Каждый последующий ганглий связан с предыдущим при помощи
1 — надглоточный нервный ганглий;
2 — подглоточный нервный ганглий;
3 — сложный слившийся ганглий грудного сегмента; 4 — брюшной ганглий; 5 — периферический нерв; 6 — коннектива
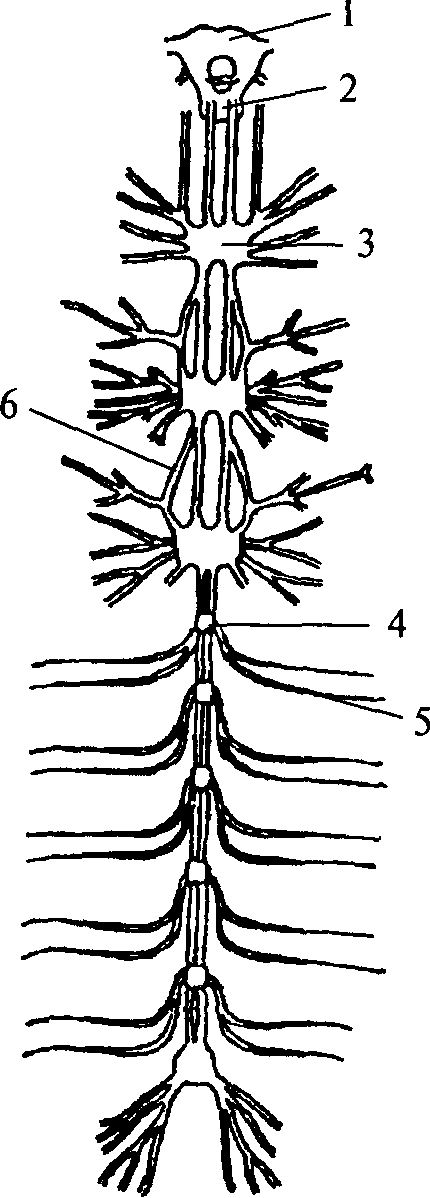
нервных стволов, которые называются коннективами. На переднем конце тела кольчатых червей два слившихся ганглия образуют крупный подглоточный нервный узел. Коннективы от подглоточного нервного узла, огибая глотку, вливаются в надглоточный нервный узел, который является самой ростральной (передней) частью нервной системы. В состав надглоточного нервного ганглия входят только чувствительные и ассоциативные нейроны. Двигательных элементов там не обнаружено. Таким образом, надглоточный ганглий кольчатых червей является высшим ассоциативным центром, он осуществляет контроль над подглоточным ганглием. Подглоточный ганглий контролирует нижележащие узлы, он имеет связи с двумя-тремя последующими ганглиями, тогда как остальные ганглии брюшной нервной цепочки не образуют связей длинней, чем до соседнего ганглия.
В филогенетическом ряду кольчатых червей есть группы с хорошо развитыми органами чувств (многощетинковые черви). У этих животных в надглоточном ганглии обособляются три отдела. Передний отдел иннервирует щупальца, средняя часть иннервирует глаза и антенны. И наконец, задняя часть развивается в связи с совершенствованием химических органов чувств.
Сходную структуру имеет нервная система членистоногих, т.е. построена по типу брюшной нервной цепочки, однако может достигать высокого уровня развития (рис. 11). Она включает в себя значительно развитый надглоточный ганглий, выполняющий функ-
1 — грибовидное тело; 2 — протоцеребрум; 3 — зрительная лопасть; 4 — дейтоцеребрум; 5 — тритоцеребрум
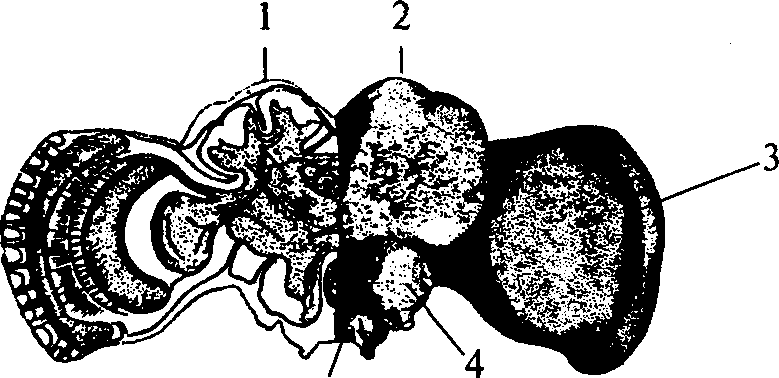
цию мозга, подглоточный ганглий, управляющий органами ротового аппарата, и сегментарные ганглии брюшной нервной цепочки. Ганглии брюшной нервной цепочки могут сливаться между собой, образуя сложные ганглиозные массы.
Головной мозг членистоногих состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Сложным строением отличается мозг насекомых. Особенно важными ассоциативными центрами насекомых являются грибовидные тела, располагающиеся на поверхности протоцеребрума, причем чем более сложным поведением характеризуется вид, тем более развиты у него грибовидные тела. Поэтому наибольшего развития грибовидные тела достигают у общественных насекомых (рис. 12).
Практически во всех отделах нервной системы членистоногих существуют нейросекреторные клетки. Нейросекреты играют важную регулирующую роль в гормональных процессах членистоногих.
В процессе эволюции первоначально диффузно расположенные биполярные нейросекреторные клетки воспринимали сигналы либо отростками, либо всей поверхностью клетки, затем сформировались нейросекреторные центры, нейросекреторные тракты и нейросекреторные контактные области. В последующем произошла специализация нервных центров, увеличилась степень надежности во взаимоотношениях двух основных регуляторных систем (нервной и гуморальной) и сформировался принципиально новый этап регуляции — подчинение нейросекреторным центрам периферических эндокринных желез.
1 — церебральная комиссура; 2 — церебральные ганглии; 3 — педальные ганглии; 4 — коннектива; 5 — висцеральные ганглии
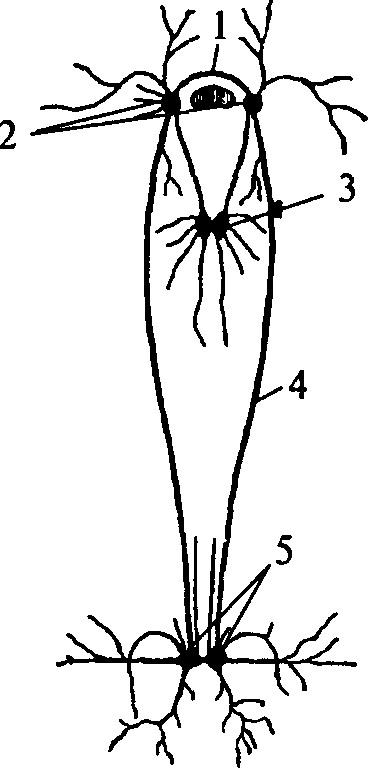
Нервная система моллюсков также имеет ганглионарное строение (рис. 13). У простейших представителей типа она состоит из нескольких пар ганглиев. Каждая пара ганглиев управляет определенной группой органов: ногой, висцеральными органами, легкими и т.д. — и расположена рядом с иннервируемыми органами или внутри их. Одноименные ганглии попарно соединены между собой комиссурами. Кроме того, каждый ганглий связан длинными коннективами с церебральным комплексом ганглиев.
У более высокоорганизованных моллюсков (головоногие) нервная система преобразуется (рис. 14). Ганглии ее сливаются и образуют общую окологлоточную массу — головной мозг. От заднего отдела головного мозга отходят два крупных мантийных нерва и образуют два больших звездчатых ганглия. Таким образом, у головоногих наблюдается высокая степень цефализации.
Структура нервной системы
Нервная система содержит две основных категории, или типа, клеток: нейроны и глиальные клетки.
Сравнительная анатомия и эволюция
Губки не имеют никаких клеток, соединенных друг с другом посредством синаптической щели, то есть, не имеют нейронов, и, следовательно, не имеют нервной системы. Они, однако, имеют гомологи многих генов, которые играют ключевую роль в синаптической функции. Недавние исследования показали, что клетки губок экспрессируют группу белков, которые группируются, формируя структуру, напоминающую постсинаптическую плотность (часть синапса, передающую сигналы). Тем не менее, функция этой структуры в настоящее время неясна. Хотя клетки губок не демонстрируют синаптической передачи, они общаются друг с другом с помощью волн кальция и других импульсов, которые опосредуют некоторые простые действия, такие как сжатие всего организма. 10)
Медузы, гребневики и связанные с ними животные имеют диффузные нервных сети, а не центральную нервную систему. У большинства медуз, нервная сеть распределяется более или менее равномерно по всему телу; у гребневиков она сосредоточена вблизи рта. Нервные сети состоят из сенсорных нейронов, которые принимают химические, тактильные и визуальные сигналы; двигательных нейронов, которые могут активировать сокращения стенок организма; и промежуточных нейронов, которые обнаруживают паттерны активности в сенсорных нейронах и, в ответ, посылают сигналы в группы двигательных нейронов. В некоторых случаях, промежуточные нейроны группируются в дискретные ганглии. Развитие нервной системы у радиальных относительно неструктурировано. В отличие от билатерий, радиальные имеют только два изначальных слоя клеток, энтодерму и эктодерму. Нейроны генерируются из специального набора эктодермальных клеток-предшественников, которые также служат в качестве предшественников для любого другого эктодермального типа клеток. 11)
Членистоногие, такие как насекомые и ракообразные, имеют нервную систему, состоящую из ряда ганглий, соединенных вентральной нервной цепочкой, состоящей из двух параллельных связок, проходящей вдоль живота. Как правило, каждый сегмент тела имеет по одном ганглии на каждой стороне, хотя некоторые ганглии сплавлены, образуя мозг и другие крупные ганглии. Сегмент головки содержит мозг, также известный как надглоточный ганглий. В нервной системе насекомых, мозг анатомически делится на протоцеребрум, дейтоцеребрум и триоцеребрум. Сразу за мозгом располагается подглоточный ганглий, который состоит из трех пар сплавленных ганглиев. Он контролирует ротовые, слюнные железы и некоторые мышцы. Многие членистоногие имеют хорошо развитые органы чувств, в том числе, сложные глаза для зрения и усики для обоняния и ощущения феромонов. Сенсорная информация, поступающая от этих органов, обрабатывается мозгом насекомых. У насекомых, многие нейроны имеют клеточные тела, которые расположены на краю мозга и являются электрически пассивными – клетки тела служат только для обеспечения поддержки обмена веществ и не участвуют в передаче сигналов. Протоплазматическое волокно обильно проходит от тела и ветвей клеток, при этом некоторые части передают сигналы, в то время как другие части принимают сигналы. Таким образом, большинство частей мозга насекомого имеют пассивные клеточные тела, расположенные по периферии, в то время как обработка нервного сигнала происходит в волокнах протоплазмы, называемых нейропилем, в передней части. 15)
Функция
Развитие
Патология
Центральная нервная система защищена основными физико-химическими барьерами. Физически, головной и спинной мозг окружены жесткими менингеальными оболочками, и заключены в костях черепа и позвонках, которые объединяются, формируя сильную физическую защиту. Химически, мозг и спинной мозг изолированы так называемым гематоэнцефалическим барьером, который предотвращает перемещение большинства видов химических веществ из кровотока во внутреннюю часть центральной нервной системы. Эти средства защиты делают во многих отношениях ЦНС менее восприимчивой, чем ПНС; обратной стороной, однако, является то, что повреждение центральной нервной системы, как правило, имеют более серьезные последствия. Хотя нервы, как правило, лежат глубоко под кожей, за исключением нескольких участков, таких как локтевой нерв около локтевого сустава, они все еще относительно подвержены физическим повреждениям, которые могут причинить боль, вызвать потерю чувствительности, или потерю мышечного контроля. Повреждение нервов может также быть вызвано припухлостью или гематомой в местах, где нерв проходит через плотный костяной канал, как это происходит при кистевом туннельном синдроме. Если полностью перерезать нерв, он будет часто регенерировать, но у длинных нервов этот процесс может занять несколько месяцев. В дополнение к физическому повреждению, периферическая невропатия может быть вызвана многими другими медицинскими проблемами, в том числе, генетическими заболеваниями, метаболическими заболеваниями, такими как диабет, воспалительными состояниями, такими как синдром Гийена-Барре, витаминной недостаточностью, инфекционными заболеваниями, такими как лепра или опоясывающий лишай, или отравлением токсинами, такими как тяжелые металлы. Во многих случаях, нет никаких причин, которые могут быть идентифицированы, и называться идиопатическими. Для нервов также возможна временная потеря функции, в результате приводящая к нечувствительности – общие причины такой нечувствительности включают в себя механическое давление, понижение температуры или химическое взаимодействие с местными обезболивающими препаратами, такими как лидокаин. Физическое повреждение спинного мозга может привести к потере чувствительности или нарушению движений. Если травма позвоночника вызывает только отеки, симптомы могут быть временными, однако, если нервные волокна в позвоночнике фактически уничтожены, потеря функции, как правило, является постоянной. Экспериментальные исследования показали, что спинномозговые нервные волокна пытаются повторно расти таким же образом, как и нервные волокна, но в спинном мозге разрушение ткани обычно производит рубцовую ткань, которая не может быть пронизана заново растущими нервами.
3. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ФИЛОГЕНЕЗЕ
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. У одного и того же типа животных нервные клетки могут одновременно и независимо происходить из трех разных зародышевых листков. Полигенез нервных клеток беспозвоночных является основой разнообразия медиаторных механизмов их нервной системы.
Нервная система впервые появляется у кишечнополостных животных. Кишечнополостные — это двухслойные животные. Их тело представляет собой полый мешок, внутренняя полость которого является пищеварительной полостью. Нервная система кишечнополостных принадлежит к диффузному типу. Каждая нервная клетка в ней длинными отростками соединена с несколькими соседними, образуя нервную сеть. Нервные клетки кишечнополостных не имеют специализированных поляризованных отростков. Их отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками диффузной нервной системы бывают нескольких типов. Существуют плазматические контакты, обеспечивающие непрерывность сети (анастомозы). Появляются и щелевидные контакты между отростками нервных клеток, подобные синапсам. Причем среди них существуют контакты, в которых синаптические пузырьки располагаются по обе стороны контакта — так называемые симметричные синапсы, а есть и несимметричные синапсы: в них везикулы располагаются только с одной стороны щели.
Нервные клетки типичного кишечнополостного животного гидры равномерно распределены по поверхности тела, образуя некоторые скопления в районе ротового отверстия и подошвы (рис. 8). Диффузная нервная сеть проводит возбуждение во всех направлениях. При этом волну распространяющегося возбуждения сопровождает волна мышечного сокращения.
Рис. 8. Схема строения диффузной нервной системы кишечнополостного животного:
1 — ротовое отверстие; 2 — щупальце; 3 — подошва
Рис. 9. Схема строения диффузностволовой нервной системы турбеллярии:
1 — нервный узел; 2 — глотка; 3 — брюшной продольный ствол; 4 — боковой нервный ствол
Следующим этапом развития беспозвоночных является появление трехслойных животных — плоских червей. Подобно кишечнополостным они имеют кишечную полость, сообщающуюся с внешней средой ротовым отверстием. Однако у них появляется третий зародышевый слой — мезодерма и двусторонний тип симметрии. Нервная система низших плоских червей принадлежит диффузному типу. Однако из диффузной сети уже обособляются несколько нервных стволов (рис. 9, 3, 4).
У свободно живущих плоских червей нервный аппарат приобретает черты централизации. Нервные элементы собираются в несколько продольных стволов (рис. 10, 4, 5) (для самых высокоорганизованных животных характерно наличие двух стволов), которые соединяются между собой поперечными волокнами (комиссурами) (рис. 10, 6). Упорядоченная таким образом нервная система называется ортогоном. Стволы ортогона представляют собой совокупность нервных клеток и их отростков (рис. 10).
1 — щупальцевидный вырост; 2 — нерв, иннервирующий вырост; 3 — мозговой ганглий; 4 — боковой продольный нервный ствол; 5 — брюшной продольный нервный ствол; 6 — комиссура
ганглия появляются длинные отростки, идущие в продольные стволы ортогона (рис. 10, 4, 5).
Таким образом, ортогон представляет собой первый шаг к централизации нервного аппарата и его цефализации (появлению мозга). Централизация и цефализация являются результатом развития сенсорных (чувствительных) структур.
Следующим этапом развития беспозвоночных животных является появление сегментированных животных — кольчатых червей. Их тело метамерно, т.е. состоит из сегментов. Структурной основой нервной системы кольчатых червей является ганглий — парное скопление нервных клеток, расположенных по одному в каждом сегменте. Нервные клетки в ганглии размещаются по периферии. Центральную его часть занимает нейропиль — переплетение отростков нервных клеток и глиальные клетки. Ганглий расположен на брюшной стороне сегмента под кишечной трубкой. Он посылает свои чувствительные и двигательные волокна в свой сегмент и в два соседних. Таким образом, каждый ганглий имеет три пары боковых нервов, каждый из которых является смешанным и иннервирует свой сегмент. Приходящие с периферии чувствительные волокна попадают в ганглий через вентральные корешки нервов. Двигательные волокна выходят из ганглия по дорсальным корешкам нервов. Соответственно этому чувствительные нейроны расположены в вентральной части ганглия, а двигательные — в дорсальной. Кроме того, в ганглии есть мелкие клетки, иннервирующие внутренние органы (вегетативные элементы), они расположены латерально — между чувствительными и двигательными нейронами. Среди нейронов чувствительной, двигательной или ассоциативной зон ганглиев кольчатых червей не обнаружено группирования элементов, нейроны распределены диффузно, т.е. не образуют центров.
Ганглии кольчатых червей соединены между собой в цепочку. Каждый последующий ганглий связан с предыдущим при помощи
1 — надглоточный нервный ганглий;
2 — подглоточный нервный ганглий;
3 — сложный слившийся ганглий грудного сегмента; 4 — брюшной ганглий; 5 — периферический нерв; 6 — коннектива
нервных стволов, которые называются коннективами. На переднем конце тела кольчатых червей два слившихся ганглия образуют крупный подглоточный нервный узел. Коннективы от подглоточного нервного узла, огибая глотку, вливаются в надглоточный нервный узел, который является самой ростральной (передней) частью нервной системы. В состав надглоточного нервного ганглия входят только чувствительные и ассоциативные нейроны. Двигательных элементов там не обнаружено. Таким образом, надглоточный ганглий кольчатых червей является высшим ассоциативным центром, он осуществляет контроль над подглоточным ганглием. Подглоточный ганглий контролирует нижележащие узлы, он имеет связи с двумя-тремя последующими ганглиями, тогда как остальные ганглии брюшной нервной цепочки не образуют связей длинней, чем до соседнего ганглия.
В филогенетическом ряду кольчатых червей есть группы с хорошо развитыми органами чувств (многощетинковые черви). У этих животных в надглоточном ганглии обособляются три отдела. Передний отдел иннервирует щупальца, средняя часть иннервирует глаза и антенны. И наконец, задняя часть развивается в связи с совершенствованием химических органов чувств.
Сходную структуру имеет нервная система членистоногих, т.е. построена по типу брюшной нервной цепочки, однако может достигать высокого уровня развития (рис. 11). Она включает в себя значительно развитый надглоточный ганглий, выполняющий функ-
1 — грибовидное тело; 2 — протоцеребрум; 3 — зрительная лопасть; 4 — дейтоцеребрум; 5 — тритоцеребрум
цию мозга, подглоточный ганглий, управляющий органами ротового аппарата, и сегментарные ганглии брюшной нервной цепочки. Ганглии брюшной нервной цепочки могут сливаться между собой, образуя сложные ганглиозные массы.
Головной мозг членистоногих состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Сложным строением отличается мозг насекомых. Особенно важными ассоциативными центрами насекомых являются грибовидные тела, располагающиеся на поверхности протоцеребрума, причем чем более сложным поведением характеризуется вид, тем более развиты у него грибовидные тела. Поэтому наибольшего развития грибовидные тела достигают у общественных насекомых (рис. 12).
Практически во всех отделах нервной системы членистоногих существуют нейросекреторные клетки. Нейросекреты играют важную регулирующую роль в гормональных процессах членистоногих.
В процессе эволюции первоначально диффузно расположенные биполярные нейросекреторные клетки воспринимали сигналы либо отростками, либо всей поверхностью клетки, затем сформировались нейросекреторные центры, нейросекреторные тракты и нейросекреторные контактные области. В последующем произошла специализация нервных центров, увеличилась степень надежности во взаимоотношениях двух основных регуляторных систем (нервной и гуморальной) и сформировался принципиально новый этап регуляции — подчинение нейросекреторным центрам периферических эндокринных желез.
1 — церебральная комиссура; 2 — церебральные ганглии; 3 — педальные ганглии; 4 — коннектива; 5 — висцеральные ганглии
Нервная система моллюсков также имеет ганглионарное строение (рис. 13). У простейших представителей типа она состоит из нескольких пар ганглиев. Каждая пара ганглиев управляет определенной группой органов: ногой, висцеральными органами, легкими и т.д. — и расположена рядом с иннервируемыми органами или внутри их. Одноименные ганглии попарно соединены между собой комиссурами. Кроме того, каждый ганглий связан длинными коннективами с церебральным комплексом ганглиев.
У более высокоорганизованных моллюсков (головоногие) нервная система преобразуется (рис. 14). Ганглии ее сливаются и образуют общую окологлоточную массу — головной мозг. От заднего отдела головного мозга отходят два крупных мантийных нерва и образуют два больших звездчатых ганглия. Таким образом, у головоногих наблюдается высокая степень цефализации.
Читайте также:
