
Биоэлектрических явлениях в нервах и мышцах
Возникновение и распространение возбуждения сопровождаются электрическими явлениями в тканях.
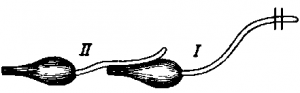
Рис. СХЕМА ОПЫТА ВТОРИЧНОГО СОКРАЩЕНИЯ (ОБЪЯСНЕНИЕ В ТЕКСТЕ).
Возражения, которые выдвигал Вольта, основывались на доказанном им факте, что при соединении двух разных металлов, в опыте Гальвани — меди и железа, возникает разность потенциалов, которая и вызывает сокращение мышцы.
Начался интересный научный спор между Гальвани и Вольта. По мере развертывания этого спора каждая из сторон прибегала к новым опытам для доказательства правоты своих взглядов. Решающим был опыт Гальвани, который он проводил без участия металлов. Этот опыт, получивший название второго опыта Гальвани, или сокращения без металлов, заключался в следующем: у лягушки отпрепаровывали седалищный нерв и набрасывали на обнаженные мышцы голени; при этом мышцы сокращались.
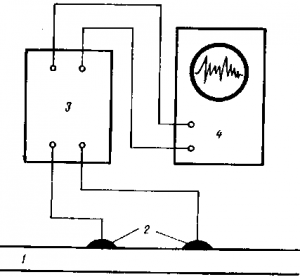
Рис. 2 Схема осциллографа для записи биопотенциалов, 1 — возбудимая ткань, 2 — электроды, 3- усилитель, 4- осцилограф
Особенно интересными были опыты, поставленные в середине прошлого века и получившие название вторичного тетануса, или вторичного сокращения. На мышцу одного нервно-мышечного препарата накладывали нерв другого нервно-мышечного препарата (рис. ). При раздражении индукционным током нерва первого нервно-мышечного препарата сокращалась и мышца второго препарата, нерв которого был наброшен на мышцу первого препарата. Это явление объяснялось тем, что при возбуждении в мышце возникали биоэлектрические явления, которые вызывали возбуждение второго нервно-мышечного препарата.
Исследования, проведенные уже во второй половине XIX века и в XX веке с использованием новейших физических приборов, внесли очень много в учение о биоэлектрических явлениях.
Очень ценные факты были получены крупнейшим русским физиологом Н Е. Введенским при помощи разработанной им методики телефонического выслушивания токов действия мышц и нервов. Выслушивание в телефонную трубку мышц и нервов при их возбуждении давало возможность улавливать особые звуки, связанные с возникновением токов действия и их ритмами.
Детальное изучение биоэлектрических явлений стало возможным благодаря тому, что был сконструирован и введен в физиологическую практику струнный гальванометр. Это настолько чувствительный прибор, что при его помощи удается обнаружить и зарегистрировать весьма малые электрические колебания в живых тканях.
Опишем схематически устройство гальванометра. Между двумя сильными электромагнитами протянута очень тонкая платиновая или кварцевая нить, покрытая золотом или серебром. Концы этой нити (струны) соединяются с электродами, которые прикладывают к нерву или мышце. Когда по струне проходит электрический ток, она отклоняется к одному из электромагнитов. Гальванометр устроен так, что при помощи специальных осветителей, струна освещается и ее движения фотографируются на фотографической бумаге. Колебания струны, сфотографированные на бумаге, дают возможность более детально изучить особенности биоэлектрических явлений. Гальванометр позволяет изучить весьма слабые токи. Струна гальванометра в новейших приборах может совершить до 1000 колебаний в секунду.
В настоящее время пользуются осциллографами разной конструкции, которые дают возможность еще более детально изучить биоэлектрические явления в живых тканях.
Наиболее чувствительным и не обладающим инертностью прибором является катодный осциллограф. Принцип устройства этого прибора заключается в том, что при помощи спе циального приспособления к так называемой катодной лампе пропускается тонкий пучок электронов. Этот пучок проходит между двумя металлическими пластинками, которые соединены с нервом или мышцей. Разности потенциалов, которые появляются в исследуемой живой ткани, передаются металлическим пластинкам, и катодный луч отклоняется к той пластинке, которая имеет положительный электрический заряд. Отклонение луча за писывается на фотографической пленке, которая непрерывно движется. Полученные таким образом осциллограммы дают возможность изучать малейшие колебания электрических зарядов в исследуемой ткани.
При записи биотоков, которые очень малы, между объектом и осциллографом включаются усилители (рис. 2), что дает возможность записать биопотенциалы в тысячные доли вольта и продолжительностью в тысячные доли секунды.
При записи биопотенциалов мозга — электроэнцефалограммы, электрокардиограммы и др., часто пользуются чернильнопишущим приспособлением, что создает возможность записать колебания биопотенциалов чернилами на бумаге.
Статья на тему Биоэлектрические явления в тканях
БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ — процессы распределения и транспорта электрических зарядов в живом организме, обусловленные присутствием в живых клетках и тканях большого количества фиксированных (заряженные группы био-макромолекул) и подвижных (свободные ионы и электроны) электрических зарядов. Многие вещества, поступающие в организм и выводимые из него в процессе метаболизма, находятся в виде ионов. Перераспределение и транспорт этих зарядов служат причиной возникновения биоэлектрических потенциалов (см.). Наиболее крутой градиент ионов обнаруживается на клеточных мембранах (см. Мембраны биологические), которые способны эффективно регулировать неравномерное и избирательное распределение ионов. У большинства клеток мембранный потенциал, называемый потенциалом покоя, зависит от градиента ионов калия. Его величина может быть рассчитана по уравнению Нернста:

В возбудимых образованиях (напр., нервные и мышечные клетки) регистрируется особый вид мембранного потенциала — так наз. потенциал действия, который, в отличие от потенциала покоя, представляет собой пиковый потенциал, передвигающийся в форме волны возбуждения по поверхности клетки со скоростью до нескольких десятков метров в секунду. В возбужденном участке потенциал приобретает обратный знак (т. е. потенциал реверсируется) и внутренняя поверхность заряжается положительно, а наружная — отрицательно. Абсолютная величина потенциала, как правило, превышает уровень потенциала покоя. Согласно ионной теории Ходжкина (A. L. Hodgkin, 1958) возникновение потенциала действия обусловлено избирательным увеличением проницаемости мембраны для ионов натрия.
Основные биоэнергетические процессы в живом организме связаны с биологическим окислением (см. Окисление биологическое), в процессе к-рого электроны движутся по цепочке дыхательных ферментов, расположенных во внутренней мембране митохондрий, по градиенту окислительно-восстановительного потенциала. В соответствии с хемиосмотической гипотезой Митчелла (P. Mitchell, 1966) перенос электронов по дыхательной цепи, звенья которой располагаются поперек мембраны, сопровождается появлением мембранного потенциала.
Живой организм является не только генератором биоэлектричества, но и пассивным проводником электрического тока, возникающего в организме или приложенного извне (см. Электропроводность биологических систем). Электропроводность живых клеток и тканей зависит от импеданса, т. е. суммы активного (омического) и реактивного (емкостного) сопротивлений. Прохождение постоянного электрического тока через живые клетки сопровождается поляризацией, к-рая может быть рассчитана по формуле:
где I — сила тока, R — электрическое сопротивление, U — напряжение тока, P — напряжение поляризации (см. Поляризация). При пропускании переменного тока напряжение поляризации уменьшается с возрастанием частоты. Это явление носит название дисперсии электропроводности (см. Дисперсия). Б. Н. Тарусовым в 1938 г. было показано, что дисперсия электропроводности может служить чувствительным индикатором жизнеспособности клеток и тканей.
Как правило, поверхность живых клеток и тканей несет значительный отрицательный заряд, т. к. ионогенные группы на поверхности клеток, представленные концевыми группами макромолекул клеточной поверхности, при физиологических значениях pH ионизованы. Под действием сил электростатического притяжения положительные ионы из раствора будут притягиваться к поверхности и создавать двойной электрический слой. Часть энергии двойного слоя можно измерить по скорости движения заряженной клетки в электрическом поле. Этот так наз. электрокинетический потенциал (см. Электрокинетические явления) рассчитывается по уравнению Смолуховского: Z = 4πηu/DE, где D — диэлектрическая постоянная среды, η — коэффициент вязкости, E — градиент напряжения, u — электрофоретическая подвижность. В наст, время электрокинетический потенциал используют как индикатор состояния клеточной поверхности. В частности, было показано, что подвижность асцитных клеток несколько ниже по сравнению с нормальными клетками.
К электрокинетическим явлениям относят также потенциал течения. В живом организме имеется постоянное течение жидкости. Всякое движение жидкости (напр., течение крови по артериям и венам) сопровождается появлением потенциала течения между жидкой фазой и стенкой сосуда. Поскольку удельная электропроводность крови велика, эти потенциалы малы — не больше 1 —2 мв. Изменение этого потенциала, вероятно, может иметь и диагностическое значение.
К электрокинетическим явлениям относится и электроосмос, который наблюдается при движении жидкости через пористую перегородку под влиянием внешнего электрического поля. В ряде случаев положительный и отрицательный аномальный осмос в почках объясняется электроосмотическим переносом воды.
Библиография: Катц Б. Нерв, мышца и синапс, пер. с англ., М., 1968, библиогр.; Курелла Г. А. Биоэлектрические потенциалы, в кн.: Биофизика, под ред. Б. Н. Тарусова и О. Р. Колье, с. 238, М., 1968, библиогр.; Скулачев В. П. Трансформация энергии в биомембранах, М., 1972, библиогр.
При определенных условиях в нервах и мышцах возникают электрические токи. Различают: 1) токи, или потенциалы, покоя и повреждения и 2) токи, или потенциал, действия – акционные точки.
Токи покоя
При отсутствии раздражения и, следовательно, возбуждения, существует разность потенциалов между наружной поверхностью цитоплазмы живой клетки (мембраной) и её внутренним содержимым.
Поверхностный слой цитоплазмы живой клетки, находиться в состоянии покоя, имеет положительный электрический заряд, а цитоплазма внутри клетки заряжена электроотрицательно. Поэтому, введя в клетку микроэлектрод и приложив другой электрод к поверхности клетки, можно отвести ток к регистрирующему прибору и измерить потенциал покоя, мембранный или клеточный потенциал.
Токи покоя, или клеточные потенциалы, обнаруживаются в неповрежденных живых клетках. Современными методами отведения электрического тока и его регистрации установлено, что токи покоя отдельных клеток животных, например гигантских нервных волокон кальмара, — 50-60 мв (милливольт), миелиновых нервных волокон амфибий – 70 мв, теплокровных – 90 мв, большинства тел нейронов – 80-100 мв, поперечнополосатых мышц амфибий – 80 мв, эпителия амфибий – 20 мв, волокон сердечной мышцы животных с постоянной температурой тела – 95 мв.
Токи повреждения
При повреждении живой ткани открывается доступ к внутреннему содержимому клеток, которое заряжено электроотрицательно. Поэтому если приложить электрод к неповрежденному участку ткани, находящемуся в состоянии покоя и, следовательно, заряженному электроположительно, а другой электрод приложить к поврежденному или перерожденному участку, заряженному электроотрицательно, то при соединении обоих электродов проводником тока с регистрирующим прибором можно измерить разность потенциалов.
Токи повреждения можно обнаружить на нервно-мышечном препарате. Если разрезать мышцу или любым способом повредить ее в одном участке (прижечь, раздавить и т. д.) и затем, приложив неполяризующиеся электроды, соединить срез мышцы или участок повреждения с целой поверхностью мышцы, не подвергающейся раздражению, то от целой поверхности к участку разреза или повреждения направится электрический ток. Сильно поврежденный участок всегда электроотрицателен по отношению к электроположительному неповрежденному участку. Следовательно, биоэлектрический ток повреждения, или альтерационный ток, возникает на границе целой и поврежденной частей ткани. Он обнаруживается и на некотором расстоянии от места повреждения, уменьшаясь с увеличением расстояния от места повреждения. Поэтому по месту происхождения его называют также демаркационным током. Однако ток повреждения появляется только тогда, когда отрицательно заряженный поврежденный участок соединяется проводником тока с положительно заряженным целым участком.
Величина тока повреждения меньше, чем клеточного потенциала, так как межклеточная и внеклеточная жидкости производят шунтирование (укорочение) при его измерении.
Разность потенциалов между целым и поврежденным участками скелетных мышц лягушки сразу же после повреждения равна 25-50 мв, наибольшая разность доходит до 80. В скелетных мышцах кролика токи повреждения равны 40 мв. В гладких мышцах токи повреждения меньше и быстрее уменьшаются до нуля. Разность потенциалов между целым и поврежденным участками седалищного нерва лягушки достигает 20-30 мв, седалищного нерва лошади — 6-16 мв, а седалищного нерва человека и обезьяны — 6-7 мв. Следовательно, токи повреждения нервов меньше, чем токи повреждения мышц. Особенно малы токи повреждения нервов человека и высших животных. Постепенно разность потенциалов уменьшается.
Когда нерв второго нервно-мышечного препарата набрасывается на первый нервно-мышечный препарат таким образом, чтобы он одновременно коснулся неповрежденного поврежденного участков мышцы, электрическая цепь замыкается. При этом мышца второго препарата сокращается в результате раздражения нерва током, возникающим в момент его замыкания между положительно заряженным нормальным участком мышцы первого препарата и отрицательно заряженным поврежденным ее участком.
Токи действия
Акционные токи, или токи (потенциалы) действия, возникают в раздражаемых тканях. Их появление характеризует возбуждение.
Для обнаружения тока действия нужно приложить один электрод к неповрежденному участку живой ткани, который находится в состоящих покоя и поэтому заряжен электроположительно, а другой — к раздражаемому участку и присоединить оба электрода к регистрирующему прибору. Раздражаемый участок при приложении к нему раздражителя немедленно или через несколько сотых миллисекунды становится электроотрицательным. Это падение потенциала’ доходит до максимума, а затем данный участок снова становится электроположительным. Различают однофазные и двухфазные токи действия.
При прикладывании одного из неполяризующихся электродов к неповрежденному участку нерва или мышцы, а другого — к поврежденному участку возникает электрический ток. Этот ток регистрируется при включении в цепь гальванометра или другого более чувствительною прибора.
Стрелка гальванометра отклонится в одну сторону вследствие электроотрицательного заряда поврежденного участка и электроположительного заряда нормального участка. Но если нанести раздражение по соседству с нормальным участком, то волна возбуждения, которая возникнет в месте раздражения, на некоторое время превратит заряд под нормальным участком в электроотрицательный. В этот момент тока не будет, так как под обоими электродами заряд станет электроотрицательным и стрелка гальванометра станет на нулевую линию. Когда волна возбуждения продвинется дальше и будет находиться между электродами, нормальный участок снова станет электроположительным и стрелка гальванометра снова отклонится в ту же сторону.
Когда волна возбуждения дойдет до поврежденного участка, положение стрелки гальванометра не изменится, так как под вторым электродом в месте повреждения уже имеется электроотрицательный заряд. Таким образом, стрелка гальванометра движется только в одну сторону, а запись этого движения регистрирует однофазный, или монофазный ток действия.
Это явление Дюбуа-Реймон назвал отрицательным колебанием тока покоя, которое возникает каждый раз, когда волна возбуждения проходит под электродом, расположенным на неповрежденном участке ткани.
Однофазный ток действия можно зарегистрировать также при раздражении ткани во время отведения клеточного потенциала. При внутриклеточном введении одного электрода и приложении другого электрода к поверхности клетки установлено, что клеточный потенциал не только исчезает в течение тысячных долей секунды, но что в течение этого промежутка времени амплитуда тока действия на 30-50 мв больше клеточного потенциала. Это объясняется тем, что в пункте прохождения волны возбуждения наружная поверхность мембраны становится электроотрицательной, а внутренняя — электроположительной. Продолжительность тока действия в нервных волокнах и клетках скелетных мышц составляет 0,1-5 мс.
Однофазный ток действия представляет собой местный электроотрицательный потенциал, мгновенно возникающий при очень слабых, подпороговых одиночных раздражениях и ограниченный несколькими миллиметрами от пункта раздражения. Величина местного низковольтного электроотрицательного заряда убывает по мере удаления от пункта раздражения. Чем больше подпороговое раздражение, тем больше возникающее возбуждение и амплитуда или отклонение однофазного тока действия.
Следовательно, местное возбуждение или местный сдвиг обмена веществ отличается градуальностью — возрастанием возбуждения по мере увеличения силы раздражения. Местное возбуждение наступает сразу после раздражения, т. е. без латентного периода. В участке слабого местного нераспространяющегося возбуждения возбудимость сохраняется, рефрактерность отсутствует. Если бы при местном возбуждении наблюдалась рефрактерность, то оно не могло бы перейти в пороговое, распространяющееся возбуждение.
Местный однофазный ток действия предшествует распространяющейся волне возбуждения и подготавливает ее. При одиночном пороговом раздражении волна возбуждения возрастает, достигает критического уровня и превращается из местной в распространяющуюся. Поэтому распространяющееся возбуждение наступает после латентного периода, продолжительность которого равна длительности местного нераспространяющегося возбуждения. В отличие от местного возбуждения распространяющееся возбуждение не градуально и сопровождается фазами изменения возбудимости. Потенциал действия, возникший в раздражаемом участке мембраны, вызывает возбуждение соседних участков мембраны и таким образом возбуждение распространяется по нервным и мышечным волокнам.
О прохождении волны возбуждения по возбудимой ткани судят прежде всего по токам действия. Однофазное колебание тока действия распространяющейся волны возбуждения в отдельных нервных и мышечных клетках состоит из нескольких составляющих: 1) высоковольтного потенциала, пика, 2) отрицательного следового потенциала и 3) положительного следового потенциала.
По сравнению со следовыми потенциалами пик имеет наибольшую высоту и наименьшую продолжительность. Пик имеет приблизительно одинаковую высоту на всем протяжении возбудимых тканей животных с постоянной температурой тела. Это указывает на то, что по мере распространения возбуждения вольтаж потенциала не падает. Этот факт свидетельствует, что химические вещества, необходимые для возникновения возбуждения, равномерно распределены по возбудимой ткани и что распространение возбуждения происходит самостоятельно, автоматически. Раздражение при достаточной интенсивности вызывает возбуждение только в том участке, который раздражается, а дальше возбуждение движется по ткани, вовлекая в сдвиг обмена веществ вещества, имеющиеся в возбудимой ткани.
Пик играет ведущую роль в распространении возбуждения. Высота пика различна у нервных волокон разных групп, имеющих неодинаковый диаметр, и изменяется в зависимости от физиологического состояния волокна и скорости проведения возбуждения. Чем толще нервное миелиновое волокно, тем меньше длительность пика. По мышечному волокну человека возбуждение проходит за 4-5 м/с.
В волокнах скелетных мышц однофазные токи действия различаются по амплитуде и по продолжительности. В волокнах сердечной мышцы они отличаются большей продолжительностью пика, который после начального крутого снижения начинает снижаться постепенно, а затем снова круто падает.
Токи действия в гигантских нервных волокнах кальмара достигают 90-110 мв, в миелиновых нервных волокнах амфибий 110 мв, в волокнах поперечнополосатых мышц амфибий — 120 мв, а в волокнах сердечной мышцы животных с постоянной темпера турой тела — 135 мв. Чем больше частота тока действия, тем меньше его вольтаж. Гладкие мышечные волокна отличаются тем, что их токи действия не превосходят мембранных потенциалов (не больше 70-80 мв).
Условия, улучшающие обмен веществ (повышение темпера туры, отдых), уменьшают продолжительность волны возбуждения, а условия, ухудшающие обмен веществ (утомление), увеличивают ее. С понижением температуры амплитуда пика уменьшается, а продолжительность его возрастает.
При температуре тела продолжительность подъема высоковольтного потенциала нерва составляет примерно 1/3 а продолжительность снижения — 2/3 его общей продолжительности. Предполагается, что крутой подъем пика обусловлен быстрым движением ионов натрия внутрь клетки.
Отрицательный следовой потенциал соответствует остаточным сдвигам распределения ионов калия и натрия. В отличие от пика он неустойчив и изменяется от условий среды. Он достигает 0,05 потенциала пика. Положительный низковольтный следовой потенциал также связан с перемещением ионов и равен примерно 0,002 потенциала пика.
Тепло и отдых уменьшают, а охлаждение и истощение увеличивают продолжительность следовой электроотрицательности. В нерве, лишенном кислорода, при действии наркоза и солей калия следовая низковольтная электроотрицательность отсутствует, а начальная высоковольтная электроотрицательность сохраняется, и при известных концентрациях наркоза даже не наступает заметных изменений её величины. Это указывает на то, что пик может возникать и при отсутствии кислорода, а следовая электроотрицательность связана с восстановительными процессами, которые совершаются в ткани вслед за возбуждением, и для её появления необходим кислород. Нарушение обмена веществ, утомление увеличивают продолжительность следовых потенциалов. Чем больше лабильность, тем меньше выражены следовые потенциалы. В высоколабильных мякотных волокнах человека и высших животных они почти не проявляются.
Двухфазный ток действия
В естественных условиях одиночные волны возбуждения встречаются чрезвычайно редко; В организме в нервах и мышцах передаются серии волн возбуждения, взаимно влияющих друг на друга. Возбуждение, которое достигает порога или несколько превышает его, носит ритмический характер. В других случаях возбуждение в естественных условиях является градуальным, длительным, стойким, переходящим в торможение. В последнем случае при чрезмерно большой силе или частоте раздражения, превышающей меру лабильности, полностью угнетается высоковольтная электрическая активность.
Волновое распространяющееся возбуждение возникает из неволнового, градуального. Такая же эволюция возбуждения происходила в филогенезе. Представление о возникновении волнового возбуждения дает следующий простой опыт.
После приложения обоих неполяризующихся электродов к нормальным целым участкам возбудимой ткани и включения этой ткани в цепь гальванометра его стрелка останется на нуле, так как все участки нормальной ткани, находящейся в состоянии покоя, имеют одинаковый заряд. Но когда ткань раздражают у одного из электродов, то возникает возбуждение, и заряд ткани под этим электродом становится отрицательным. При этом стрелка гальванометра отклоняется в одну сторону. Передвигаясь далее, волна возбуждения оказывается между электродами, и в это время заряды под электродами вновь становятся одинаковыми и стрелка гальванометра возвращается к нулю. Но когда волна возбуждения доходит до второго электрода, то гальванометр опять отмечает разность потенциалов, так как под вторым электродом заряд становится отрицательным, а в то же самое время под первым электродом заряд положительный, так как под ним нег возбуждения. Теперь стрелка гальванометра отклоняется в противоположную сторону, а затем, после прекращения возбуждения, возвращается к нулю. Следовательно, когда по ткани проходит волна возбуждения, то стрелка гальванометра отклоняется сначала в одну сторону, потом в другую. Запись этого движения стрелки гальванометра регистрирует двухфазный ток действия.
Форма регистрации токов, или потенциалов, действия зависит от места возникновения возбуждения, от используемых приборов и от расположения электродов. В настоящее время их записывают посредством катодных осциллографов, не имеющих инерции, в отличие от гальванометров. Современные катодные осциллографы регистрируют биопотенциалы в миллионные доли вольта, продолжающиеся одну стомиллионную или даже миллиардную долю секунды. Эти токи отличаются сложностью, и их нередко расшифровывают при помощи электронных устройств.
Биопотенциалы каждой возбудимой ткани, например сердца, имеют характерный вид и изменяются в зависимости от ее функционального состояния. Записываются биопотенциалы кожи, эпителия, соединительной ткани, рецепторов, сетчатки глаза, скелетных мышц, пищеварительных желез, гладкой мускулатуры пищеварительного канала и других органов, разных отделов центральной нервной системы и даже одного нейрона. Первые исследования биопотенциалов больших полушарий головного мозга провели В. Я. Данилевский, Р. Кетон (1875), продолговатого мозга — И. М. Сеченов (1882).
Н. В. Введенский (1883, 1884) соединил мышцу проводами с телефонной трубкой и установил важнейшие законы, по которым протекает в ней возбуждение. Он же впервые применил телефон для изучения явлений возбуждения в нерве.
Процессы жизнедеятельности неразрывно связаны с различными формами биоэлектрических явлений. В частности, биоэлектрические явления сопровождают возникновение возбуждения и его проведение по нервным волокнам, сокращение волокон скелетных, гладких мышц, миокарда, а также процессы всасывания в желудочно-кишечном тракте, восприятия, вкуса, запаха, деятельность анализаторов и т. д.
Причиной возникновения биоэлектрических явлений являются постоянно происходящие в процессе жизнедеятельности перераспределение ионов на мембранах и их трансмембранный транспорт [4,с.21]. На практике биоэлектрические явления регистрируют как разность электрически потенциалов между двумя точками живой ткани. Например, с помощью микро электродов можно измерить разность потенциалов между наружной и внутренней сторонами оболочки (мембраны) клетки.
Та разность потенциалов называется потенциалом покоя, или мембранным потенциалом. Наличие его обусловлено неравномерным распределением ионов (в первую очередь ионов натрия и калия) между внутренним содержимым клетки (ее цитоплазмой) и окружающей клетку средой.
Величина мембранного потенциала различна, причем для каждого типа клетки свойственна строго определенная величина потенциала покоя, которая отражает интенсивность обменных процессов, протекающих в них.
При возбуждении клетки возникает так называемый потенциал действия, который в отличие от потенциала покоя распространяется в виде волны возбуждения по поверхности клетки со скоростью нескольких десятков метров в секунду.
Существуют и другие виды потенциалов, например, потенциал повреждения.
Биоэлектрические явления исторически возникли как способ более совершенной связи между отдельными образованьями многоклеточного организма.
Биоэлектрические явления в деятельности ЦНС животных и человека
Существует два типа общения между клетками. Первый связан с химическим взаимодействием, при котором вещество, продуцируемое одной клеткой, достигает другой, вызывая в ней ответную реакцию.
Такой способ общения не обеспечивает быстрой передачи информации. Более совершенный способ передачи информации и сигнализации осуществляется при помощи нервных импульсов, одним из компонентов которых являются биоэлектрические явления.
Особенно отчетливо он выражен в деятельности центральной нервной системы животных и человека. Живой организм является не только генератором биопотенциалов, но и проводником электрического тока, причем изменение степени электропроводности живых тканей в зависимости от их жизнедеятельности может служить показателем жизнеспособности клеток или тканей [8, с. 83].
К наиболее важным функциям, присущим всем живым существам, можно отнести следующие:
- самовоспроизведение: запись, хранение и использование генетической информации;
- энергообеспечение: получение энергии за счет внешних энергетических ресурсов;
- превращение веществ: использование химических соединений окружающей среды для образования компонентов своего тела;
Раздражимость: способность принимать и обрабатывать сигналы, поступающие из внешней и внутренней сред организма, и адекватно отвечать на эти сигналы.
Биоэнергетика - это отрасль функциональной биологии, изучающая превращение энергии внешних ресурсов в биологически полезную работу.
Липидный компонент мембран более или менее стандартен: это фосфолипиды, реже глико - и сульфолипиды.
Белок - это тот компонент, который определяет специфику мембраны.
Важнейшей функцией многих мембран является превращение одной формы энергии в другую. Эта функция осуществляется особыми белками, встроенными в особого типа мембрану, носящую название энергопреобразующей. Такая мембрана непроницаема для большинства веществ, которые имеются в растворах по обе стороны.
Любая мембрана, выполняющая энергетическую функцию, способна к превращению химической энергии или энергии света в электрическую энергию, а именно в трансмембранную разность электрических потенциалов или в энергию разности концентраций веществ, содержащихся в разделенных мембраной растворах.
Некоторые энергообразующие мембраны могут превращать электрическую энергию в механическую. Определенные ткани теплокровных животных и человека превращают накопленную энергию в тепло в целях терморегуляции, что оказывается биологически оправданным при понижении окружающей температуры.
Таким образом, человеческий организм является генератором биопотенциалов, проводником электрического тока.
Биоэлектрические явления особенно ярко выражены в деятельности центральной нервной системы.
Любой агент, повышающий натриевую проницаемость мембраны, является раздражителем возбудимой ткани. Раздражителями нервных и мышечных волокон могут быть: электрический ток, механические воздействия (щипок, удар, разрез), резкое охлаждение или согревание, различные кислоты, щелочи, концентрированные растворы солей и т. д.
Среди всех указанных раздражителей электрический ток занимает особое место, так как, во-первых, он может быть легко и точно дозирован по силе, длительности и крутизне нарастания, а во-вторых, он не повреждает живую ткань и его действие быстро и полностью обратимо при тех его силах, которые достаточны для вызова возбуждения.
Изучение действия электрического раздражения на возбудимые ткани представляет большой интерес для физиологии, потому что проведение возбуждения в нервах и мышцах осуществляется с помощью локальных электрических токов, возникающих между возбужденным и покоящимся участком ткани.
В лабораторных условиях и при проведении некоторых клинических исследований для раздражения нервов и мышц применяют электрические стимулы различной формы: прямоугольной, синусоидальной, линейно и экспоненциально нарастающей, индукционные удары, конденсаторные разряды и т. п.
Механизм раздражающего действия тока при всех видах стимулов в принципе одинаков, однако в наиболее отчетливой форме он выявляется при использовании постоянного тока прямоугольной формы.
Для того чтобы раздражитель вызвал возбуждение, он должен иметь достаточную силу, длительность и крутизну нарастания. Та наименьшая сила раздражителя, которая необходима для возникновения потенциала действия в возбудимой ткани, называется порогом раздражения. Стимулы, сила которых ниже пороговой величины, называются подпороговыми, а более сильные, чем пороговые,- сверхпороговыми.
При использовании в качестве раздражителя электрического тока порог выражается в единицах силы тока или напряжения. Абсолютная величина порога зависит от свойств и физиологического состояния ткани, а также от способа нанесения раздражения.
Существует два способа подведения электрического тока к ткани: внеклеточный и внутриклеточный. Первый состоит в том, что оба электрода располагают на поверхности раздражаемой ткани. Ток входит в ткань в области анода и выходит в области катода.
Недостаток этого метода заключается в значительном ветвлении тока: только часть его проходит через мембраны клеток, часть же ответвляется в межклеточные щели. Вследствие этого при раздражении приходится применять значительно большую силу тока, чем это в действительности необходимо для возникновения возбуждения.
Более точным является второй способ раздражения посредством внутриклеточного электрода. Микроэлектрод с диаметром кончика около 0,5 мК вводят в клетку, второй - обычный электрод - прикладывают к поверхности ткани. В этом случае весь приложенный ток проходит через мембрану клетки, что позволяет точно определить величину порога раздражения: у различных клеток он варьирует в пределах 10
9 а. Внутриклеточное раздражение обычно сочетают с регистрацией потенциалов через другой, внутриклеточный электрод.
Минимальное время, в течение которого электрический ток должен действовать на ткань, чтобы вызвать распространяющееся возбуждение, находится в обратной зависимости от напряжения и силы тока.
Если по оси абсцисс отложить минимально необходимое время действия электрического стимула (например, толчка постоянного тока) в миллисекундах, а по оси ординат -- напряжение или силу тока, то мы получим кривую силы -- времени. Эта кривая была подробно изучена в опытах на различных нервах и мышцах Л. Гоорвегом, Г. Вейссом, Л. Лапиком, а в недавнее время Д. Н. Насоновым с сотрудниками.
Из рассмотрения этой кривой прежде всего следует, что ток ниже некоторой минимальной силы или напряжения не вызывает возбуждения, как бы длительно он не действовал. Минимальная сила тока (или напряжение), способная вызвать возбуждение, названа Л. Лапиком реобазой.
Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно.
При очень коротких стимулах кривая силы - времени становится параллельной оси ординат. Это означает, что при таких кратковременных раздражениях возбуждение возникает, как бы ни была велика сила раздражения. Кривая силы - времени имеет форму равносторонней гиперболы.
Определение полезного времени практически трудно, так как величина реобазы претерпевает непрерывно небольшие колебания, отражающие колебания функционального состояния мембраны в покое. По этой причине Л. Лапик (1909) предложил измерять другую, условную, величину, названную им хронаксией. Хронаксия - это наименьшее время, в течение которого электрический ток, равный удвоенной реобазе (ОО), должен действовать на ткань, чтобы вызвать возбуждение. Полезное время и хронаксия характеризуют скорость возникновения возбуждения при действии раздражителя.
Опыты показали, что кривые силы - времени у самых разнообразных тканей, например нервов и мышц человека и теплокровных животных, желудка лягушки, ноги улитки и др., имеют одну и ту же форму.
Различия между ними лишь количественные: в нервных и мышечных волокнах позвоночных животных хронаксия измеряется тысячными и десятитысячными долями секунды, а в так называемых медленных тканях, например в мышечных волокнах ноги улитки или желудка лягушки,- в сотых долях секунды.
Эти факты привели исследователей к выводу, что возбудимые ткани отличаются друг от друга временной константой.
Определение хронаксии -- хронаксиметрия -- получило распространение не только в эксперименте, но и в клинической практике (А. Бургиньон, Ю. М. Уфлянд и др.). В частности, путем измерения хронаксии мышцы невропатолог может установить наличие повреждения волокон двигательного нерва. Дело в том, что при приложении электрического стимула к мышце ток проходит и через находящиеся в ней нервные волокна и их окончания.
Порог раздражения и хронаксия нервных волокон ниже, чем мышечных волокон. Поэтому при раздражении мышцы возбуждение прежде возникает в нервных волокнах и от них уже передается мышечным волокнам. Из этого следует, что при определении хронаксии нормальной мышцы человека фактически измеряется хронаксия иннервирующих ее нервных волокон. Если же нерв поврежден или произошла гибель иннервирующих мышцу клеток в спинном мозгу, то нервные волокна перерождаются, и тогда приложенный к мышце стимул выявляет хронаксию мышечных волокон, которая имеет большую продолжительность.
Величина порога раздражения нерва или мышцы зависит не только от длительности действия стимула, но и от крутизны нарастания его силы. Порог раздражения имеет наименьшую величину при толчках тока прямоугольной формы, характеризующихся максимально быстрым нарастанием силы.
Если же вместо толчков прямоугольной формы применять линейно или экспоненциально нарастающие стимулы, то пороги оказываю увеличенными и тем в большей мере, чем медленнее нарастает сила тока.
При уменьшении крутизны нарастания тока ниже некоторой минимальной величины потенциал действия вообще не возникает, до какой бы конечной силы не увеличивался ток. Обусловлено это тем, что за время нарастания силы раздражителя в ткани успевают развиться активные изменения, повышающие порог и препятствующие возникновению возбуждения.
Такое явление приспособления возбудимой ткани к медленно нарастающему раздражителю получило название аккомодации. Чем выше скорость аккомодации, тем более круто должен нарастать стимул, чтобы не утратить своего раздражающего действия.
Аккомодация развивается не только при раздражении возбудимых тканей электрическим током, но также и при применении механических, термических и прочих раздражителей.
Показателем скорости аккомодации является та наименьшая крутизна нарастания тока, при которой раздражающий стимул еще сохраняет способность вызывать потенциал действия. Эту минимальную крутизну нарастания тока называют минимальным градиентом, или критическим наклоном. Его выражают или в абсолютных величинах -- мА /сек, или в относительных единицах - реобаза/сек. При этом реобазу измеряют прямоугольным током, а затем рассчитывают, на сколько реобаз в секунду должен нарастать ток, чтобы он не утратил раздражающего действия.
Скорость аккомодации различных возбудимых образований широко варьирует. Наиболее велика скорость аккомодации двигательных нервных волокон теплокровных животных. Чувствительные волокна характеризуются меньшей скоростью аккомодации. Очень мала скорость аккомодации волокон сердечной мышцы, гладких мышц кишечника, мочеточников, желудка, т. е. всех образований, которые склонны к автоматической активности.
Постоянный электрический ток обладает полярным действием на возбудимую ткань. Оно выражается в том, что в момент замыкания цепи постоянного тока возбуждение в нерве или мышце всегда возникает только под катодом, а в момент размыкания только под анодом.
Э. Пфлюгер, открывший эти явления, доказал их путемследующего опыта: он умерщвлял участок нерва под одним из электродов, а второй электрод устанавливал на неповрежденный участок. Если с неповрежденным участком соприкасался катод, то возбуждение возникало в момент замыкания тока; если же катод устанавливали на поврежденном участке, а анод на неповрежденном, то возбуждение возникало только при размыкании тока. Порог раздражения при размыкании, когда возбуждение возникает под анодом, значительно выше, чем при замыкании, когда возбуждение возникает под катодом.
О возникновении возбуждения Пфлюгер судил косвенно по сокращению мышцы, иннервируемой раздражаемым нервом. В дальнейшем эти явления, обобщенные в форме полярного закона раздражения, были подтверждены и прямым способом - путем регистрации потенциалов действия непосредственно в участке приложения к ткани полюсов постоянного тока.
Для изучения механизма полярного действия электрического тока в настоящее время производят раздражение нервных и мышечных волокон и отведение от них электрических потенциалов с помощью внутриклеточных микроэлектродов. Установлено, что потенциал действия возникает только в том случае, если катод соприкасается с наружной поверхностью мембраны, а анод находится внутри клетки. При обратном расположении полюсов, т. о. наружном аноде и внутреннем катоде, возбуждение при замыкании тока не возникает, как бы он силен ни был.
Читайте также:
