
Что такое проведение нервного импульса

Проведение нервного импульса, передача сигнала в виде волны возбужденияв пределах одного нейрона и от одной клетки к второй. П. н. и. по нервным проводникам происходит посредством электротонических потенциалов и потенциалов действия, каковые распространяются на протяжении волокна в обоих направлениях, не переходя на соседние волокна (см. Биоэлектрические потенциалы, Импульс нервный).
Передача межклеточных сигналов осуществляется через синапсы значительно чаще посредством медиаторов, вызывающих появление потенциалов постсинаптических. Нервные проводники возможно разглядывать как кабели, владеющие довольно низким осевым сопротивлением (сопротивление аксоплазмы — ri) и более высоким сопротивлением оболочки (сопротивление мембраны — rm).
Нервный импульс распространяется на протяжении нервного проводника при помощи прохождения тока между покоящимися и активными участками нерва (локальные токи). В проводнике по мере повышения расстояния от места происхождения возбуждения происходит постепенное, а при однородной структуры проводника экспоненциальное затухание импульса, что в 2,7 раза значительно уменьшается на расстоянии l = (константа длины).
Так как rm и ri находятся в обратном отношении к диаметру проводника, то затухание нервного импульса в узких волокнах происходит раньше, чем в толстых. Несовершенство кабельных особенностей нервных проводников восполняется тем, что они владеют возбудимостью. Главное условие возбуждения — наличие у нервов потенциала спокойствия.
В случае если локальный ток через покоящийся участок приведёт к деполяризации мембраны, достигающую критического уровня (порога), это приведёт к происхождению распространяющегося потенциала действия (ПД). Соотношение уровня пороговой деполяризации и амплитуды ПД, в большинстве случаев составляющее не меньше 1: 5, снабжает высокую надёжность проведения: участки проводника, владеющие свойством генерировать ПД, смогут отстоять друг от друга на таком расстоянии, преодолевая которое нервный импульс снижает собственную амплитуду практически в 5 раз. Данный ослабленный сигнал будет опять усилен до стандартного уровня (амплитуда ПД) и сможет продолжить собственный путь по нерву.
Скорость П. н. и. зависит от быстроты, с которой мембранная ёмкость на участке впереди импульса разряжается до отметки порога генерации ПД, что, со своей стороны, определяется геометрическими изюминками нервов, трансформациями их диаметра, наличием узлов ветвления. В частности, узкие волокна владеют более высоким ri, и большей поверхностной ёмкостью, а потому скорость П. н. и. по ним ниже.
Одновременно с этим толщина нервных волокон ограничивает возможности существования солидного числа параллельных каналов связи. Конфликт между физическими особенностями нервных проводников и требованиями компактности нервной совокупности был разрешен возникновением на протяжении эволюции позвоночных т. н. мякотных (миелинизированных) волокон (см. Нервы). Скорость П. н. и. в миелинизированных волокнах теплокровных (не обращая внимания на их небольшой диаметр — 4—20 мкм) достигает 100—120 м/сек.
Генерация ПД происходит лишь в ограниченных участках их поверхности — перехватах Ранвье, а по межперехватным участкам П. и. и. осуществляется электротонически (см. Сальтаторное проведение). Кое-какие лекарственные вещества, к примеру анестетики, очень сильно замедляют впредь до полного блока П. н. и. Этим пользуются в практической медицине для обезболивания.
Лит. см. при статьях Возбуждение, Синапсы.
- Радиолярии
- Решётчатые конструкции
- Реаниматология
Вегетативная нервная совокупность, часть нервной совокупности, регулирующая деятельность органов кровообращения, дыхания, пищеварения, выделения,…
Симпатическая нервная совокупность (от греч. sympathes — чувствительный, чувствительный к влиянию), часть вегетативной нервной совокупности позвоночных…
ПРОВЕДЕНИЕ НЕРВНОГО ИМПУЛЬСА, передача сигнала в виде волны возбуждения в пределах одного нейрона и от одной клетки к другой. П. н. и. по нервным проводникам происходит с помощью электротонич. потенциалов и потенциалов действия, к-рые распространяются вдоль волокна в обоих направлениях, не переходя на соседние волокна (см. Биоэлектрические потенциалы, Импульс нервный). Передача межклеточных сигналов осуществляется через синапсы чаще всего с помощью медиаторов, вызывающих появление потенциалов постсинаптических. Нервные проводники можно рассматривать как кабели, обладающие относительно низким осевым сопротивлением (сопротивление аксоплазмы - ri) и более высоким сопротивлением оболочки (сопротивление мембраны - rm). Нервный импульс распространяется вдоль нервного проводника посредством прохождения тока между покоящимися и активными участками нерва (локальные токи). В проводнике по мере увеличения расстояния от места возникновения возбуждения происходит постепенное, а в случае однородной структуры проводника экспоненциальное затухание импульса, к-рый в 2,7 раза уменьшается на расстоянии Л = корень квадратный из rm/ri (константа длины). Так как rm и ri находятся в обратном отношении к диаметру проводника, то затухание нервного импульса в тонких волокнах происходит раньше, чем в толстых. Несовершенство кабельных свойств нервных проводников восполняется тем, что они обладают возбудимостью. Осн. условие возбуждения - наличие у нервов потенциала покоя. Если локальный ток через покоящийся участок вызовет деполяризацию мембраны, достигающую критич. уровня (порога), это приведёт к возникновению распространяющегося потенциала действия (ПД). Соотношение уровня пороговой деполяризации и амплитуды ПД, обычно составляющее не менее 1:5, обеспечивает высокую надёжность проведения: участки проводника, обладающие способностью генерировать ПД, могут отстоять друг от друга на таком расстоянии, преодолевая к-рое нервный импульс снижает свою амплитуду почти в 5 раз. Этот ослабленный сигнал будет снова усилен до стандартного уровня (амплитуда ПД) и сможет продолжить свой путь по нерву.
Рис. 5. Сооружение тоннелей щитовым комплексом: 1 - исполнительный орган; 2 - погрузочный орган; 3 - щит; 4 - блокоукладчик; 5 - передвижная платформа; 6 - перегружатель; 7 - блоковоз; 8 - вагонетки; 9 - блочная крепь.
Скорость П. н. и. зависит от быстроты, с к-рой мембранная ёмкость на участке впереди импульса разряжается до уровня порога генерации ПД, что, в свою очередь, определяется геометрич. особенностями нервов, изменениями их диаметра, наличием узлов ветвления. В частности, тонкие волокна обладают более высоким ri и большей поверхностной ёмкостью, а потому скорость П. н. и. по ним ниже. В то же время толщина нервных волокон ограничивает возможности существования большого числа параллельных каналов связи. Конфликт между физич. свойствами нервных проводников и требованиями "компактности" нервной системы был разрешён появлением в ходе эволюции позвоночных т. н. мякотных (миелинизированных) волокон (см. Нервы). Скорость П. н. и. в миелинизированных волокнах теплокровных (несмотря на их малый диаметр - 4-20 мкм) достигает 100-120 м/сек. Генерация ПД происходит только в ограниченных участках их поверхности - перехватах Ранвье, а по межперехватным участкам П. н. и. осуществляется электротонически (см. Сальтаторное проведение). Нек-рые лекарственные вещества, напр. анестетики, сильно замедляют вплоть до полного блока П. н. и. Этим пользуются в практической медицине для обезболивания.
Лит. см. при статьях Возбуждение, Синапсы. Л. Г. Магазаник.
нервного импульса, передача сигнала в виде волны возбуждения в пределах одного нейрона и от одной клетки к другой. П. н. и. по нервным проводникам происходит с помощью электротонических потенциалов и потенциалов действия, которые распространяются вдоль волокна в обоих направлениях, не переходя на соседние волокна (см. Биоэлектрические потенциалы , Импульс нервный ). Передача межклеточных сигналов осуществляется через синапсы чаще всего с помощью медиаторов, вызывающих появление потенциалов постсинаптических . Нервные проводники можно рассматривать как кабели, обладающие относительно низким осевым сопротивлением (сопротивление аксоплазмы - ri ) и более высоким сопротивлением оболочки (сопротивление мембраны - rm ). Нервный импульс распространяется вдоль нервного проводника посредством прохождения тока между покоящимися и активными участками нерва (локальные токи). В проводнике по мере увеличения расстояния от места возникновения возбуждения происходит постепенное, а в случае однородной структуры проводника экспоненциальное затухание импульса, который в 2,7 раза уменьшается на расстоянии l (константа длины). Так как rm и ri находятся в обратном отношении к диаметру проводника, то затухание нервного импульса в тонких волокнах происходит раньше, чем в толстых. Несовершенство кабельных свойств нервных проводников восполняется тем, что они обладают возбудимостью . Основное условие возбуждения - наличие у нервов потенциала покоя . Если локальный ток через покоящийся участок вызовет деполяризацию мембраны, достигающую критического уровня (порога), это приведёт к возникновению распространяющегося потенциала действия (ПД). Соотношение уровня пороговой деполяризации и амплитуды ПД, обычно составляющее не менее 1: 5, обеспечивает высокую надёжность проведения: участки проводника, обладающие способностью генерировать ПД, могут отстоять друг от друга на таком расстоянии, преодолевая которое нервный импульс снижает свою амплитуду почти в 5 раз. Этот ослабленный сигнал будет снова усилен до стандартного уровня (амплитуда ПД) и сможет продолжить свой путь по нерву.
Скорость П. н. и. зависит от быстроты, с которой мембранная ёмкость на участке впереди импульса разряжается до уровня порога генерации ПД, что, в свою очередь, определяется геометрическими особенностями нервов, изменениями их диаметра, наличием узлов ветвления. В частности, тонкие волокна обладают более высоким ri , и большей поверхностной ёмкостью, а потому скорость П. н. и. по ним ниже. В то же время толщина нервных волокон ограничивает возможности существования большого числа параллельных каналов связи. Конфликт между физическими свойствами нервных проводников и требованиями 'компактности' нервной системы был разрешен появлением в ходе эволюции позвоночных т. н. мякотных (миелинизированных) волокон (см. Нервы ) . Скорость П. н. и. в миелинизированных волокнах теплокровных (несмотря на их малый диаметр - 4-20 мкм ) достигает 100-120 м/сек. Генерация ПД происходит только в ограниченных участках их поверхности - перехватах Ранвье, а по межперехватным участкам П. и. и. осуществляется электротонически (см. Сальтаторное проведение ). Некоторые лекарственные вещества, например анестетики, сильно замедляют вплоть до полного блока П. н. и. Этим пользуются в практической медицине для обезболивания.
Лит. см. при статьях Возбуждение , Синапсы .
Нервный импульс (лат. nervus нерв; лат. impulsus удар, толчок) — волна возбуждения, распространяющаяся по нервному волокну; единица распространяющегося возбуждения.
Нервный импульс обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам — скелетной мускулатуре, гладким мышцам внутренних органов и сосудов, железам внутренней и внешней секреции и т. д.
Распространение Нервных импульсов отождествляется с проведением потенциалов действия (см. Биоэлектрические потенциалы). Возникновение возбуждения может быть результатом раздражения (см.), напр, воздействие света на зрительный рецептор, звука на слуховой рецептор, или процессов, протекающих в тканях (спонтанное возникновение Н. и.). В этих случаях Н. и. обеспечивают согласованную работу органов при протекании какого-либо физиологического процесса (напр., в процессе дыхания Н. и. вызывают сокращение скелетных мышц и диафрагмы, результатом чего являются вдох и выдох, и т. д.).
В живых организмах передача информации может осуществляться и гуморальным путем, посредством выброса в русло крови гормонов, медиаторов и т. п. Однако преимущество информации, передаваемой при помощи Н. и., состоит в том, что она более целенаправленна, передается быстро и может быть точнее закодирована, чем сигналы, посылаемые гуморальной системой.
Факт, что нервные стволы являются путем, по к-рому передаются влияния от мозга к мышцам и в обратном направлении, был известен еще в эпоху античности. В средние века и вплоть до середины 17 в. считалось, что по нервам распространяется некая субстанция, подобная жидкости или пламени. Идея о электрической природе Н. и. возникла в 18 в. Первые исследования электрических явлений в живых тканях, связанных с возникновением и распространением возбуждения, были осуществлены Л. Гальвани. Г. Гельмгольц показал, что скорость распространения Н. и., к-рую ранее считали близкой к скорости света, имеет конечное значение и может быть точно измерена. Германн (L. Hermann) ввел в физиологию понятие потенциала действия. Объяснение механизма возникновения и проведения возбуждения стало возможным после создания С. Аррениусом теории электролитической диссоциации. В соответствии с этой теорией Бернштейн (J. Bernstein) предположил, что возникновение и проведение Н. и. обусловлено перемещением ионов между нервным волокном и окружающей средой. Англ. исследователи А. Ходжкин, Б. Катц и Э. Хаксли детально исследовали трансмембранные ионные токи, лежащие в основе развития потенциала действия. Позже стали интенсивно изучаться механизмы работы ионных каналов, по к-рым происходит обмен ионами между аксоном и окружающей средой, и механизмы, обеспечивающие способность нервных волокон проводить ряды Н. и. разного ритма и продолжительности.
Н. и. распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками нервного волокна. Ток, выходящий из волокна наружу в покоящемся участке, служит раздражителем. Наступающая после возбуждения в данном участке нервного волокна рефрактерность обусловливает поступательное движение Н. и.
Количественно соотношения разных фаз развития потенциала действия можно охарактеризовать, сопоставляя их по амплитуде и длительности во времени. Так, напр., для миелиновых нервных волокон группы А млекопитающих диаметр волокна находится в пределах 1—22 мк, скорость проведения — 5—120 м/сек, длительность и амплитуда высоковольтной части (пика, или спайка) — 0,4—0,5 мсек и 100—120 мв соответственно, следовой негативный потенциал — 12—20 мсек (3—5% от амплитуды спайка), следовой позитивный потенциал — 40—60 мсек (0,2% от амплитуды спайка).
Возможности передачи разнообразной информации расширяются за счет повышения скорости развития потенциала действия, скорости распространения, а также за счет повышения лабильности (см.) — т. е. способности возбудимого образования воспроизводить в единицу времени высокие ритмы возбуждения.
Возникновение Н. и. в нервных клетках (см.) или рецепторах (см.) связано с деполяризацией мембраны, т. е. со снижением величины электрического потенциала на мембране (потенциала покоя, или мембранного потенциала). Если величина мембранного потенциала снижается на 10—20% (пороговый критический уровень), то местный процесс переходит в распространяющийся — возникает потенциал действия (см. Возбуждение).
Конкретные особенности распространения Н. и. связаны со строением нервных волокон (см.). Сердцевина волокна (аксоплазма) обладает низким сопротивлением и, соответственно, хорошей проводимостью, а окружающая аксоплазму плазматическая мембрана — большим сопротивлением. Особенно велико электрическое сопротивление наружного слоя у миелинизированных волокон, у к-рых свободны от толстой миелиновой оболочки только перехваты Ранвье. В безмиелиновых волокнах Н. и. движется непрерывно, а в миелиновых — скачкообразно (сальтаторное проведение).
Различают декрементное и бездекрементное распространение волны возбуждения. Декрементное проведение, т. е. проведение возбуждения с угасанием, наблюдается в безмиелиновых волокнах. В таких волокнах скорость проведения Н. и. невелика и по мере отдаления от места раздражения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания. Декрементное проведение свойственно волокнам, иннервирующим внутренние органы, обладающие низкой функц, подвижностью. Без декрементное проведение характерно для миелиновых и тех безмиелиновых волокон, к-рые передают сигналы к органам, обладающим высокой реактивностью (напр., сердечной мышце). При бездекрементном проведении Н. и. проходит весь путь от места раздражения до места реализации информации без затухания.
Передача Н. и. с нервного волокна на мышечное или какой-либо другой эффектор осуществляется через синапсы (см.). У позвоночных животных в подавляющем большинстве случаев передача возбуждения на эффектор происходит при помощи выделения ацетилхолина (нервно-мышечные синапсы скелетной мускулатуры, синаптические соединения в сердце и др.). Для таких синапсов характерно строго одностороннее проведение импульса и наличие временной задержки передачи возбуждения.
В синапсах, в синаптической щели которых сопротивление электрическому току благодаря большой площади контактирующих поверхностей мало, происходит электрическая передача возбуждения. В них нет синаптической задержки проведения и возможно двустороннее проведение. Такие синапсы свойственны беспозвоночным животным.
Регистрация Н. и. нашла широкое применение в биол, исследованиях и клин, практике. Для регистрации используют шлейфные и чаще катодные осциллографы (см. Осциллография). При помощи микроэлектродной техники (см. Микроэлектродный метод исследования) регистрируют Н. и. в одиночных возбудимых образованиях — нейронах и аксонах. Возможности исследования механизма возникновения и распространения Н. и. значительно расширились после разработки метода фиксации потенциала. Этим методом были получены основные данные о ионных токах (см. Биоэлектрические потенциалы).
Нарушение проведения Н. и. происходит при повреждении нервных стволов, напр, при механических травмах, сдавливании в результате разрастания опухоли или при воспалительных процессах. Такие нарушения проведения Н. и. зачастую бывают необратимы. Следствием прекращения иннервации могут быть тяжелые функциональные и трофические расстройства (напр., атрофия скелетных мышц конечностей после прекращения поступления Н. и. вследствие необратимой травмы нервного ствола). Обратимое прекращение проведения Н. и. может быть вызвано специально, в терапевтических целях. Напр., с помощью анестезирующих средств блокируют импульсацию, идущую от болевых рецепторов в ц. н. с. Обратимое прекращение проведения Н. и. вызывает и новокаиновая блокада. Временное прекращение передачи Н. и. по нервным проводникам наблюдается и во время общего наркоза.
Библиография: Бpеже М. А. Электрическая активность нервной системы, пер. с англ., М., 1979; Жуков Е. К. Очерки по нервно-мышечной физиологии, Л., 1969; Коннели К. Восстановительные процессы и обмен веществ в нерве, в кн.: Совр, пробл. биофизики, пер. с англ., под ред. Г. М. Франка и А. Г. Пасынского, т. 2, с. 211, М., 1961; Костюк П. Г. Физиология центральной нервной системы, Киев, 1977; Латманизова Л. В. Очерк физиологии возбуждения, М., 1972; Общая физиология нервной системы, под ред. П. Г. Костюка, Л., 1979; Тасаки И. Нервное возбуждение, пер. с англ., М., 1971; Ходжкин А. Нервный импульс, пер. с англ., М., 1965; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975.
Структура нервного волокна. Проведение нервных импульсов является специализированной функцией нервных волокон, т.е. отростков нервных клеток.
Нервные волокна разделяют намякотные, или миелинизированные, и безмякотные, или немиелинизированные. Мякотные, чувствительные и двигательные волокна входят в состав нервов, снабжающих органы чувств и скелетную мускулатуру; они имеются также в вегетативной нервной системе. Безмякотные волокна у позвоночных животных принадлежат в основном симпатической нервной системе.
Нервы обычно состоят как из мякотных, так и из безмякотных волокон, причем их соотношение в разных нервах различное. Например, во многих кожных нервах преобладают безмякотные нервные волокна. Так, в нервах вегетативной нервной системы, например в блуждающем нерве, количество безмякотных волокон достигает 80—95%. Наоборот, в нервах, иннервирующих скелетные мышцы, имеется лишь относительно небольшое количество безмякотных волокон.
Как показали электронно-микроскопические исследования, мие- линовая оболочка создается в результате того, что миелоцит (шван- новская клетка) многократно обертывает осевой цилиндр (рис. 2.27'), слои ее сливаются, образуя плотный жировой футляр — миелиновую оболочку. Миелиновая оболочка через промежутки равной длины прерывается, оставляя открытыми участки мембраны шириной примерно 1 мкм. Эти участки получили название перехватов Ранвье.

Рис. 2.27. Роль миелоцита (шванновской клетки) в образовании миелиновой оболочки в мякотных нервных волокнах: последовательные стадии спиралеобразного закручивания миелоцита вокруг аксона (I); взаимное расположение миелоцитов и аксонов в безмякотных нервных волокнах (II)
Длина межперехватных участков, покрытых миелиновой оболочкой, примерно пропорциональна диаметру волокна. Так, в нервных волокнах диаметром 10—20 мкм длина промежутка между перехватами составляет 1—2 мм. В наиболее тонких волокнах (диаметром
1—2 мкм) эти участки имеют длину около 0,2 мм. [1]
Безмякотные нервные волокна не имеют миелиновой оболочки, они изолированы друг от друг только шванновскими клетками. В простейшем случае одиночный миелоцит окружает одно безмякот- ное волокно. Часто, однако, в складках миелоцита оказывается несколько тонких безмякотных волокон.
Миелиновая оболочка выполняет двоякую функцию: функцию электрического изолятора и трофическую функцию. Изолирующие свойства миелиновой оболочки связаны с тем, что миелин как вещество липидной природы препятствует прохождению ионов и потому обладает очень высоким сопротивлением. Благодаря существованию миелиновой оболочки возникновение возбуждения в мякот- ных нервных волокнах возможно не на всем протяжении осевого цилиндра, а только в ограниченных участках — перехватах Ранвье. Это имеет важное значение для распространения нервного импульса вдоль волокна.
Трофическая функция миелиновой оболочки, по-видимому, состоит в том, что она принимает участие в процессах регуляции обмена веществ и роста осевого цилиндра.
Прямые электрофизиологические исследования, проведенные Като (1924), а затем Тасаки (1953) на одиночных миелинизированных нервных волокнах лягушки, показали, что потенциалы действия в этих волокнах возникают только в перехватах, а участки между перехватами, покрытые миелином, являются практически невозбудимыми.
В состоянии покоя наружная поверхность возбудимой мембраны всех перехватов Ранвье заряжена положительно. Разности потенциалов между соседними перехватами не существует. В момент возбуждения поверхность мембраны перехвата С становится заряженной электроотрицательно по отношению к поверхности мембраны соседнего перехвата D. Это приводит к возникновению местного (ло 
Рис. 2.28. Сальтаторное распространение возбуждения в мякотном нервном волокне от перехвата к перехвату:
А — немиелинизированное волокно; В — миелинизированное волокно. Стрелками показано направление тока
кального) электрического тока, который идет через окружающую волокно межтканевую жидкость, мембрану и аксоплазму в направлении, показанном на рисунке стрелкой. Выходящий через перехват D ток возбуждает его, вызывая перезарядку мембраны. В перехвате С возбуждение еще продолжается, и он на время становится рефрактерным. Поэтому перехват D способен привести в состояние возбуждения только следующий перехват и т.д.
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
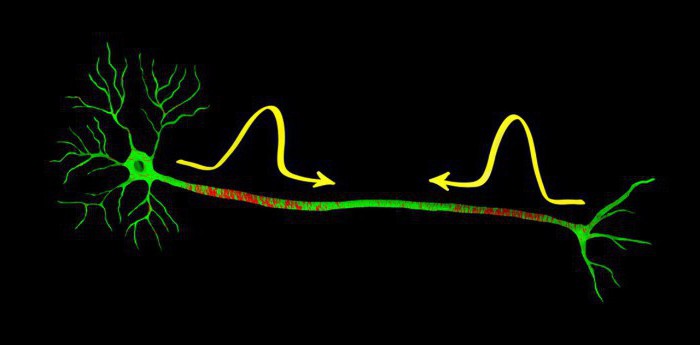
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.
Читайте также:
