
Что такое ганглии у насекомых
Насекомые - самые многочисленные животные нашей планеты, распространившиеся практически повсеместно. Порой, эти существа демонстрируют отнюдь не заурядные способности: одни обладают отличной памятью, другие без труда планируют эффективные маршруты, справляясь с логическими задачками не хуже людей. Но как им это удается? Есть ли у насекомых мозг? Об их строении и умственных талантах мы и поговорим.
Особенности насекомых
Насекомые - отдельный класс беспозвоночных членистоногих животных, освоивший все мыслимые и немыслимые экологические ниши. Они обитают в воде, могут передвигаться по воздуху и под землей и встречаются даже в Антарктиде.
Внешний вид и расцветка насекомых очень разнообразны, а их размеры колеблются от 0,2 миллиметров до 30-40 сантиметров. Их симметричное тело состоит из нескольких отделов и сверху покрыто кутикулой из хитина, защищающей его от повреждений. У всех насекомых только три пары ног, что отличает их от других членистоногих. Многие виды обладают крыльями. Но конструкция их сильно отличается от крыльев птиц, представляя собой тонкие пластины, пронизанные жилками, которые играют роль каркаса.
Умственные способности насекомых всегда были спорным вопросом. В древности им нередко приписывали человеческие черты и считали интеллектуалами животного мира. Века спустя все изменилось, и внимание ученых было приковано к млекопитающим. С развитием нейронаук и психологии ученые снова обратили свой взгляд к насекомым, разглядев в них потенциал к усвоению новой информации.

Есть ли у насекомых мозг?
В нашем восприятии насекомые часто воспринимаются как примитивные создания. По строению и поведению он, действительно, уступают человеку и другим млекопитающим. Однако на вопрос, есть ли у насекомых мозг, ответ будет положительным.
Их центральная нервная система представлена цепочкой нервных узлов ганглиев, которые соединены между собой одиночными или парными стволами нервных волокон. В передней части ЦНС находится мозг. Конечно, он совсем не похож на человеческий, и очень упрощен в сравнении с ним. Мозг насекомых состоит из трех ганглиев, сросшихся друг с другом. Каждый из них представляет один отдел:
- Протоцеребрум – отвечает за зрение и сложное поведение.
- Дейтоцеребрум – отвечает за антенны, или усики насекомых.
- Тритоцеребрум – отвечает за мышцы вокруг рта и деятельность внутренних органов.
Первые два отдела особенно важны для взаимодействия насекомых с окружающим миром и своими собратьями. Ученые отмечают, что протоцеребрум у видов с более сложным поведением развит лучше, а количество в нем грибовидных тел, отвечающих за выработку устойчивых ассоциаций, больше. Например, у пчелы около грибовидные тела занимают около 20% объема мозга, а у мухи дрозофилы только 2%.
Отдел дейтоцеребрум не менее полезен. Он отвечает не столько за умственные способности животных, сколько за их ориентацию в пространстве. Усики насекомых, которые контролирует этот отдел, являются органами чувств и выполняют множество функций одновременно. Они могут заменять зрительные, слуховые, осязательные, обонятельные рецепторы и чувствовать температуру воздуха.

Разум насекомых
Теперь, когда мы узнали, есть ли у насекомых мозг, давайте разберемся, на что он способен. Начнем с того, что его размеры невероятно малы. Весить он может всего один миллиграмм, и содержать около 100 миллионов нейронов. У человека же мозг весит 1,5-2 килограмма и содержит 100 миллиардов нейронов. Несмотря на это, насекомые могут выполнять довольно сложные действия и способны обучаться.

Обучение
Интеллект насекомых позволяет им усваивать новую информацию и использовать ее для поисков пищи. Например, пчела отлично различает цвета и запоминает расположение объектов. По ним она и ориентируется, чтобы возвращаться по нескольку раз к цветку, в котором нашла много нектара. Кроме того, она запоминает и время, когда бутон был раскрыт.
Как показали недавние исследования, шмели тоже способны к обучению. В Лондонском университете их сумели научить закатывать мячик в обозначенное место для получения сладкого сиропа. После того как им несколько раз показали принцип действия, шмели легко запомнили и повторяли его.

Навигация
Насекомые отлично ориентируются в пространстве и могут без труда находить места, где бывали ранее. Медоносные пчелы и муравьи запоминают обстановку нужной им локации, а также объекты-маркеры по дороге к ней. В отличие от них, жуков-навозников не останавливает даже ночь. Для поиска нужного пути они полагаются на звезды, а именно на Млечный путь, который отчетливо видно в ясную погоду.
Шмели, относящиеся к семейству настоящих пчел, тоже прекрасно ориентируются. Помимо запоминания местности, они способны прокладывать к местам кормежки наиболее эффективные маршруты. Ученые утверждают, что решая задачку коммивояжера, они используют те же схемы и алгоритмы, которые используют и люди.
 |
 |
Головной мозг насекомых представляет собой сложно организованный надглоточный ганглий. Его верхнюю часть составляет протоцеребрум с большими зрительными долями, иннервирующими глаза. Туда же относятся грибовидные тела, управляющие наиболее сложными движениями и рефлекторной деятельностью, поэтому грибовидные тела лучше всего развиты у общественных насекомых (например, муравьи, пчелы и др.) со сложными внутрисемейными отношениями. Ниже располагается дейтоцеребрум, его значительную часть составляют обонятельные доли, иннер-вирующие усики. Самую нижнюю часть головного мозга и большую часть окологлоточных коннективов, ведущих к подглоточному ганглию, образует тритоцеребрум. Этот отдел мозга развит значительно хуже предыдущих, поскольку у насекомых отсутствует вторая пара антенн (напомним, что у ракообразных тритоцеребрум иннервирует именно их). Все отделы головного мозга парные и сообщаются с одноименным ганглием противоположной стороны посредством комиссуральных проводящих путей.
Брюшная нервная цепочка состоит из крупного под-глоточного ганглия (иннервирует части ротового аппарата), трех менее крупных грудных ганглиев (иннервируют, главным образом, мышцы конечностей и крыловые мышцы) и самых мелких брюшных ганглиев (иннервируют сегменты брюшка). Число брюшных ганглиев у разных видов неодинаково, но всегда меньше количества сегментов в брюшке (только у некоторых личинок может быть 11 ганглиев, но у взрослых особей их всегда меньше). У многих насекомых соседние ганглии сливаются, при этом возникают более крупные скопления нервных клеток.
У насекомых хорошо развита вегетативная (симпатическая) нервная система. Она начинается от нервов, отходящих от головного мозга, и включает в себя собственные ганглии. Вегетативная нервная система управляет деятельностью внутренних органов.
Секреты нейросекреторных клеток наряду с другими железами внутренней секреции обеспечивают гуморальную регуляцию деятельности организма.
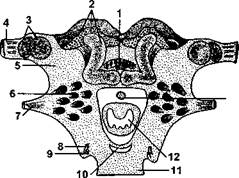 |
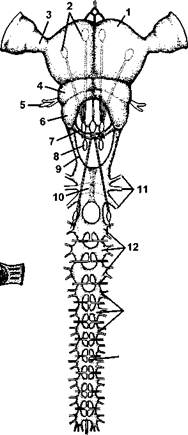
Рис. 980.Схема строения нервной системы насекомого: 1 - протоцеребрум; 2 - нейросекретор-ные клетки; 3 - оптическая область мозга; 4 - дейтоцеребрум; 5 - антеннальный нерв; 6 - тритоцеребрум; 7 - кардиаль-ные тела; 8 - прилежащие тела; 9 — окологлоточные коннективы; 10 - подгло-точный ганглий; 11 - нервы, идущие к ротовым конечностям; 12 — ганглии грудных сегментов; 13 - ганглии брюшных сегментов; 14 - непарный нерв симпатической системы (из Вюрмбаха)
Рис. 981.Схема внутреннего строения надглоточного ганглия черного таракана: 1 - центральное тело; 2 - грибовидное тело; 3 - зрительные доли; 4 - зрительный нерв; 5 — протоцереб-ральные доли; 6 - обонятельные доли; 7 — антеннальный нерв; 8 - фронтальный нерв; 9 - верхнегубной нерв; 10- тритоцеребральная коммиссура; 11 — подглоточ-ный ганглий; 12 - глотка; 13 - ротожелудочный нерв (из Брайтшнайдера)
Рис. 982.Развитие брюшной нервной цепочки
у домашней пчелы Ар1з теННега:
А - личинка; Б - взрослое насекомое (по Бланшару
Основной структурный и рабочий элемент нервной системы — нейрон. Это специализированная нервная клетка с собственной оболочкой, набором внутриклеточных органелл и нейрофибриллами (рис. 111). От её тела отходят длинный осевой отросток — аксон и короткие ветвящиеся дендриты. Получая нервные импульсы от других нейронов, дендриты переводят их на аксон, по которому возбуждение распространяется без затухания на другие нейроны или эффекторы — разного рода железы и мышцы.
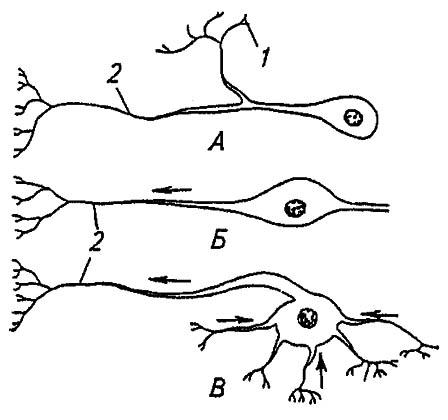
Рисунок 111. Типы нейронов насекомых (по Gillot, 1980):
А, Б, В — соответственно униполярный, биполярный, мультиполярный нейрон; 1 — дендриты; 2 — аксон
В зависимости от исполняемых функций нейроны подразделяются на сенсорные (чувствительные), проводящие возбуждение от рецепторов к нервным центрам; ассоциативные (вставочные, или интернейроны), перерабатывающие поступающую в центры информацию; моторные (двигательные) нейроны, которые доносят в центробежном направлении до эффекторов возбуждение, полученное от ассоциативных нейронов (рис. 112). Сформированная таким образом трёхнейронная рефлекторная дуга обеспечивает целесообразное реагирование на стимулы, тогда как образованная только сенсорным и моторным нейронами двухнейронная дуга обусловливает однозначный ответ на раздражение. Свойственная более примитивным организмам, она не характерна для насекомых, рефлекторные дуги которых нередко включают по нескольку ассоциативных нейронов, обеспечивая тем самым весьма сложные формы реагирования.
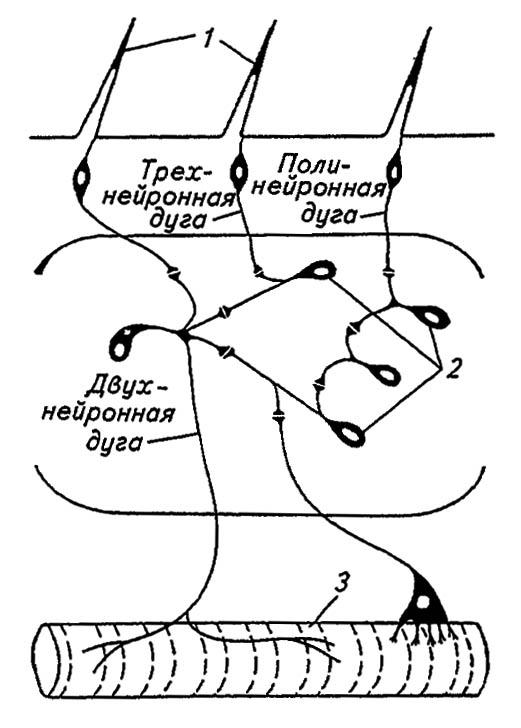
Рисунок 112. Рефлекторные дуги насекомых (по Тыщенко, 1977):
1 — рецепторы; 2 — ассоциативные нейроны; 3 — эффектор
Взаимоотношения разных нейронов в нервных центрах показаны на схеме рефлекторных дуг (см. рис. 112), а их взаимное положение — на поперечном срезе одного из ганглиев брюшной нервной цепочки (рис. 113). Облечённый соединительнотканной оболочкой (нейрилемма) и подстилающим слоем клеток (перинейрум), каждый ганглий прямокрылых насекомых имеет дополнительную защиту от контакта с гемолимфой в виде слоя клеток жирового тела и эпителия трахей. Клетки перинейрума регулируют транспорт ионов через нейрилемму и, поддерживая постоянство ионного состава, обеспечивают проведение нервного возбуждения без внешних помех. Эти клетки также переводят из гемолимфы необходимые для нейронов вещества, передавая их внутриганглионарным (нейроглиальным) клеткам. Последние через цитоплазматическне выросты питают нейроны или образуют вокруг их аксонов плотные спирально закрученные оболочки со свойствами изоляторов.
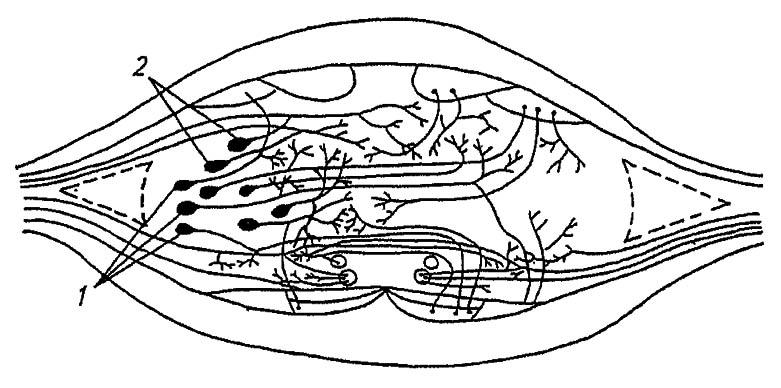
Рисунок 113. Поперечный срез через ганглий брюшной нервной цепочки. Тела ассоциативных (1) и моторных (2) нейронов (по Шванвичу, 1949)
Нейроны с обслуживающими их нейроглиальными клетками распределены во внешнем, корковом, слое ганглия, непосредственно под перинейрумом, тогда как центральная часть ганглия — нейропиль — занята сплетенными нервными волокнами. На периферии нейропиля с дорзальной стороны проходят аксоны моторных нейронов, формируя выходящие из ганглия дорзальные корешки боковых нервов. С вентральной стороны в нейропиле сосредоточены аксоны сенсорных нейронов, входящие в ганглий через вентральные корешки боковых нервов, а центр нейропиля заполнен разветвлениями ассоциативных нервных клеток.
Сквозь нейропиль проходят также пучки нервных волокон, связывающих нейроны обеих сторон одного ганглия и соседних ганглиев друг с другом. Общее число нейронов в пределах одного ганглия обычно не превышает 1 000, а в пределах всей нервной системы насекомого измеряется несколькими миллионами. Это весьма малые величины по сравнению с таковыми у млекопитающих, однако сложность реакций и форм поведения насекомых свидетельствует о совершенстве их нервной системы.
Центральная нервная система
Она образована двойной цепью ганглиев, связанных между собой продольными коннективами и поперечными комиссурами (рис. 114). Её передний отдел — надглоточный ганглий, по положению и функциям отвечающий головному мозгу позвоночных животных, происходит из слившихся между собой ганглиев по меньшей мере трёх сегментов. Вслед за ним расположен под-глоточный ганглий, связанный с головным мозгом окологлоточными коннективами. В его состав входят соединённые друг с другом ганглии трёх сегментов челюстей. Далее следуют три грудных ганглия, изредка образующие единую ганглиозную массу, и, наконец, ганглии брюшных сегментов. Закладывающиеся у эмбриона в виде сегментарных зачатков, они позднее смещаются вперёд и нередко объединяются. У имаго наиболее примитивных насекомых (Thysanura) сохраняется не более восьми парных ганглиев брюшка, причём последний, самый крупный из них, имеет следы объединения 2–3 ганглиев.
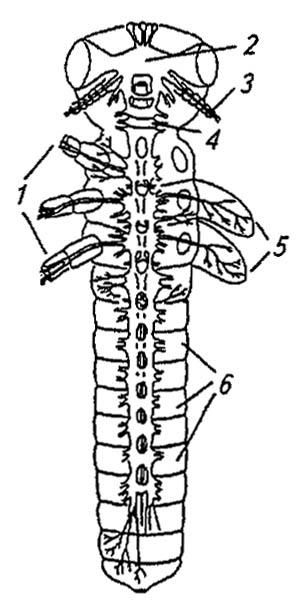
Рисунок 114. Центральная нервная система насекомых (по Шванвичу, 1949):
1 — ноги; 2 — надглоточный ганглий; 3 — антенна; 4 — подглоточный ганглий; 5 — крылья; 6 — ганглии брюшной нервной цепочки
У нимф подёнок и стрекоз в брюшке отмечено семь парных ганглиев, у прямокрылых — пять, другие насекомые имеют часто ещё большую концентрацию брюшной нервной цепочки. Например, центральная нервная система высших мух представлена лишь двумя ганглиозными массами (синганглиями): одна из них — головной мозг, другая — продукт объединения остальных ганглиев. Обычно личинки имеют менее концентрированную нервную систему, чем имаго, но у личинок мух единая ганглиозная масса вообще не разделена (рис. 115).
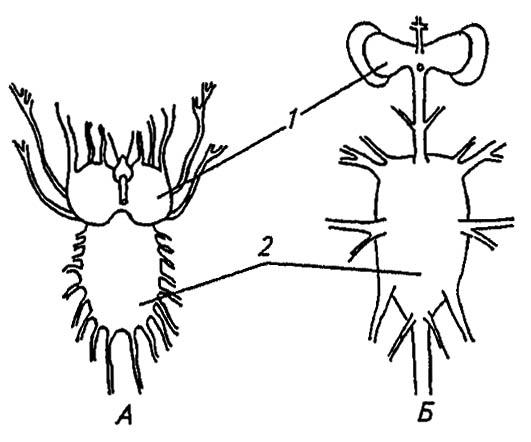
Рисунок 115. Центральная нервная система личинки и имаго мух:
А — личинка; Б — имаго; 1, 2 — соответственно головной и туловищный синганглий
Строение ганглиев брюшной нервной цепочки и ганглиев подглоточного нервного узла сходно; различия между ними определяются различиями иннервируемых органов. В частности, самый последний ганглий контролирует спаривание и откладку яиц и, как и впереди лежащие ганглии, имеет по две пары нервов, включающих сенсорные и моторные волокна. В ганглии крылоносных сегментов входят три пары нервов, обслуживающих ноги, крылья и туловищную мускулатуру, а в ганглий лишённой крыльев переднегруди и в ганглиозные массы подглоточного нервного узла — две пары нервов.
Головной мозг
Особой сложности в строении и во взаимодействии отдельных нервных центров достигает надглоточный ганглий (рис. 116). Образованный протоцеребрумом, дейтоцеребрумом и тритоцеребрумом, он включает ассоциативные центры и ганглиозные массы объединённых сегментов головы.
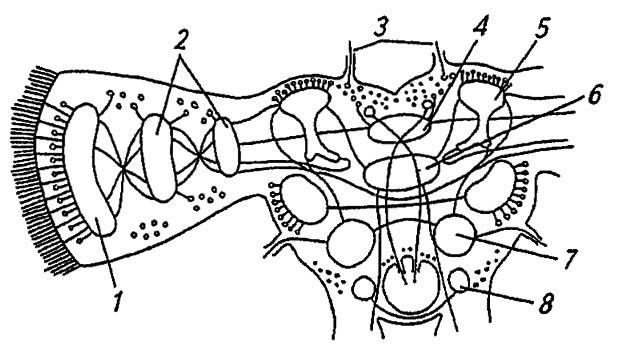
Рисунок 116. Головной мозг насекомых (по Romoser, 1981):
1 — оптическая пластинка; 2 — медуллярные пластинки; 3 — оцеллярные нервы; 4 — протоцеребральный мост; 5, 6 —соответственно грибовидное и центральное тело; 7 — дейтоцеребрум; 8 — тритоцеребрум
Более крупный протоцеребрум обладает явственными оптическими долями (с тремя парами нейропилярных масс и оцеллярным центром), воспринимающими и перерабатывающими сенсорную информацию от сложных глаз и дорзальных глазков насекомого. При этом тела сенсорных нейронов глазков связываются с оцеллярными ганглиями, от которых отходят оцеллярные нервы к межцеребральной части головного мозга. Нейропилярные, то есть образованные отростками нервных клеток, массы протоцеребрального моста и центрального тела соединяют симметричные доли протоцеребрума и, воспринимая аксоны от других центров, координируют моторную активность сегментов тела, определяя, в частности, согласованность работы дыхалец. Примыкающие с обеих сторон к долям протоцеребрума парные стебельчатые (или грибовидные) тела являются высшим ассоциативным центром, где замыкаются условно-рефлекторные связи. Их разрушение ведёт к утрате приобретенных навыков, но безусловные рефлексы сохраняются.
В чашечках стебельчатых тел расположены многочисленные (например, у тараканов до 300 000–400 000) ассоциативные нейроны, аксоны которых проходят в два стебелька, где образуют контакты с аксонами других центров протоцеребрума. Один из этих стебельков (а-доля) связывается с сенсорными, второй (?-доля) — с моторными нейронами (рис. 117). Степень развития стебельчатых тел коррелирует со сложностью поведения насекомых. Например, у муравьёв эти тела занимают 1/5 объёма мозга, а у рабочих пчёл они развиты значительно сильнее, чем у маток или трутней.
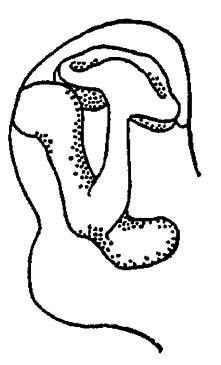
Рисунок 117. Грибовидное (стебельчатое) тело головного мозга насекомых (по Тыщенко, 1977)
Следующий отдел головного мозга — дейтоцеребрум — образован парными антеннальными долями, связанными друг с другом и с сенсорными и моторными аксонами антенн. Отходящий от них ольфакторный тракт достигает чашечек стебельчатых тел, обеспечивая участие ольфакторных стимулов в общей координации нервной деятельности.
Наконец, самый задний отдел головного мозга — крошечный тритоцеребрум — иннервирует головную капсулу с верхней губой и соединяется со стоматогастрической нервной системой (см. ниже) посредством фронтального ганглия и с брюшной нервной цепочкой посредством окологлоточного нервного кольца. Связывающие волокна (поперечная комиссура) между правой и левой долями тритоцеребрума проходят позади рта. Это свидетельствует об исходной принадлежности тритоцеребрума к туловищным сегментам насекомых, лишь вторично вошедшим в состав головы.
Завершая обсуждение строения центральной нервной системы, отметим, что подглоточный ганглий, обслуживающий рецепторы и мышцы челюстей и шейной области, содействует поддержанию двигательной активности насекомого. В этом смысле он служит антагонистом тормозящих влияний головного мозга.
Симпатическая нервная система
Именуемая также висцеральной или вегетативной, она слагается из стоматогастрической (ротожелудочной) системы, системы непарного нерва и каудальной системы (рис. 118). Её высший нервный центр — тритоцеребрум, от которого отходят два фронтальных коннектива к непарному фронтальному ганглию. Образуемый из стенки передней кишки эмбриона, этот ганглий вскоре теряет с ней связь и, соединяясь с головным мозгом, отсылает пару нервов к верхней губе и непарный возвратный нерв к гипоцеребральному ганглию. Последний связывается с нейросекреторными органами ретроцеребрального комплекса и с двумя вентрикулярными ганглиями. Иннервируя передние отделы кишечника и аорту, ганглии стоматогастрической системы включают в себя не только моторные и ассоциативные нейроны, но и сенсорные. В этом отношении они противопоставляются ганглиям брюшной нервной цепочки, с которыми связана система непарного нерва.
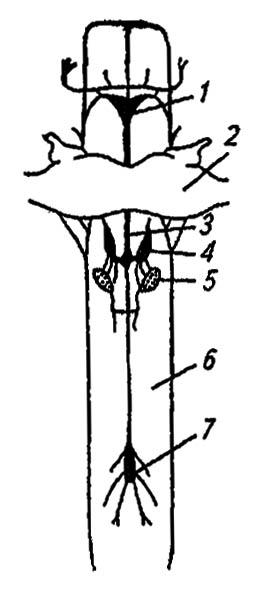
Рисунок 118. Симпатическая нервная система насекомых (по Шванвичу, 1949):
1 — фронтальный ганглий; 2 — мозг; 3 — возвратный нерв; 4 — затылочный ганглий; 5 — прилежащие тела; 6 — пищевод; 7 — вентрикулярный ганглий
Обслуживая дыхальца, эта система связана с ними боковыми ветвями, но образующие их нейроны находятся в грудных и брюшных ганглиях центральной нервной системы. Непарный нерв, по-видимому, участвует в работе крыловых мышц, так как при его разрушении резко возрастает их утомляемость.
Каудальная система, образованная двумя нервами, отходящими от последнего ганглия брюшка, иннервирует заднюю кишку и органы размножения насекомых.
Периферическая нервная система
Все нервы, выходящие из ганглиев центральной и симпатической нервной системы, наряду с сенсорными нейронами органов чувств (рецепторов) образуют периферическую нервную систему насекомых. Некоторые её компоненты будут рассмотрены позднее в связи с сенсорными системами и рецепцией.
Проведение нервного возбуждения в ганглиях
При прохождении возбуждения через ганглий возникают дополнительные ограничения, связанные с необходимостью преодоления по крайней мере нескольких синапсов рефлекторных дуг.
Отличаясь односторонней проницаемостью, синапсы допускают распространение импульсов только в одном направлении — от сенсорных нейронов к моторным, а необходимость выделения медиаторов и их диффузии по синаптической щели всякий раз задерживает импульс на 1–4 мс. Эта задержка многократно возрастает в полинейронных рефлекторных дугах со многими нейронами и синапсами. Вместе с тем нередкое для насекомых объединение множества пресинаптических волокон в одно постсинаптическое проявляется в так называемой пространственной суммации возбуждения; лишь совместное одновременное раздражение нескольких из них генерирует потенциал действия (ПД) в постсинаптическом аксоне. Аналогичным образом слишком слабые, но многократные раздражения, чередуясь в определённом ритме, приводят к временной суммации раздражений и к преодолению синаптического барьера.
Отметим также трансформацию ритма возбуждения во многих синапсах ганглия, своеобразный эффект последействия, связанный с продолжающейся после выключения раздражителя генерацией импульсов, и способность к иррадиации возбуждения на другие рефлекторные дуги ганглия. Наряду с привыканием (то есть блокированием синапса в ответ на монотонные раздражения) и обучением (облегчением переноса возбуждения через синапс) эти свойства ганглиев и синапсов приводят к фильтрации и преобразованию информации, которая поступает в нервные центры от рецепторов. Все это определяет целесообразность реагирования и сложность форм поведения насекомых.
Медиаторы нервного возбуждения в нервно-мышечных контактах и в синапсах ганглиев различны. Есть веские основания считать, что здесь, как и у позвоночных животных, медиатором служит ацетилхолин, разрушаемый ацетилхолинэстеразой. В частности, среди веществ, блокирующих активность ацетилхолинэстеразы насекомых, основное значение имеют фосфорорганические инсектициды.
Не касаясь более сложных аспектов функционирования центральной нервной системы насекомых, укажем, что каждый ганглий брюшной нервной цепочки — первичный рефлекторный центр иннервируемого им сегмента. Однако роль вторичных рефлекторных центров, контролирующих частные функции отдельных ганглиев, принадлежит головному мозгу и подглоточному нервному узлу. Комбинирование сегментарных рефлексов в целостные акты поведения возможно благодаря неспецифическим системам торможения и возбуждения, сравнимым с соответствующими системами млекопитающих.
Головной мозг насекомых исполняет роль высшего анализатора. Принимая участие в анализе сигналов, поступающих от всех органов чувств, он синтезирует акты поведения целостного организма. Лишённые мозга насекомые ведут себя как автоматы с испорченной программой: спонтанные рефлексы теряют приспособительный смысл и противоречат друг другу.
Многие аспекты высшей нервной деятельности насекомых ещё не изучены, но не следует думать, что существование и поведение насекомых целиком подчинены безусловным рефлексам, предопределяющим стереотипные реакции на все стимулы окружающего мира. Некоторые виды (например, пчела) способны к обобщению зрительных образов, превосходя в этом отношении рыб и даже крыс.
Центральная нервная система (ЦНС) насекомых представляет собой цепь отдельных ганглиев, соединённых одиночными или парными коннективами. Самая передняя часть нервной системы, называемая мозгом, занимает дорсальное положение по отношению к пищеварительному каналу. Вся остальная часть ЦНС находится под кишечником. Мозг представляет собой три сросшихся ганглия и разделяется на три отдела: протоцеребрум, отвечающий за зрение и сложные поведенческие реакции, дейтоцеребрум — иннервирует антенны — и тритоцеребрум — иннервирует нижнюю губу, мышцы вокруг ротового отверстия, внутренние органы (высший центр иннервации внутренних органов). От мозга отходят две коннективы, огибающие кишечник с обеих сторон и сходящиеся на подглоточном ганглии, с которого начинается вентральная нервная цепочка. Её ганглии лежат под пищеварительным трактом. Подглоточный ганглий состоит из трёх сросшихся ганглиев и регулирует работу челюстей, гипофаринкса и слюнных желёз. Грудные ганглии развиты сильнее брюшных, так как регулируют работу конечностей и крыльев. Брюшные ганглии у прогрессивных форм имеют тенденцию к слиянию, а вся нервная цепочка — к укорочению. Так, у водомерки Hydrometra в груди имеется один слитный ганглий, а у высших двукрылых (Musca, Sarcophaga и др.) — два ганглия, подглоточный и общий грудной; в брюшке в обоих этих случаях ганглии вообще отсутствуют. Полный метаморфоз часто сопровождается концентрацией ЦНС — уменьшением числа ганглиев у имаго по сравнению с личинками.

У насекомых со сложным поведением (общественные насекомые) особенно сильно развит протоцеребрум, а в нём — увеличенная пара грибовидных тел. В них сосредоточены координирующие и высшие ассоциативные центры нервной системы и замыкаются условно-рефлекторные связи. Приобретённые навыки при их разрушении теряются (но безусловные рефлексы сохраняются). Наблюдается корреляция между сложностью поведения и степенью развития грибовидных тел. Например, среди медоносных пчёл грибовидные тела наиболее развиты у рабочих пчёл (выполняющих все работы), а наименее у трутней. В среднем размер грибовидных тел у медоносных пчёл равен 1⁄15 от размеров всего головного мозга, а у рабочих муравьёв рода формика они составляют почти половину всего мозга.
Вегетативная нервная система иннервирует внутренние органы. Она состоит из трёх отделов — краниального, туловищного и каудального. Краниальный отдел представлен стоматогастрической системой, которая иннервирует переднюю кишку и сердце. Стоматогастрическая система представляет собой соединённый с тритоцеребрумом фронтальный ганглий перед мозгом, соединённый с ним возвратным нервом затылочный ганглий за мозгом, а также кардиальные и прилежащие тела. Кардиальные тела осуществляют секрецию своих гормонов, а также накопление гормонов нейросекреторных клеток мозга. Основная функция прилежащих тел — синтез ювенильных гормонов. Туловищный отдел вегетативной нервной системы представлен непарным нервом, который считается аналогом симпатической нервной системы у позвоночных. В последнем брюшном ганглии непарный нерв образует две ветви, иннервирующие заднюю кишку и половые органы (каудальный отдел вегетативной нервной системы).
1. Краткие сведения об анатомии и гистологии нервной системы
По особенностям организации и функции нервная система насекомых может быть подразделена на центральную, симпатическую и периферическую.
Центральная нервная система имеет метамерное строение: она состоит из серии нервных узлов, или ганглиев, соединенных в нервную цепочку межсегментными стволами - коннективами. Каждый ганглий парный, его левая и правая половины соединены более или менее выраженными комиссурами. Передний отдел нервной цепочки из надглоточного и подглоточного ганглиев называют головным мозгом, а остальные ганглии - брюшной нервной цепочкой. Число ганглиев в брюшной цепочке у разных филогенетических групп насекомых и даже разных стадий их индивидуального развития различно, однако степень интеграции ганглиев, как правило, не коррелирует с уровнем поведения насекомого.
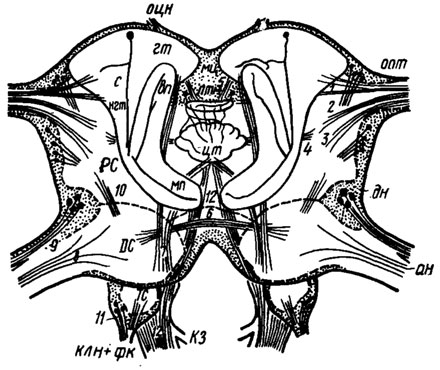
Рис. 1. Схематизированная картина фронтального среза через надглоточный ганглий сверчка Gryllus campestris(по Huber, 1960). Границы нейропиля прото-(PC), дейто-(ДС) и тритоцеребрума (ТС) обведены прерывистой линией. Ассоциативные ядра протоцеребрума обведены сплошными линиями. ан - антеннальный нерв; вп - возвратный придаток ножки грибовидного тела: гт - грибовидное тело; дн - двигательные нейроны дейтоцеребрума; клн+фк - клинео-лабральный нерв и фронтальный коннектив; кЗ - комиссура тритоцере-брума; мп - медиальный придаток ножки грибовидного тела; мц - межцеребральная часть; нгт - ножка грибовидного тела; опт - оптические доли; оцн - оцеллярный нерв; цт - центральное тело; птм - протоцеребральный мост. Нервные пути: 1-3 - оптический тракт; 4 - оптическая комиссура; 5 - оцеллярный тракт; 6 - комиссура дейтоцеребрума; 7- ольфакторно-глобулярный тракт; 8 - афферентные нервы антенн; 9 - эфферентные аксоны, идущие к мышцам антенн; 10 - связь между прото- и дейтоцеребрумом; 11 - моторные аксоны, идущие к верхней губе; 12-нервные пути, связывающие надглоточный ганглий с подглоточным
Надглоточный ганглий - самый крупный и наиболее сложный нервный узел, сходный по функциям с головным мозгом позвоночных. Его образуют, по крайней мере, три слившихся сегментальных ганглия: прото-, дейто- и тритоцеребрум. Более крупный протоцеребрум (рис. 1, PC) тесно связан с фасеточными глазами (или стеммами у личинок Holometabola) и дорсальными глазками, а его грибовидные тела (гт) и примыкающие к ним мозговые ядра служат высшим ассоциативным центром, где замыкаются условные связи. Грибовидные тела наиболее развиты у общественных насекомых, ведущих, в сравнении с остальными, более сложный образ жизни и обладающих высокой способностью к научению. Двустороннее разрушение грибовидных тел влечет выпадение приобретенных навыков, но безусловные реакции на те же стимулы у насекомого сохраняются. Дейтоцеребрум (DC) связан с антеннами, их сенсорным аппаратом и мышцами, поэтому считается обонятельным центром. Миниатюрный тритоцеребрум (ТС) иннервирует верхнюю губу, а также связывает надглоточный ганглий с симпатической нервной системой.
Подглоточный ганглий, также состоящий из трех сегментальных нервных масс, обслуживает сенсиллы и мышцы ротовых частей - соответственно мандибул, максилл и лабиума, а также рецепторы и эффекторы шейной области.
Брюшная нервная цепочка начинается тремя грудными ганглиями, которые у одних насекомых лежат раздельно, у других - сливаются вместе (иногда только мезо- и метаторакальный ганглии) или даже объединяются с несколькими (и изредка со всеми) брюшными ганглиями. Грудные ганглии ответственны в первую очередь за управление движениями ног и крыльев. В них могут также замыкаться условные локомоторные рефлексы.
Менее сложно устроены брюшные ганглии, число которых не превышает 8, иннервируют мышцы брюшных сегментов и их придатков и получают от них сенсорные нервы. Представление о том, в каких комбинациях могут сливаться ганглии брюшной нервной цепочки, дает рис. 2.
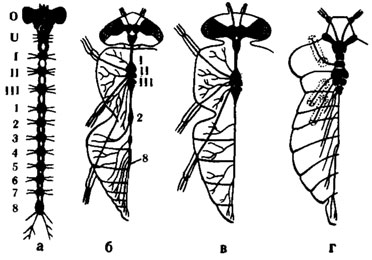
Рис. 2. Различные ступени концентрации центральной нервной системы насекомых(по Eidmann, Kuhlhorn, 1970): а - гипотетическое исходное состояние; б - слепень Tabanus bovinus; в - муха Sarcophaga; г - клоп семейства Pentatomidae. О - надглоточный ганглий; U - подглоточный ганглий; I-III - грудные ганглии; 1-8 - брюшные ганглии
Симпатической, или вегетативной, нервной системы, регулирующей работу внутренних органов, и связанного с ней нейросекреторного аппарата насекомых мы практически не будем касаться в дальнейшем изложении.
Периферическая нервная система состоит из афферентных и эфферентных нервов, чувствующих клеток со вспомогательным аппаратом и эфферентных окончаний. Наше последующее описание будет посвящено в основном именно этой части нервной системы.
Основная структурная единица нервной системы насекомых, как и всех высокоорганизованных животных, - нервная клетка, или нейрон. От тела нейрона, в котором заключено ядро, отходят обычно один длинный неразветвленный аксон и несколько коротких ветвящихся дендритов. В зависимости от числа и взаимного расположения этих отростков говорят о моно-, би- и мультиполярных нейронах. Дендрит служит входом нейрона, а аксон - выходом. Морфологически и функционально различают три главных типа нейронов: чувствующие, двигательные и вставочные. Нас в первую очередь будут интересовать чувствующие и вставочные.
Чувствующие нейроны. Тела чувствующих, или сенсорных, клеток обычно биполярной или мультиполярной формы * лежат всегда вблизи чувствующего органа или иннервируемой ткани. Дендриты одних нейронов, чаще всего биполярных, связаны с кутикулярными образованиями, других, всегда мультиполярных, - с тканями полости тела или же они образуют субэпидермальную сеть, как у личинок с мягкой кожей.
* ( Некоторые авторы относят чувствующие клетки к особой категории нервных клеток и не называют их нейронами.)
Соответственно различают две большие категории чувствующих клеток. Клетки первого типа отличаются тем, что практически всегда связаны с кутикулой или ее впячиваниями: аподемами, трахеями, выстилкой предротовой и ротовой полостей и т. п. К ним принадлежат разнообразные экстерорецепторные клетки, в том числе и зрительные, хотя их дендриты выражены неясно. Клетки второго типа никогда не связаны с кутикулой и лежат только на внутренней поверхности тела, стенках пищеварительного тракта, в мышечной и соединительной тканях. Электрофизиологически показано, что они принадлежат интеро или проприоцепторам.
Аксоны чувствующих клеток идут непосредственно в соответствующие ганглии ЦНС, иногда находящиеся непосредственно в головном мозгу, например оптические или обонятельные центры. Вопрос о каналах связи рецепторных клеток с нервным центром чрезвычайно важен для правильной интерпретации работы анализатора и механизма управления поведением насекомого. Теперь, по-видимому, все признают несостоятельным прежнее мнение о том, что в некоторых рецепторных системах, например в антеннах клопа Rhodnius, имеет место срастание аксонов нескольких чувствующих клеток в единое волокно. Но замыкание группы рецепторов на один периферический нейрон второго порядка, т. е. потеря "адреса" входного сигнала, характерно для первого оптического ганглия насекомых. Смысл такого способа связи с центром, ведущего к частичной потере информации от совокупности датчиков, пока не всегда ясен (см. ниже).
Нервная ткань, в том числе и сенсорные клетки, происходят из эктодермы. Их принадлежность покрову тела выражается и в том, что связь чувствующего органа с ЦНС устанавливается центростремительно. Так, В. Вигглесворс показал на клопе Rhodnius, что перерезанный афферентный нерв регенерирует в направлении к ЦНС. Точно так же во время каждой линьки, когда образуются дополнительные рецепторы, чтобы обслуживать увеличивающуюся поверхность тела, их чувствующие клетки посылают аксоны центростремительно.
Выявленный на гистологических препаратах факт центростремительного развития аксона может стать одним из оснований для важного заключения о том, что путь от чувствующей клетки до ЦНС прямой, без синаптического переключения. Вблизи рецепторных клеток и афферентных нервов встречаются другие, например, нейроглиальные (питающие) клетки, но они не имеют отношения к передаче рецепторного сигнала.
Двигательные (моторные) нейроны. Тела этих обычно униполярных нейронов лежат всегда в сегментальном ганглии, чаще с его вентральной стороны и с боков. Их дендриты густо ветвятся в нейропиле ганглия, вступая в синаптические контакты с вставочными нейронами или даже непосредственно с разветвлениями аксонов чувствующих клеток. Аксоны двигательных нейронов уходят за пределы ганглия к мышцам, где заканчиваются множеством моторных концевых пластинок, или бляшек. Одну и ту же мышцу иннервируют так называемые быстрые и медленные двигательные волокна: по первым идут импульсы, вызывающие быстрые сокращения мышцы, по вторым - вызывающие тоническое сокращение. При этом тело иннервирующего двигательного нейрона может лежать в ганглии как того же сегмента, которому принадлежит мышца, так и соседнего. Часто встречается контралатеральное положение тела двигательного нейрона по отношению к эффектору.
Вставочные нейроны, или интернейроны, выполняют функцию передачи возбуждения с одного нейрона на другой. Они связывают чувствующие клетки с моторными, причем в последовательную цепь может быть включено несколько клеток, и тогда говорят об интернейронах 2-го, 3-го и более высокого порядка. Их обычно униполярные небольшие (от 2-3 мкм и меньше) тела лежат всегда по периферии ганглия, а отростки, в числе которых аксон, нередко т-образный, вступают внутрь ганглия, в его нейропиль. Интернейроны могут быть сегментальными или мультисегментальными, с восходящими и нисходящими волокнами. Последние участвуют, например, в управлении локомоторными реакциями. Благодаря множеству связей интернейроны образуют коммутационные сети для выработки нервных команд, направляемых по специальным трактам в составе комиссур и коннектив.
Среди восходящих волокон у насекомых обращают на себя внимание аксоны так называемых гигантских интернейронов, в частности, берущие начало в последнем брюшном ганглии и идущие по цепочке до грудных и даже до головных центров. Они отличаются большой длиной и толщиной (до 30 мкм и более), а главное, высокой скоростью проведения сигналов. Так, если в обычных афферентных двигательных нервах импульсы распространяются со скоростью 1,5-3,0 м/с, то в гигантском волокне, например таракана Periplanefa, скорость проведения может достигать 5-7 м/с. Считалось, что с помощью таких "быстрых" интернейронов осуществляется реакция убегания, когда опасность грозит насекомому сзади, но теперь этот взгляд признан несостоятельным.
Очень сложна область ганглиозного нейропиля, в котором генерируются нервные команды, управляющие поведением насекомого. Ее образует переплетение отростков дендритов моторных нейронов с разветвлениями ассоциативных и афферентных волокон, связанных друг с другом множеством синаптических контактов. Волокна здесь особенно тонкие: от 0,1 мкм по сравнению 1-3 мкм на выходе из дорсальных (двигательных) и вентральных (чувствующих) корешков. Они содержат протонейрофибриллы диаметром 100-200 А, митохондрии, гранулярный эндоплазматический ретикулум, нейросекреторные включения и прочие элементы. Синаптическая передача основана на поперечных и продольных контактах волокон или на контакте типа терминальной пуговки. Ультраструктура синаптических образований напоминает таковую синапсов у позвоночных: пресинаптическое волокно в районе контакта содержит синаптические пузырьки, наполненные химическим медиатором. Между трехслойными пре- и постсинаптическими мембранами находится синаптическая щель шириной 100-300 А. Как полагают, у насекомых имеется не только холинэнергический, но и адренэнергический механизм синаптической передачи. Не исключается также возможность электрической передачи возбуждения с помощью соответствующих синапсов, в которых импульсы передаются в результате электрического воздействия пресинаптического тока на мембрану постсинаптического волокна. Такие электрические сикапсы недавно обнаружены между зрительными интeрнейронами - детекторами движения, где они обеспечивают максимально высокую скорость проведения сигналов.
Читайте также:
