
Ацетилхолин его роль передачи нервных импульсов
Связь миллиардов нейронов мозга осуществляется посредством медиаторов. Химическое вещество можно отнести к числу медиаторов лишь в том случае, если оно удовлетворяет ряду критериев. В нервных волокнах должны содержаться ферменты, необходимые для синтеза этого вещества. При раздражении нервов это вещество должно выделяться, реагировать со специфическим рецептором на постсинаптической клетке и вызывать биологическую реакцию. Должны существовать механизмы, быстро прекращающие действие этого вещества.
Всем этим критериям удовлетворяют два вещества – ацетилхолин и но-радреналин. Содержащие их нервы называют соответственно холинерги-ческими и адренергическими. В соответствии с этим все эфферентные системы делят на холинорецепторы и адренорецепторы.
Ряд других химических веществ удовлетворяют многим, но не всем перечисленным критериям. К таким медиаторам относят дофамин, адреналин, серотонин, октопамин, гистамин, ГАМК и др.
Обширная группа холинорецепторов весьма неоднородна как в структурном, так и в функциональном отношении. Объединяют их медиатор ацетилхолин и общая схема строения синапса.
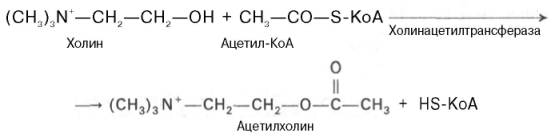
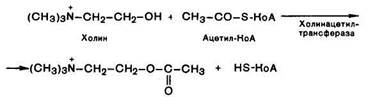
Ацетилхолин представляет собой сложный эфир уксусной кислоты и холина. Он синтезируется в нервной клетке из холина и активной формы ацетата – ацетилкоэнзима А при помощи специального фермента холин-ацетилтрансферазы (холинацетилазы):
Синапс можно представить себе как узкое пространство (щель), ограниченное с одной стороны пресинаптической, а с другой – постсинапти-ческой мембраной (рис. 19.4). Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других истончена и имеет отверстия для сообщения цитоплазмы аксона с синаптическим пространством. Постсинапти-ческая мембрана менее плотная, не имеет отверстий. Подобным образом построены и нервно-мышечные синапсы, но они имеют более сложное строение мембранного комплекса.
В общих чертах картину участия ацетилхолина в осуществлении передачи нервного импульса возбуждения можно представить следующим образом. В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30–80 нм, которые содержат нейромедиаторы. Эти пузырьки покрыты оболочкой, которая образована белком клатрином (мол. масса 180000). В холинергических синапсах каждый пузырек диаметром 80 нм содержит
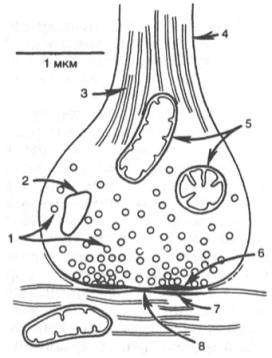
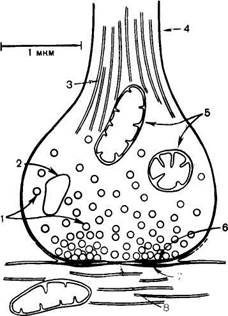
Рис. 19.4. Схематическое изображение синапса (по Мецлеру).
1 - синаптические пузырьки; 2 - лизосо-ма; 3 - микрофибриллы (нейрофибрил-лы); 4 - аксон; 5 - митохондрии; 6 - пре-синаптическое утолщение мембраны; 7 - постсинаптическое утолщение мембраны; 8 - синаптическая щель (около 20 нм).
В холинергических синапсах это происходит двумя путями. Первый путь заключается в том, что ацетилхолин подвергается ферментативному гидролизу. Второй путь – это энергозависимый активный транспорт ацетилхо-лина в нейрон, где он накапливается для последующего повторного использования.
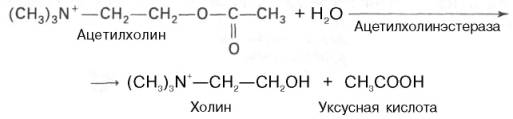
В большинстве отделов головного мозга гидролиз ацетилхолина осуществляется ацетилхолинэстеразой (истинная холинэстераза, которая гид-ролизует ацетилхолин быстрее, чем иные эфиры холина). В нервной ткани существуют и другие эстеразы, которые способны гидролизовать ацетил-холин, но значительно медленнее, чем, например, бутирилхолин. Эти эстеразы называются холинэстеразой (или псевдохолинэстеразой). К числу холинергических систем относятся моторные нейроны, образующие нервно-мышечные соединения, все преганглионарные нейроны автономной нервной системы и постганглионарные нейроны парасимпатической нервной системы. Большое количество холинергических симпатических областей обнаружено также в головном мозге. В зависимости от чувствительности к той или иной группе химических соединений холинергические нейроны делятся на мускариновые (активируемые мускарином) и никотиновые (активируемые никотином). Мускариновые рецепторы ацетилхо-лина, имеющиеся во многих нейронах автономной нервной системы, специфически блокируются атропином. Никотиновые синапсы присутствуют в ганглиях и скелетных мышцах. Их ингибиторами являются кураре и активный компонент этого яда D-тубокурарин.
Необходимо подчеркнуть, что в адренорецепторах существует два вида рецепторов для норадреналина: α- и β-адренергические рецепторы. Эти рецепторы можно отличить друг от друга по специфическим реакциям, которые они вызывают, а также по тем специфическим агентам, которые способны блокировать данные реакции.
Установлено, что как только β-адренергический рецептор, расположенный на наружной поверхности мембраны эффекторной клетки, начинает взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Этот сложный ряд последовательных реакций может быть заблокирован пропраноло-лом – веществом, препятствующим связыванию норадреналина с β-адре-нергическим рецептором.
Известно, что в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (—NH2) у норадреналина, серотонина, дофамина и адреналина, тем самым инактивируя указанные медиаторы. В последние годы было показано, что, помимо ферментативного превращения, существует и другой механизм быстрой инактивации, точнее удаления, медиаторов. Оказалось, что норадреналин быстро исчезает из синаптической щели в результате вторичного поглощения симпатическими нервами; вновь оказавшись в нервном волокне, медиатор, естественно, не может воздействовать на постсинаптические клетки. Конкретный механизм этого явления пока не вполне ясен.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности использующими серотонин в качестве медиатора. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами. Исследования, проведенные с ингибитором синтеза серотонина n-хлорфенилаланином, а также с другими ингибиторами, дают основания считать, что серотонин влияет на процессы сна. Выявлено также, что торможение кортикостероидами секреторной активности гипофиза оказывается менее эффективным у тех животных, мозг которых беднее серотонином.
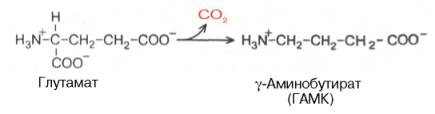
Важным нейромедиатором, выполняющим тормозные функции, является γ-аминомасляная кислота (ГАМК), количество которой в головном мозге во много раз больше, чем других нейромедиаторов. Так, в гипоталамусе суммарное содержание ацетилхолина, норадреналина, дофамина и серотонина не превышает 10 мкг/г, в то время как ГАМК в этом отделе головного мозга более 600 мкг/г. ГАМК увеличивает проницаемость постсинаптических мембран для ионов К + и тем самым отдаляет мембранный потенциал от порогового уровня, при котором возникает потенциал действия; таким образом, ГАМК – это тормозной нейромедиатор. ГАМК образуется при декарбоксилировании глутамата в реакции, катализируемой глутаматдекарбоксилазой:
В терапевтической практике применяется большое количество лекарственных средств, которые действуют через систему медиаторов. Многие лекарственные препараты, успешно применяемые при лечении гипертонии, влияют на накопление и выделение адренергических медиаторов. Например, резерпин – понижающее артериальное давление средство специфически тормозит процесс переноса катехоламинов в специальные гранулы нейронов и тем самым делает эти амины доступными действию эндогенной МАО.
Многие антидепрессанты (вещества, снимающие депрессию) увеличивают содержание катехоламинов в синаптической щели, т.е. количество медиаторов для стимулирования рецептора возрастает. К таким веществам, в частности, относятся имипрамин (блокирует поглощение норадреналина нервными волокнами), амфетамин (одновременно способствует выделению норадреналина и блокирует его поглощение), ингибиторы МАО (подавляют метаболизм катехоламинов) и др. В связи с этим возникла катехол-аминовая гипотеза депрессивных состояний, согласно которой психическая депрессия связана с недостатком катехоламинов в мозге.
В начале 50-х годов фармакологи выяснили, что известный галлюциноген диэтиламин лизергиновой кислоты (ЛСД) не только сходен по химическому строению с серотонином, но и нейтрализует некоторые его фармакологические эффекты (блокируя рецепторы серотонина). Поэтому было высказано предположение, что нарушение обмена серотонина может быть причиной возникновения особых психических заболеваний.
Считают, что такие антипсихотические средства, как аминазин (хлор-промазин) и галоперидол, усиливая синтез катехоламинов, способны блокировать дофаминовые рецепторы в мозге.
Связь миллиардов нейронов мозга осуществляется посредством медиаторов — ацетилхолин и норадреналин. Содержащие их нервы называют соответственно холинергическими и адренергическими. В соответствии с этим все эфферентные системы делят на холинорецепторы и адренорецепторы.
К другим медиаторам относятся дофамин, адреналин, серотонин, октопамин, гистамин, ГАМК и др.
Обширная группа холинорецепторов весьма неоднородна как в структурном, так и в функциональном отношении. Объединяют их медиатор — ацетилхолин — и общая схема строения синапса.
 |
Ацетилхолин представляет собой сложный эфир уксусной кислоты и холина. Он синтезируется в нервной клетке из холина и активной формы ацетата — ацетилкоэнзима А при помощи специального фермента холинацетилтрансферазы (холйнацетилазы):
| Схематическое изображение синапса (по Мецлеру). 1 — синаптические пузырьки; 2 — лизосома; 3 — микрофибриллы (нейрофибриллы); 4 - аксон; 5 - митохондрии; 6 — пресинаптическое утолщение мембраны; 7 - постсинаптическое утолщение мембраны; 8 — синаптическая щель (около 20 нм). |
Синапс можно представить себе как узкое пространство (щель), ограниченное с одной стороны пресинаптической, с другой — постсинаптической мембраной. Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других — истончена и имеет отверстия, с помощью которых цитоплазма аксона может сообщаться с синаптическим пространством. Постсинаптическая мембрана менее плотная, не имеет отверстий.
В адренорецепторах существует два вида рецепторов для норадреналина: α- и β-адренергические рецепторы.
Как только β-адренергический рецептор (расположенный на наружной поверхности мембраны эффекторной клетки) начнет взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Известно, что в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (-NH2) у норадреналина, серотонйна, дофамина и адреналина, тем самым инактивируя упомянутые медиаторы.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности использующими серотонин в качестве медиатора. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами.
Важным нейромедиатором, выполняющим тормозные функции, является ГАМК, количество которой в головном мозге во много раз выше, чем других нейромедиаторов.
Ацетилхолин осуществляет передачу нервных импульсов в холинергических синапсах. Открытие медиаторной роли ацетилхолина принадлежит австрийскому фармакологу О. Леви (Loewi). Холинергические синапсы имеются как в соматической, так и вегетативной нервной системе. Двигательные волокна соматической нервной системы иннервируют скелетные мышцы, и с их окончаний выделяется ацетилхолин. Эфферентные проводящие пути вегетативной нервной системы состоят из двух нейронов: первый расположен в центральной нервной системе (в стволе головного мозга и спинном мозге), второй — в вегетативном ганглии, который относится к периферической нервной системе (рис.
Таким образом, ацетилхолин служит передатчиком импульсов с окончаний всех парасимпатических постганглионарных волокон, с окончаний постганглионарных симпатических волокон, иннервирующих потовые железы, с окончаний всех (как симпатических, так и парасимпатических) преганглионарных волокон, с окончаний двигательных нервов поперечно-полосатых мышц, а также во многих центральных синапсах.
Химически ацетилхолин представляет собой сложный эфир холина и уксусной кислоты. Его синтез проходит в окончаниях нервных волокон из спирта холина и ацетил-КоА под влиянием фермента холинацетилтрансферазы. Скорость реакции синтеза лимитируется концентрацией холина в синаптических окончаниях. Синтезированный медиатор депонируется в везикулах в результате активного транспорта с участием фермента — Mg^-зависимая АТФаза. Основным механизмом выделения ацетилхолина в синаптическую щель, в результате чего формируется постсинаптический потенциал, служит Са2+-зависимый экзоцитоз. Деполяризация нервного окончания, которая увеличивает проницаемость пресинаптической мембраны для Са2+, — необходимое условие выделения ацетилхолина.
Ацетилхолин химически нестоек, в щелочной среде быстро распадается на холин и уксусную кислоту. Разрушение его в холинергическом синапсе катализируется ферментом ацетилхолинэстера- зой, открытым О. Леви. Ацетилхолинэстераза находится на постсинаптической мембране рядом с холинорецептором и является одним из самых быстродействующих ферментов. Быстрое разрушение медиатора обеспечивает лабильность холинергической нервной передачи. Образовавшийся холин захватывается белками-транспортерами пресинаптической мембраны и служит далее для восстановления ацетилхолина в терминали (рис. 6).

/>Рис. 6. Схема строения холинергического синапса (цит. по: Маркова И. Н., Неженцева М. Н., 1997):
АХ - ацетилхолин; ХР - холинорецептор; М - мускариновый холинорецептор; Н - никотиновый холинорецептор; АХЭ — ацетилхолинэстераза; ТМ — транспортный механизм; ХА — холинацетилтрансфераза; (+) — активация; (-) — торможение
Действие ацетилхолина на мембрану состоит в его реакции с хо- линорецепторами, входящими в структуру клеточной мембраны (рис. 7). Так, реакция ацетилхолина с Н-холинорецептором вызывает изменение пространственного расположения атомов белковой молекулы рецептора. В результате увеличивается размер межмоле- кулярных пор мембраны, образуя свободный проход для ионов Na+, а затем и К+, и происходит деполяризация клеточной мембраны с последующей реполяризацией. Вызываемые ацетилхолином изменения молекулы рецептора легко обратимы. После передачи импульса уже приблизительно через 1 мс заканчивается деполяризация и восстанавливается обычная проницаемость мембраны. К этому времени холинорецептор уже свободен от связи с ацетилхолином.
Полагают, что вызываемая ацетилхолином деформация молекулы рецептора приводит не только к увеличению межмолекулярных пор мембраны, но и способствует отторжению ацетилхолина от рецептора. Отторжение это необходимо для взаимодействия освобождающего ацетилхолина с ацетилхолинэстеразой и его последующего разрушения (см. рис. 7).
Вещества, влияющие на холинорецепторы, способны вызывать стимулирующий (холиномиметический) или угнетающий (холинолитический) эффект.

Рис. 7. Схема взаимодействия ацетилхолина с холинорецептором
и ацетилхолинэстеразой (цит. по: Закусов В. В., 1973):
ХР — холинорецептор; АХЭ — ацетилхолинэстераза; А — анодный центр ХР и АХЭ; Э — эстеразный центр АХЭ и эстерофильный центр ХР
Фармакологические вещества могут воздействовать на следующие этапы синаптической передачи холинергических синапсов: синтез ацетилхолина; 2) процесс высвобождения медиатора; 3) взаимодействие ацетилхолина с холинорецепторами; 4) разрушение ацетилхолина; 5) захват пресинаптическим окончанием холина, образующегося при разрушении ацетилхолина. Например, на уровне пресинаптических окончаний действует ботулиновый токсин, препятствующий высвобождению медиатора. Транспорт холина через пресинаптическую мембрану (нейрональный захват) угнетается гемихолином. Непосредственное влияние на холинорецепторы оказывают холиномиметики (пилокарпин, цитизин) и холинолитики (М- холиноблокаторы, ганглиоблокаторы и периферические миорелак- санты). Для угнетения фермента ацетилхолинэстеразы могут быть использованы антихолинэстеразные средства (прозерин).
Ацетилхолин (Acetylcholinum), сокращенно АЦХ (ACh) – быстродействующий нейротрансмиттер в нервно-мышечном соединении и в вегетативных ганглиях, обеспечивающий передачу сигналов между нейронами. Нейромедиатор изменяет состояние нейронных сетей по всему мозгу, выполняя классическую функцию нейромодулятора. Рассматривается в качестве основного возбуждающего нейротрансмиттера на периферии.Ацетилхолин – первый обнаруженный учеными нейротрансмиттер, а также самый распространенный нейромедиатор в организме.

Функции ацетилхолина
Разнообразные эффекты ацетилхолина зависят от места его высвобождения, подтипов рецепторов и целевых функций нейронов. Однако обобщенное действие нейротрансмиттера состоит в том, что он стимулирует поведение, адаптивное к актуальным стимулам окружающей среды, и снижает реакцию на текущие раздражители, не требующие немедленных действий. Предполагается, что АЦХ является критически важным для реакции на неопределенность, улучшая соотношение сигнал/шум в процессе обучения.
В центральной нервной системе ацетилхолин выполняет стабилизирующие функции: как возбуждающую, так и угнетающую (ингибирующую). Он приводит мозг в состояние бодрствования, когда требуются энергичные действия, и, наоборот, замедляет передачу импульсов, когда нужно действовать сосредоточенно. Действие ацетилхолина обеспечивают два вида рецепторов: никотиновые – ускорение, а мускариновые – торможение.
В ЦНС основная роль ацетилхолина – возбуждающая. Он активирует сенсорные системы при пробуждении, помогает поддерживать быструю фаза сна – REM-сон. Ацетилхолин играет важную роль в процессе обучения и запоминания информации. Этот нейромедиатор принимает участие в процессах логического мышления, отвечает за концентрацию внимания. Также ацетилхолин обеспечивает синаптогенез – продукцию новых синапсов (связей) между нейронами.
Активная когнитивная деятельность, например, при внимательном изучении важного материала, приводит к повышенному синтезу ацетилхолина. Продолжительное интеллектуальное бездействие мозга стимулирует разрушение нейромедиатора специальным ферментом – ацетилхолинэстеразой.
Дефицит ацетилхолина
Холинергическая система – часть головного мозга, в которой производится ацетилхолин. По одной из версий, повреждение этой зоны связано с развитием сенильной деменции альцгеймеровского типа. Холинергические эффекты обуславливают старт нейродегенеративных процессов, проявляющихся расстройством кратковременной и долговременной памяти, нарушениями речевой и когнитивных функций, утратой способности ориентироваться в обстановке, невозможностью самообслуживания.
Нехватка ацетилхолина также играет роль в развитии идиопатического синдрома паркинсонизма. Ацетилхолин взаимодействует с нейромедиатором дофамином, обеспечивая плавность движений. Когда существует дисбаланс между ацетилхолином и дофамином, движения человека становятся неточными, возникает:
- ограничение темпа и объема движений;
- непроизвольные быстрые ритмичные колебательные движения в разных частях тела;
- постуральная неустойчивость – невозможность сохранять положение тела;
- отсутствие мышечных реакций на стимулы.
В периферической нервной системе нейромедиатор обеспечивает сокращение сердечной и скелетных мышц. Деструкция ацетилхолиновых рецепторов факторами гуморального и клеточного иммунитета провоцирует развитие миастении – аутоиммунного заболевания, проявляющегося эпизодами мышечной слабости и быстрой утомляемости. Болезнь возникает вследствие аутоиммунной атаки на постсинаптические ацетилхолиновые рецепторы, результатом чего является нарушение нервно-мышечной передачи сигналов. В лечении миастении используют лекарственные вещества, ингибирующие (тормозящие) активность холинэстеразы (фермента, расщепляющего ацетилхолин). Применение этих средств усиливает эффекты ацетилхолина на железы внутренней секреции, сердце, нервные узлы, гладкую и скелетные мышцы.
Доказана роль ацетилхолина в формировании табачной зависимости. Когда никотин достигает структур головного мозга, то активизируются рецепторы ацетилхолина. Со временем мозг привыкает, что никотина, похожего на ацетилхолин, теперь в избытке. Соответственно, раз этого вещества много, то необходимо приспособить процесс синтеза медиаторов к новой реальности. Чтобы это откорректировать, нейроны головного мозга сокращают производство ацетилхолина. Когда собственных запасов ацетилхолина перестает хватать, курильщик будет тянуться к сигарете,чтобы взбодриться, сосредоточиться, успокоиться. То есть у курящего человека функциональные обязанности ацетилхолина переданы никотину.
Как увеличить уровень ацетилхолина в организме
Предшественником ацетилхолина, то есть строительным блоком, на основе которого синтезируется эта биологическая субстанция, является органическое соединение – витаминоподобное вещество холин. В организме производство ацетилхолина происходит при воздействии фермента холин-ацетилтрансферазы. Соответственно, чтобы вырабатывался и поддерживался достаточный уровень ацетилхолина, в рационе должно присутствовать достаточное количество продуктов, содержащих холин.
Умеренное потребление холина улучшает когнитивные функции, предотвращает возрастное ухудшение памяти. Однако, как показывают исследования, современники испытывают дефицит этого витаминоподобного вещества, ранее именуемого витамин В4. Рекомендуемое суточное количество холина составляет 425 мг для лиц женского пола, 550 мг для особ мужского пола.
Перечень продуктов, содержащих холин:
- яичный желток;
- субпродукты (печень и почки);
- свинина, говядина;
- рыба;
- бобовые культуры;
- крестоцветные овощи (брокколи, цветная, брюссельская капуста, редька, кольраби);
- цельные зерна;
- творог, твердые сыры;
- орехи.
При необходимости холин может приниматься в качестве пищевой добавки, зарегистрированной под маркером E1001. Однако высокие дозы могут вызвать побочные эффекты, такие как тошнота, рвота, аллергические реакции.
В некоторых ситуациях, например, при дегенеративном психоорганическом синдроме, деменциях, после черепно-мозговых травм требуется внутримышечное или внутривенное введение холина в форме холина альфосцерата (Cholini alfosceras). Препарат положительно влияет на познавательные и поведенческие реакции, усиливает концентрацию внимания, улучшает процесс запоминания и воспроизведения информации. Средство улучшает настроение, устраняет эмоциональную лабильность, раздражительность и апатию.

Избыток ацетилхолина
Накопление ацетилхолина в организме может инициировать головную боль, мышечную дисфункцию, серьезные психоэмоциональные нарушения.
При стимуляции мотонейронов и вегетативных нейронов происходит выброс ацетилхолина в узкое пространство – синаптическую щель. Благодаря этому передается импульс к мышце или внутреннему органу. В нормальной ситуации после передачи импульса биологический передатчик расщепляется ферментом ацетилхолинэстеразой. Соответственно процесс передачи сигнала завершается. Однако, если каким-то образом подавляется активность ацетилхолинэстеразы, то возникает избыток ацетилхолина. Такое наблюдается при воздействии на организм человека нервнопаралитическими газами или пестицидами.
Приведем пример. Зарин – отравляющее вещество нервнопаралитического действия – ингибирует ацетилхолинэстеразу, в результате чего увеличивается содержание ацетилхолина в синаптической щели. Соответственно, активирующие сигналы передаются непрерывно. Постоянно поступающие импульсы поддерживают гиперактивность органов, иннервируемых вегетативными и двигательными нейронами. Это приводит к их полному истощению.
Фосфорорганические соединения также входят в состав инсектицидов. Воздействие на организм человека химическими веществами, используемыми в сельском хозяйстве, приводит к накоплению ацетилхолина в нервной системе.
Избыток нейромедиатора вызывает опасные для жизни состояния. Симптомы (по нарастающей):
- выделения из носа;
- сужение зрачков;
- затруднение дыхания, бронхоспазм;
- тошнота и рвота;
- усиленное слюноотделение;
- непроизвольное мочеиспускание и дефекация;
- подергивание мышц и конвульсии;
- коматозное состояние (паралич, остановка дыхания и сердца).
Организм подвергается воздействию этих химических веществ через кожный покров, при вдыхании или при проглатывании. Отравление чаще всего возникает при контакте с пестицидами при обработке сельскохозяйственных культур, или при потреблении продуктов, на которые попали капли химических реагентов, используемых в борьбе с тараканами.
Заключение
Ацетилхолин – важный медиатор нервной системы. Дефицит нейротрансмиттера провоцирует тяжелые неврологические дефекты в рамках деменции альцгеймеровского типа и болезни Паркинсона. Чрезмерное возбуждение холинергических рецепторов чревато дисфункцией вегетативной нервной системы, вплоть до коматозного состояния. Здоровое сбалансированное питание, соблюдение мер безопасности при работе с химическими соединениями – залог поддержания адекватного уровня ацетилхолина в организме.
Содержание
Ацетилхолин (АХ) — медиатор в постганглионарных синапсах — накапливается в высокой концентрации в везикулах аксоплазмы нервного окончания. АХ образуется из холина и активированной уксусной кислоты (ацетилкофермент А) под действием фермента ацетилхолинтрансферазы. Высокополярный холин активно захватывается аксоплазмой. На мембране холинергического аксона и нервных окончаний имеется специальная транспортная система. Механизм высвобождения медиатора до конца не раскрыт. Везикулы закреплены в цитоскелете при помощи белка синапсина таким образом, что их концентрация около пресинаптической мембраны высокая, однако контакт с мембраной отсутствует. При возникновении возбуждения повышается концентрация Са2+ в аксоплазме, активируются протеинкиназы, и происходит фосфорилирование синапсина, приводящее к отсоединению везикул и связыванию их с пресинаптической мембраной. Затем содержимое везикул выбрасывается в синаптическую щель. Ацетилхолин мгновенно проходит сквозь синаптическую щель (молекула АХ имеет длину около 0,5 нм, а ширина щели составляет 30-40 нм). На постсинаптической мембране, т. е. мембране целевого органа, АХ взаимодействует с рецепторами. Эти рецепторы возбуждаются также алкалоидом мускарином и поэтому называются мускариновыми ацетилхолиновыми рецепторами (М-холинорецепторы). Никотин имитирует действие ацетилхолина на рецепторы ганглионарных синапсов и концевой пластинки. Никотин возбуждает холинорецепторы ганглионарных синапсов и концевой пластинки мотонейрона, поэтому этот тип рецепторов назван никотиновыми ацетилхолиновыми рецепторами (N-холинорецепторы).
В синаптической щели ацетилхолин быстро инактивируется специфической ацетилхолинэстеразой, находящейся в щели, а также менее специфической сывороточной холинэстеразой (бутирилхолинэстеразой), находящейся в сыворотке крови и интерстициальной жидкости.
По своему строению, способу передачи сигнала и сродству к различным лигандам М-холинорецепторы подразделяются на несколько типов. Рассмотрим M1, М2- и М3-рецепторы. M1-Рецепторы находятся на нервных клетках, например ганглиях, и их активация способствует переходу возбуждения с первого на второй нейрон. М2-Рецепторы расположены в сердце: открытие калиевых каналов приводит к замедлению диастолической деполяризации и уменьшению частоты сердечных сокращений. М3-Рецепторы играют роль в поддержании тонуса гладких мышц, например, кишечника и бронхов. Возбуждение этих рецепторов приводит к активации фосфолипазы С, деполяризации мембраны и повышению тонуса мышц. М3-Рецепторы расположены также в клетках желез, которые активируются посредством фосфолипазы С. В головном мозге имеются разные типы М-холинорецепторов, играющие роль во многих функциях: передаче возбуждения, памяти, обучаемости, болевой чувствительности, контроле активности ствола мозга. Активация М3-рецепторов в эндотелии сосудов может приводить к высвобождению оксида азота N0 и таким образом расширять сосуды.
Ацетилхолин (лат. Acetylcholinum) — медиатор нервной системы, биогенный амин, относящийся к веществам, образующимся в организме.
Ацетилхолину принадлежит важная роль как медиатору центральной нервной системы. Он участвует в передаче импульсов в разных отделах мозга, при этом малые концентрации облегчают, а большие — тормозят синаптическую передачу. Изменения в обмене ацетилхолина могут привести к нарушению функций мозга.
Ацетилхолин является посредником передачи нервного импульса к мышце. При недостатке ацетилхолина снижается сила сокращений мышц.
Окончания нервных волокон, для которых он служит медиатором, называются холинергическими, а рецепторы, взаимодействующие с ним, называют холинорецепторами. Холинорецепторы постганглионарных холинергических нервов (сердца, гладких мышц, желез) обозначают как м-холинорецепторы (мускариночувствительные), а расположенные в области ганглионарных синапсов и в соматических нервномышечных синапсах — как н-холинорецепторы (никотиночувствительнные). Такое деление связано с особенностями реакций, возникающих при взаимодействии ацетилхолина с этими биохимическими системами: мускариноподобных в первом случае и никотиноподобных — во втором; м- и н-холинорецепторы находятся также в разных отделах ЦНС.
При микроэлектродной регистрации электрических потенциалов постсинаптической мембраны нервно-мышечного синапса Фетт и Катц (Fatt and Katz, 1952) выявили спонтанные небольшие (0,1—3 мВ) деполяризующие потенциалы, возникающие случайным образом примерно 1 раз в секунду. Авторы назвали эти потенциалы миниатюрными потенциалами концевой пластинки. Их амплитуда была существенно ниже пороговой для развития потенциала действия. Они увеличивались под действием ингибитора АХЭ неостигмина и блокировались тубокурарином (конкурентным блокатором N-холинорецепторов); следовательно, они были обусловлены выделением ацетилхолина. В связи с этим было высказано предположение, что ацетилхолин выделяется из пресинаптических окончаний дробными постоянными порциями — квантами. Вскоре был обнаружен и морфологический субстрат квантов — синаптические пузырьки (De Robertis and Bennett, 1955). Когда в окончание аксона мотонейрона приходит потенциал действия, выделяется 100 и более квантов (пузырьков) ацетилхолина (Katz and Miledi, 1965). Закономерности хранения и выделения ацетилхолина, изученные на нервно-мышечном синапсе, применимы и к другим холинергическим синапсам с быстрой передачей.
Предполагается, что в каждом пузырьке содержится от 1000 до 50 000 молекул ацетилхолина, а в пресинаптическом окончании мотонейрона содержится 300 000 и более пузырьков. Кроме того, не исключено, что достаточно существенное количество ацетилхолина диффузно растворено в аксоплазме. Запись токов одиночных каналов постсинаптической мембраны нервно-мышечного синапса при постоянной аппликации ацетилхолина показала, что одна молекула этого медиатора вызывает потенциал порядка 3 х 10”7 В. Из этого следует, что даже минимальное (по расчетам) количество ацетилхолина в одном пузырьке — 1000 молекул — достаточно для того, чтобы вызвать миниатюрный потенциал концевой пластинки (Katz and Miledi, 1972).
Экзоцитоз ацетилхолина и других медиаторов из пресинаптических окончаний подавляется ботулотоксином и столбнячным токсином — ядами Clostridium botulinum и Clostridium tetani соответственно. Этими анаэробными спорообразующими организмами вырабатываются одни из самых сильных из известных токсинов (Shapiro et а. 1998). Токсины Clostridium, состоящие из связанных дисульфидными мостиками тяжелой и легкой цепей, соединяются с неизвестным пока рецептором на холинергическом окончании и затем посредством эндоцитоза переносятся в цитозоль. Легкая цепь представляет собой цинксодержащую эндопептидазу, которая после активации гидролизует компоненты ядра комплекса SNARE, участвующего в экзоцитозе. Различные типы ботулотоксина разрушают разные белки пресинаптической мембраны (синтаксин-1 и SNAP-25) и синаптических пузырьков (синаптобревин). Ботулотоксин А как лекарственное средство рассматривается в гл. 9 и 66.
Столбнячный токсин — это яд центрального действия: он ретроградно переносится по аксонам мотонейронов в тела этих нейронов в спинном мозге, далее переходит в связанные с мотонейронами тормозные нейроны и блокирует экзоцитоз медиатора из последних. Именно это и приводит к характерным для столбняка судорогам. Яд паука черная вдова — а-латротоксин — связывается с трансмембранными белками пресинаптических окончаний нейрексинами, вызывая массивный экзоцитоз синаптических пузырьков (Schiavo et al., 2000).
Читайте также:
