
Чувствительный ганглий слухового анализатора
Спиральный ганглий располагается в канале Розенталя, который находится в модиолусе улитки. Он содержит тела афферентных нейронов, дендриты, которых возбуждаются под действием нейромедиаторов, секретируемых клетками кортиева органа, и аксоны, которые направляются в улитковые ядра расположенные в стволе головного мозга. Большинство (приблизительно 95%) афферентных волокон являются толстыми, покрыты миелиновой оболочкой и берут начало от нейронов I типа.
Эти волокна иннервируют исключительно внутренние волосковые клетки. Оставшиеся афферентные волокна тонкие, не имеют миелиновой оболочки и исходят из нейронов II типа, эти волокна контактируют с наружными волосковыми клетками. Около десятка нейронов I типа иннервируют каждую внутреннюю волосковую клетку (сходящаяся иннервационная модель), в то время как афферентные нервные волокна II типа разделяются на несколько ветвей и контактируют с несколькими наружными волосковыми клетками (расходящаяся иннервационная модель).
Вся слуховая информация переносится к стволу мозга по афферентной системе. Слуховой и вестибулярный нервы соединяются друг с другом в преддверно-улитковом нерве (VIII черепной нерв).
Эфферентные волокна берут начало в стволе головного мозга от нейронов, расположенных в комплексе верхней оливы и посылают информацию к улитке посредством синаптических соединений с наружными волосковыми клетками, а также с афферентными волокнами внутренних волосковых клеток. Эфферентная система позволяет центральной нервной системе регулировать работу улитки.
Иннервационная структура Кортиева органа отражает функциональное различие двух типов улитковых волосковых клеток.
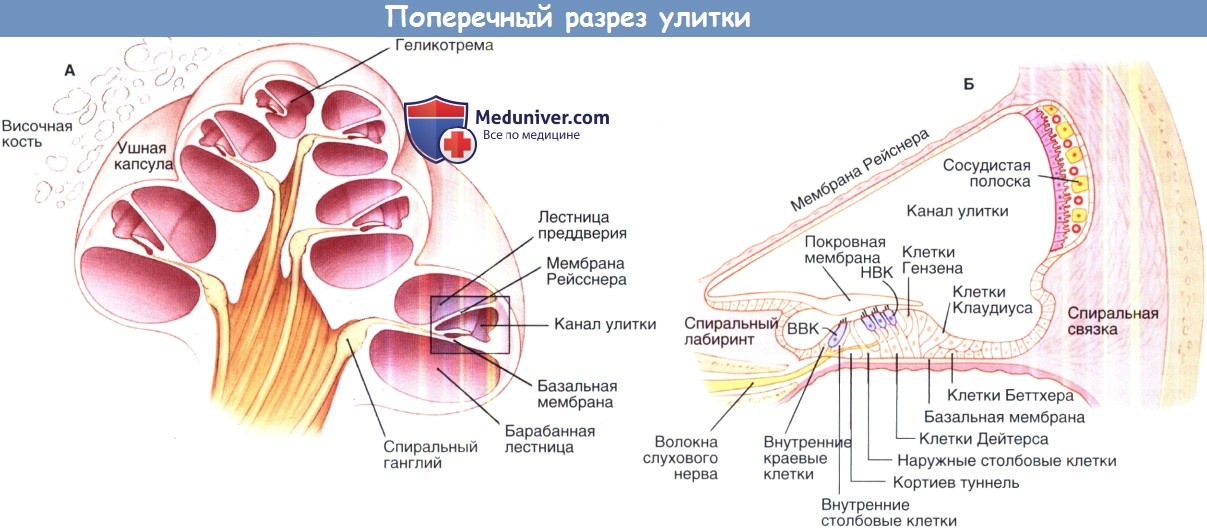
Поперечный разрез улитки. А, схематичное изображение улитки в разрезе.
Выделенная область, показанная на рисунке Б, изображает улитковый проток и окружающие структуры, такие как кортиев орган и сосудистую полоску в разрезе.
ВВК — внутренние волосковые клетки; НВК — наружные волосковые клетки.
а) Нейропроцессинг слуховой информации и синапсы внутренних волосковых клеток. Афферентная нейропередача, осуществляемая внутренними волосковыми клетками, инициируется синапсами ленточного типа количеством от 5 до 30, где поток ионов кальция через потенциал-зависимые кальциевые каналы приводит к четко дозированному высвобождению синаптических пузырьков в пресинаптическую область. В результате экзоцитоз нейротрансмиттеров прямо пропорционален пресинаптическому току кальция, который в свою очередь зависит от изменения потенциала, обусловленного механотрансдукцией.
Информация, кодируемая на афферентных синапсах, поразительно точна, что позволяет получить высокую временную точность, так же как и значительный динамический диапазон с пятикратной амплитудой от 0 до более 100 дБ.
Каждый ленточный синапс состоит из пресинаптического плотного тела (схожего с ленточным синапсом фоторецепторных клеток) в окружении пузырьков с нейротрансмиттерами, плотной полоски пресинаптической мембраны, синаптической щели и постсинаптической области, содержащей АМРА-глютаминэргические рецепторы афферентных нейронов. Считается что глутамат или вещество, близкое по составу, является нейротрансмиттером афферентных связей внутренних волосковых клеток, хотя в этом процессе могут также быть задействованы другие, еще нераспознанные трансмиттеры.
Тонотопическая организация кортиева органа происходит благодаря афферентной системе, где деполяризация внутренних волосковых клеток в определенных участках ведет к возбуждению афферентных нейронов спирального ганглия. Каждое афферентное волокно характеризуется определенной настроечной кривой, описывающей необходимый для получения ответа на заданной частоте уровень звукового давления. Особенность настроечных кривых заключается в отражении частоты наивысшей чувствительности нервного волокна, т.е. частотной характеристики.
Стимулы выше или ниже характерной частоты могут также вызывать ответ, но только при наличии более высокой интенсивности. Форма и пороги чувствительности настроечных кривых афферентных нервных волокон зависят от множества факторов, включая морфологию кортиева органа и активные процессы, связанные с улитковой амплификацией. Активность эфферентной системы играет важную роль в модуляции характеристик афферентных нервов. Потеря улитковой амплификации, в частности, в результате утраты наружных волосковых клеток, ведет к расширению настроечных кривых и увеличению порогов ответа нервного волокна.
Тонотопическая организация улитки является основой частотной кодировки в волокнах слухового нерва (кодирование позиции). Кроме того, выходные характеристики частот кодируются волокнами слухового нерва, это явление известно, как фазовая синхронизация. Волокна слухового нерва посылают сигнал в определенной фазе с частотным стимулом, приводящим к регулярному ответу с характерными интервалами между пиками, равными или кратными длине волны стимула. Фазовая синхронизация возникает только на низких частотах.
Свыше 5 кГц возбуждение, идущее от волокон слухового нерва, возникает с хаотическими интервалами. Тонотопическая организация и фазовая синхронизация крайне важны для выделения сигнала нужной частоты. Выходная частота волокон слухового нерва определяется не только частотными характеристиками, но также и интенсивностью стимулов. По мере увеличения интенсивности стимула увеличивается импульсная частота слухового нерва. Аналогичным образом количество волокон слуховых нервов, активированных на заданных частотах, увеличивается по мере усиления стимула.
Это является результатом вовлечения в процесс большего количества нервных волокон в результате того, что волокна слухового нерва при одинаковых частотных характеристиках имеют разные пороги частотного ответа. Кроме того, при росте интенсивности стимула активируются и другие афферентные нервные волокна с похожими частотными характеристиками. Эти важные физиологические характеристики определяют ряд правил, обязательных при изготовлении и применении имплантатов улитки.
Таким образом, частотно-зависимая стимуляция волосковых клеток ведет к увеличению синаптической передачи в ленточных синапсах внутренних волосковых клеток. Последующее возбуждение постсинаптических токов в волокнах афферентного нерва ведет к временной разрядке потенциала действия в слуховом нерве. В действительности, временное разрешение и динамический диапазон слуховой системы не имеют равных среди сенсорных систем.
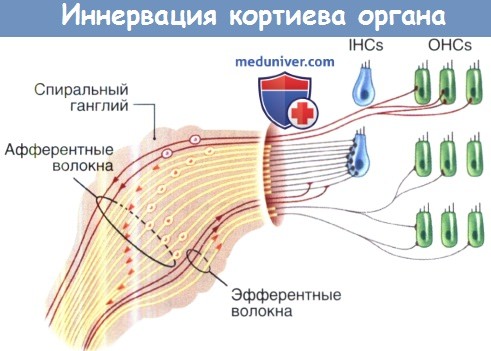
Иннервация кортиева органа.
Схема отображает афферентную и эфферентную иннервацию внутренних и наружных волосковых клеток.
Сверху вниз показаны немиелиновые и миелиновые афферентные волокна II типа,
немиелоновые эфферентные волокна латеральных оливокохлеарных путей (ЛОК) и миелиновые эфферентные волокна медиальных оливокохлеарных путей (МОК).
б) Эфферентная иннервация улитки. В головном мозге четко различают две группы эфферентных нервных волокон. Первые, миелиновые медиальные оливокохлеарные (МОК) эфферентные волокна, берут начало от нейронов, расположенных около верхнего медиального оливного ядра. МОК волокна направляются к контраи ипсилатеральной улитке, где они образуют холинергические синапсы с наружными волосковыми клетками. Вторая группа волокон представлена безмиелиновыми латеральными оливокохлеарными (ЛОК) эфферентными волокнами, которые берут начало от нейронов с маленькими телами, локализующимися около латерального верхнего ядра оливы.
ЛОК волокна направляются преимущественно к ипсилатеральной улитке, где они оканчиваются на дендритах афферентных нейронов I типа у основания внутренних волосковых клеток. Синапсы ЛОК волокон являются сложным химическим комплексом и используют холинэргическую, ГАМК-эргическую и дофаминэргическую передачу, а также различные нейропептиды.
Эффект от стимуляции МОК волокон был изучен подробнее, чем стимуляция ЛОК волокон. В целом стимуляция МОК системы ведет к увеличению порогов чувствительности из-за снижения степени улитковой амплификации наружными волосковыми клетками. Это обусловленная звуком обратная связь, следовательно, чувствительность слухового аппарата снижается в ситуациях, когда нет нужды в энергозатратных механизмах метаболической амплификации.
Функция ЛОК нервных волокон представляется более сложной. Их непосредственный вход в афферентные нейроны позволяет предположить, что они регулируют афферентную активность, тем самым оказывая влияние на динамический диапазон. Исследование различных патологических состояний подтверждает эту точку зрения тем, что снижение специфических нейротрансмиттеров или разрушение тел нейронов в стволе головного мозга ведет либо к усилению, либо к супрессии ответа слухового нерва.
Эффекты обратной связи ЛОК волокон медленны и обычно проявляются в течение минут. Еще одна функция ЛОК системы заключается в обеспечении медленной интеграции и настройки бинауральной функции, необходимой для точного различения локализации звука.
В итоге, активация МОК и ЛОК эфферентной системы, по-видимому, имеет защитный эффект, направленный против акустического повреждения и такая обратная связь может быть важна в шумной окружающей среде.
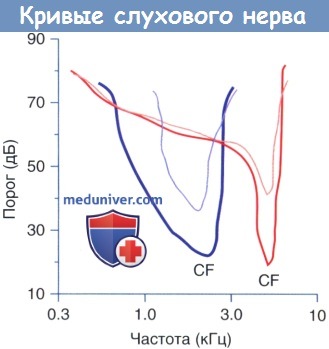
Характерные настроечные кривые слухового нерва (толстая линия).
Настроечная кривая слухового нерва с частотной характеристикой около 2 кГц (синяя) и 5 кГц (красная) (тонкие линии).
Значительное снижение порогов слуховой чувствительности в ответ на стимуляцию МОК системы.
Изменения специфической формы настроечных кривых зависят от частотной характеристики (CF) отдельно взятого волокна.
- Вернуться в оглавление раздела "отоларингология"
Слуховые проводящие пути и низшие слуховые центры - это проводниковая афферентная (приносящая) часть слуховой сенсорной системы, проводящая, распределяющая и преобразующая сенсорное возбуждение, порождённое слуховыми рецепторами, для формирования рефлекторных реакций эффекторов и слуховых образов в высших слуховых центрах коры.
Все слуховые центры, начиная от кохлеарных ядер и вплоть до коры головного мозга, устроены тонотопически, т.е. рецепторы кортиева органа проецируются в них на строго определенные нейроны. И, соответственно, эти нейроны обрабатывают информацию о звуках только определённой частоты, определённой высоты звучания. Чем дальше по слухового пути от улитки находится слуховой центр, тем более сложными звуковыми сигналами возбуждаются его отдельные нейроны. это говорит о том, что в слуховых центрах происходит всё более усложняющийся синтез отдельных характеристик звуковых сигналов.
Нельзя считать, что информация о звуковых сигналах обрабатывается только последовательно при переходе возбуждения от одного слухового центра к другому. Все слуховые центры связаны между собой многочисленными сложными связями, с помощью которых осуществляется не только перенос информации в одном направлении, но и её сравнительная обработка.
1 - улитка (Кортиев орган с волосковыми клетками - слуховыми рецепторами);
2 - спиральный ганглий;
3 - переднее (вентральное) улитковое (кохлеарное) ядро;
4 - заднее (дорзальное) улитковое (кохлеарное) ядро;
5 - ядро трапециевидного тела;
6 - верхняя олива;
7 - ядро латеральной петли;
8 - ядра задних холмиков четверохолмия среднего мозга;
9 - медиальные коленчатые тела метаталамуса промежуточного мозга;
10 - проекционная слуховая зона коры больших полушарий головного мозга.

Рис. 1. Схема слуховых сенсорных путей (по Сентаготаи).
1 — височная доля; 2 — средний мозг; 3 — перешеек ромбовидного мозга; 4 — продолговатый мозг; 5 — улитка; 6 — вентральное слуховое ядро; 7 — дорсальное слуховое ядро; 8 — слуховые полоски; 9 — оливо-слуховые волокна; 10 — верхняя олива: 11 — ядра трапециевидного тела; 12 — трапециевидное тело; 13 — пирамида; 14 — латеральная петля; 15 — ядро латеральной петли; 16 — треугольник латеральной петли; 17 — нижнее двухолмие; 18 — латеральное коленчатое тело; 19 — корковый центр слуха.
Схематический путь слухового возбуждения : слуховые рецепторы (волосковые клетки в Кортиевом органе улитки уха) - периферический спиральный ганглий (в улитке) - продолговатый мозг (сначала кохлеарные ядра, т.е. улитковые, после них - ядра оливы) - средний мозг (нижнее двухолмие) - промежуточный мозг (медиальные коленчатые тела, они же внутренние) - кора больших полушарий головного мозга (слуховые зоны височных долей, поля 41, 42).
Первые (I) слуховые афферентные нейроны (биполярные нейроны) находятся в спиральном ганглии, или узле (gangl. spirale), расположенном в основании полого улиткового веретена. Спиральный ганглий состоит из тел слуховых биполярных нейронов. Дендриты этих нейронов проходят по каналам костной спиральной пластинки к улитке уха, т.е. они начинаются от наружных волосковых клеток Кортиева органа. Аксоны выходят из спирального узла и собираются в слуховой нерв, вступающий в области мостомозжечкового угла в ствол мозга, где и заканчиваются синапсами на нервных клетках улитковых (кохлеарных) ядер: дорсального (nucl. cochlearis dorsalis) и вентрального (nucl. cochlearis ventralis). Эти клетки кохлеарных ядер являются вторыми слуховыми нейронами (II).
Слуховой нерв имеет следующие названия: N. vestibulocochlearis, sive n. octavus (PNA), n. acusticus (BNA), sive n. stato-acusticus - равновесно-слуховой (JNA). Это VIII пара черепно-мозговых нервов, состоящая из двух частей: улитковой (pars cochlearis) и вестибулярной, или преддверной (pars vestibularis). Улитковая часть является совокупностью аксонов I нейронов слуховой сенсорной системы (биполярных нейронов спирального ганглия), преддверная - аксоны афферентных нейронов лабиринта, обеспечивающие регулирование положения тела в пространстве (в анатомической литературе обе части также называются нервными корешками).
Вторые слуховые афферентные нейроны (II) находятся в дорсальном и вентральном кохлеарном (улитковом) ядре продолговатого мозга.
От нейронов II кохлеарных ядер начинаются два восходящих слуховых тракта. Контрлатеральный восходящий слуховой путь содержит в себе основную массу выходящих из комплекса кохлеарных ядер волокон и образует три пучка волокон: 1- вентральная слуховая полоска, или трапециевидное тело, 2 - промежуточная слуховая полоска, или полоска Хельда, 3 - задняя, или дорсальная, слуховая полоска - полоска Монакова. Основную часть волокон содержит в себе первый пучок - трапециевидное тело. Средняя, интермедиальная, полоска образована аксонами части клеток заднего отдела заднего вентрального ядра кохлеарного комплекса. Дорсальная слуховая полоска содержит в себе волокна, идущие от клеток дорсального кохлеарного ядра, а также аксоны части клеток заднего вентрального ядра. Волокна дорсальной полоски идут по дну четвертого желудочка, затем уходят в ствол мозга, пересекают среднюю линию и, минуя оливу, не оканчиваясь в ней, присоединяются к латеральной петле противоположной стороны, где поднимаются к ядрам латеральной петли. Эта полоска обходит верхнюю ножку мозжечка, затем переходит на противоположную сторону и присоединяется к трапециевидному телу.
Итак, аксоны II нейронов, отходящие от клеток дорсального ядра (слухового бугорка), образуют мозговые полоски (striae medullares ventriculi quarti), находящиеся в ромбовидной ямке на границе моста и продолговатого мозга. Большая часть мозговой полоски переходит на противоположную сторону и около средней линии погружается в вещество мозга, подключаясь к латеральной петле (lemniscus lateralis); меньшая часть мозговой полоски присоединяется к латеральной петле своей же стороны. Многочисленные волокна, выходящие из дорсального ядра, идут в составе боковой петли и оканчиваются в нижних бугорках четверохолмия среднего мозга (colliculus inferior) и во внутреннем (медиальном) коленчатом теле (corpus geniculatum mediate) таламуса, это промежуточный мозг. Часть волокон, минуя внутреннее коленчатое тело (слуховой центр), идет в наружное (латеральное) коленчатое тело таламуса, являющееся зрительным подкорковым центром промежуточного мозга, что указывает на тесную связь между слуховой сенсорной системой и зрительной.
Аксоны II нейронов от клеток вентрального ядра участвуют в образовании трапециевидного тела (corpus trapezoideum). Большая часть аксонов в составе боковой петли (lemniscus lateralis) переходит на противоположную сторону и оканчивается в верхней оливе продолговатого мозга и ядрах трапециевидного тела, а также в ретикулярных ядрах покрышки на слуховых нейронах III. Другая, меньшая, часть волокон оканчивается на своей же стороне в тех же структурах. Поэтому именно здесь, в оливах, проходит сравнение акустических сигналов, поступающих с двух сторон от двух разных ушей. Оливы обеспечивают бинауральный анализ звуков, т.е. сопоставляют звуки от разных ушей. Именно оливы обеспечивают стереозвучание и помогают точно нацелиться на источник звука.
Третьи слуховые афферентные нейроны (III) находятся в ядрах верхней оливы (1) и трапециевидного тела (2), а также в нижнем двухолмии среднего мозга (3) и в внутренних (медиальных) коленчатых телах (4) промежуточного мозга. Аксоны III нейронов участвуют в образовании латеральной петли, в которой имеются волокна II и III нейронов. Часть волокон II нейронов прерывается в ядре латеральной петли (nucl. lemnisci proprius lateralis). Таким образом, в ядре латеральной петли тоже есть III нейроны Волокна II нейронов латеральной петли переключаются на III нейроны в медиальном коленчатом теле (corpus geniculatum mediale). Волокна III нейронов латеральной петли, пройдя мимо медиального коленчатого тела, заканчиваются в нижнем двухолмии (colliculus inferior), где формируется tr. tectospinalis. Таким образом, в нижнем двухолмии среднего мозга находится низший слуховой центр, состоящий из IV нейронов.
Нервные волокна латеральной петли, которые относятся к нейронам верхней оливы, из моста проникают в верхние ножки мозжечка и затем достигают его ядер. Таким образом, ядра мозжечка получают слуховое сенсорное возбуждение из слуховых низших нервных центров оливы. Другая часть аксонов верхней оливы направляется к мотонейронам спинного мозга и далее к поперечнополосатым мышцам. Таким образом, слуховые низшие нервные центры верхней оливы управляют эффекторами и обеспечивают двигательные слуховые рефлекторные реакции.
Аксоны III нейронов, расположенных в медиальном коленчатом теле (corpus geniculatum mediate), пройдя через заднюю часть задней ножки внутренней капсулы, формируют слуховое сияние, которое заканчивается на IV нейронах в высшем нервном слуховом центре коры - поперечной извилине Гешля височной доли (поля 41, 42, 20, 21, 22). Итак, аксоны III нейронов медиальных коленчатых тел образуют центральный слуховой путь, ведущий в слуховые сенсорные первичные проекционные зоны коры больших полушарий головного мозга. Кроме восходящих афферентных волокон, в центральном слуховом пути проходят также и нисходящие эфферентные волокна - от коры к низшим подкорковым слуховым центрам.
Четвёртые слуховые афферентные нейроны (IV) находятся как в нижнем двухолмии среднего мозга, так и в височной доле коры больших полушарий головного мозга (поля 41, 42, 20, 21, 22 по Бродману).
Нижнее двухолмие является рефлекторным двигательным центром, через который подключается tr. tectospinalis. Благодаря этому при слуховом раздражении рефлекторно подключается спинной мозг для выполнения автоматических движений, чему способствует и подключение верхней оливы с мозжечком; подключается также медиальный продольный пучок (fasc. longitudinalis medialis), объединяющий функции двигательных ядер черепных нервов. Разрушение нижнего двухолмия не сопровождается потерей слуха, однако оно играет важную роль "рефлекторного" подкоркового центра, в котором формируется эфферентная часть ориентировочных слуховых рефлексов в виде движения глаз и головы.
Тела корковых нейронов IV образуют колонки слуховой коры, формирующих первичные слуховые образы. От некоторых IV нейронов идут пути через мозолистое тело на противоположную сторону, в слуховую кору контралатерального (противоположного) полушария. Это последний путь слухового сенсорного возбуждения. Он заканчивается тоже на IV нейронах. Слуховые сенсорные образы формируются в высшем нервном слуховом центре коры - поперечной извилине Гешля височной доли (поля 41, 42, 20, 21, 22). Низкие звуки воспринимаются в передних отделах верхней височной извилины, а высокие звуки — в её задних отделах. Поля 41 и 42, а также 41/42 височной области коры относятся к мелкоклеточным (пылевидным, кониокортикальным) чувствующим полям коры мозга. Они располагаются на верхней поверхности височной доли, скрытой в глубине латеральной (сильвиевой) борозды. В поле 41, наиболее мелко- и густоклеточном, заканчивается большая часть афферентных волокон слуховой сенсорной системы. Другие поля височной области (22, 21, 20 и 37) выполняют высшие слуховые функции, например, участвуют в слуховом гнозисе. Слуховой гнозис (gnosis acustica) - это узнавание предмета по характерному для него звуку.
При заболевании периферических отделов слуховой сенсорной системы в слуховом восприятии возникают шумы, звуки различного характера.
Для понижения слуха центрального происхождения характерно нарушение высшего акустического (звукового) анализа звуковых раздражений. Иногда отмечается патологическое обострение или извращение слуха (гиперакузия, паракузия).
При корковом поражении наступают сенсорная афазия и слуховая агнозия. Расстройство слуха наблюдается при многих органических заболеваниях центральной нервной системы.
2. В этих органах рецепцию раздражений осуществляют не нейросенсорные, а
- специализированные производные эпителия (п. 16.1.2).
2. Но, как и в первично чувствующих органах, соответствующее раздражение вызывает
17.1. Орган слуха и равновесия
17.1.1. Общие сведения
17.1.1.1. Составные части
В составе органа - 3 части: наружное, среднее и внутреннее ухо.
I. Наружное ухо
2. а) Ушная раковина и первая треть слу х ового прохода имеют хрящевую основу;
II. Среднее ухо
III,а. Внутреннее ухо: костный лабиринт
2. Средняя часть - преддверие (vestibulum) (8) - овальная полость, сообщающаяся с соседними отделами лабиринта .
III,б. Внутреннее ухо: перепончатый лабиринт
2. а) Во внутреннем ухе имеется жидкость:
3. а) Итак, в костной улитке находится перепончатая улитка (15) , занимающая примерно треть сечения спирального костного канала.
б ) В костном преддверии перепончатый лабиринт образует два мешочка -
17.1.1.2. Распределение рецепторных функций между частями лабиринта
б) Причём, в каждой части лабиринта (улитке, мешочках преддвери я , полукружных каналах) рецепторные образования имеют строго определённую функцию .
II. Четыре рецепторных образования
в) При этом колебания перилимфы улитки
В отсутствие круглого окна стремечко, из-за несжимаемости жидкости, не могло бы приводить перилимфу в движение.
г) Одновременно колебания перилимфы (и эндолимфы) улитки вызывают
б) При этом н ад чувствительными клетками пятна имеется студенистая мембрана ,
которая, в зависимости от гравитационного притяжения,
б) П ри вращении головы в определённой плоскости
III. Резюме
Вышеизложенное распределение функций можно суммировать в таблице. –
| Спиральный (кортиев) орган (в перепончатой улитке) | Восприятие звука |
| Пятно эллиптического мешочка | Восприятие гравитации |
| Пятно сферического мешочка | Восприятие гравитации и вибрации |
| Ампулы полукружных каналов | Восприятие угловых ускорений при вращении головы и тела |
17.1.1.3. Развитие органа слуха и равновесия
2. При их впячивании последовательно появляются слуховые ямки и слуховые пузырьки , выстланные
(Следовательно, все эпителиоциты рецепторных образований органа слуха и равновесия развиваются из данного типа эмбрионального эпителия ).
3. Затем слуховые пузырьки разделяются перетяжкой на два отдела -
4 . а) Одновременно у станавливается связь со слуховыми нервными ганглиями,
которые делятся на две части - преддверную и улитковую .
Теперь обратимся к гистологическому строению органа слуха и равновесия.
17.1.2. Наружное и среднее ухо
1. В основе ушной раковины (1) -
Его строение и препарат см. в п. 10.2.5.
1. В коже наружного слухового прохода (2) - не только сальные, но и церуминозные железы, которые выделяют ушную серу .
1. Барабанная перепонка (3) покрыта:
В ней преобладают коллагеновые волокна, но имеются и эластические.
Барабанная полость (4) , к ак и внутренняя поверхность барабанной перепонки, выстлана
1. Стенка слуховой трубы (6) выстлана слизистой оболочкой.
Последняя похожа на таковую в дыхательных путях:
2. Под слизистой оболочкой -
17.1.3. Улитковый отдел лабиринта
17.1.3.1. Внутреннее строение костной улитки
I. Срединная часть костной улитки
2. От стержня отходит костная пластинка - спиральный гребешок (2) . Последний

б) Край лимба делится на две части -
II. Подразделение канала костной улитки на 3 части
2. В итоге, канал костной улитки на всём своём протяжении подразделяется на 3 части:
б) Обе лестницы содержат перилимфу, а перепончатая улитка - эндолимфу.
3. а) При этом вестибулярная лестница

б) Барабанная же лестница заканчивается у круглого окна преддверия.
4. Поэтому колебания перилимфы проходят по следующей цепочке:
17.1.3.2. Стенки перепончатой улитки
а) П ерепончат ый лабиринт улитки имеет на разрезе почти треугольную форму .
б) Охарактеризуем три его стороны.
I. Верхнемедиальная стенка
б) Она п рикрепляется к лимбу в области его верхней (вестибулярной) губы (11).
2. Вестибулярная мембрана включает три слоя:

II. Наружная стенка
2. Она имеет два слоя :
3. а) Сосудистая полоска представляет собой

в) Функция сосудистой полоски - трофическая и секреторная ( продукция эндолимфы).
4. Спиральная связка
III. Нижняя стенка
б) Она натянута между
б) А. Основу пластинки составляют коллагеновые волокна, расположенные в матриксе.
Б. Возможно, волокна играют роль струн , которые откликаются на колебания определённой частоты.
3. Со стороны эндолимфы на базилярной пластинке располагается спиральный, или кортиев, орган (9), который
IV. Внутренний угол канала перепончатой улитки
2. В перепончатом канале улитки лимб образует внутренний угол - между
17.1.3.3. Спиральный (кортиев) орган
Наконец, обратимся к ключевому элементу органа слуха - кортиеву органу.
I. Клеточный состав
в (II) (Среднее увеличение)


II. Поддерживающие клетки
б) На каждой такой клетке, как на ложе, располага е тся сенсорн ая клетк а .
III. Сенсорные волосковые клетки
Сенсорные клетки находятся на фаланговых клетках (1); поэтому, в соответствии с числом последних,
2. Апикальная поверхность
На апикальной поверхности сенсорных клеток присутствуют два образования:
а) кутикула (4) - плёнка гликопротеидной природы,
б) особые микроворсинки - стереоцилии (5), которые
17.1.3.4. Восприятие звуковых раздражений
в) Видимо, эфферентные волокна оказывают
г) Медиаторами в образумых ими синапсах являются:
17.1.3.5. Спиральный ганглий

17.1.4. Вестибулярный отдел лабиринта
17.1.4.1. Эллиптический и сферический мешочки преддверия
I. Рецепторные пятна
2. Строение пятен того и другого мешочка практически одинаково. –
В эпителии пятна - три элемента:
II. Сенсорные клетки рецепторных пятен
1. Сенсорные клетки
2. Контакт с нервными окончаниями
а) Грушевидные клетки:
(хотя собственно синаптические контакты образуются только в некоторых участках этой чаши).
б) Цилиндрические клетки:
а) В апикальных отделах тех и других клеток име ю тся
III. Восприятие гравитационных воздействий
1. Цепочка событий такова:
2. В результате, мозг
17.1.4.2. Рецепторные участки полукружных каналов

б) (Среднее увеличение)

в) (Большое увеличение)

б) При вращении тела
17.2. Орган вкуса
17.2.1. Вкусовые почки
а) (Малое увеличение)

б) (Среднее увеличение)

2. а) В частности, на снимк ах - листовидные сосочки (1), покрытые многослойным плоским неороговевающим эпителием.
б) Между сосочками - узкие просветы (2).
в) С этими просветами контактируют вкусовые почки (3) округлой или овальной формы.
3. В эмбриогенезе эти почки развиваются
17.2.2. Клеточный состав
17.2.2.1. Три вида клеток вкусовой почки
( Большое увеличение) 
17.2.2.2. Характеристика клеток
I. Сенсорные, или "светлые", клетки
б) При этом, как правило, во вкусовой почке представлены рецепторы лишь какого-то одного вкуса.
б) Возбуждение вкусовых рецепторов на микроворсинках
II. Прочие клетки вкусовой почки
Б. Благодаря им, происходит постоянное обновление сенсорных и поддерживающих клеток вкусовых почек
Читайте также:
