
Дегенерация и регенерация нервного волокна
При повреждении нервов дегенеративные процессы аксона происходят дистальнее места поражения. Это связано с тем, что аксон является отростком нейрона, и его питание и рост зависят от сомы. Регенеративные процессы периферической нервной системы характеризуются высокой интенсивностью и в большинстве случае обеспечивают полное восстановление структур. В ЦНС, напротив, интенсивность процессов восстановления не высока, и регенерация не является полной.
а) Валлерова (антероградная) дегенерация периферических нервов. Основные этапы процесса дегенерации и их описание представлены на рисунке ниже и в подписи к нему. После разрушения или перерезки нерва в течение первых 48 ч аксоны и миелиновая оболочка под действием протеаз, высвобождающихся из шванновских клеток под влиянием ионов Са 2+ , распадаются на фрагменты элипсоидной формы дистальнее места нарушения целостности нерва. Продукты распада удаляют моноциты, которые перемещаются из кровотока в эндоневрий и превращаются в макрофаги.
Помимо осуществления фагоцитоза, макрофаги стимулируют митотическую активность шванновских клеток. Макрофаги и шванновские клетки выполняют трофическую (питательную) и направляющую функции для регенерирующих аксонов.
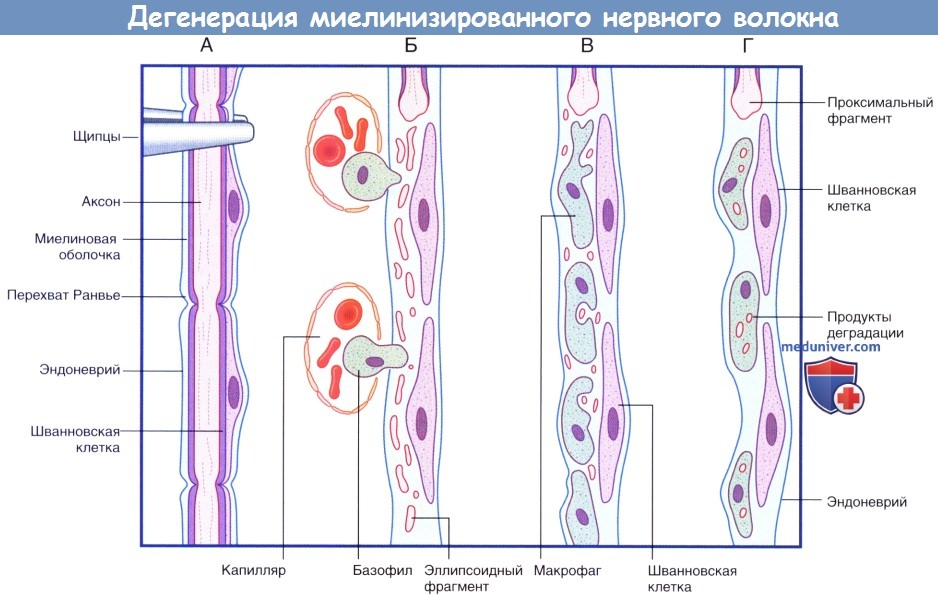
Последовательность процессов при дегенерации миелинизированного нервного волокна.
(А) Сохранное нервное волокно; продемонстрированы четыре сегмента. Выполняют пережатие нервного волокна в верхней части.
(Б) Миелиновая оболочка и аксон распадаются на мелкие частицы и фрагменты эллипсоидной формы. Моноциты проникают в эндоневрий из кровеносного русла.
(В) Мелкие продукты дегенерации поглощают моноциты.
(Г) Происходит практически полное удаление всех продуктов дегенерации. Шванновские клетки и эндоневрий сохранны. Последовательность процессов при регенерации миелинизированного нервного волокна. 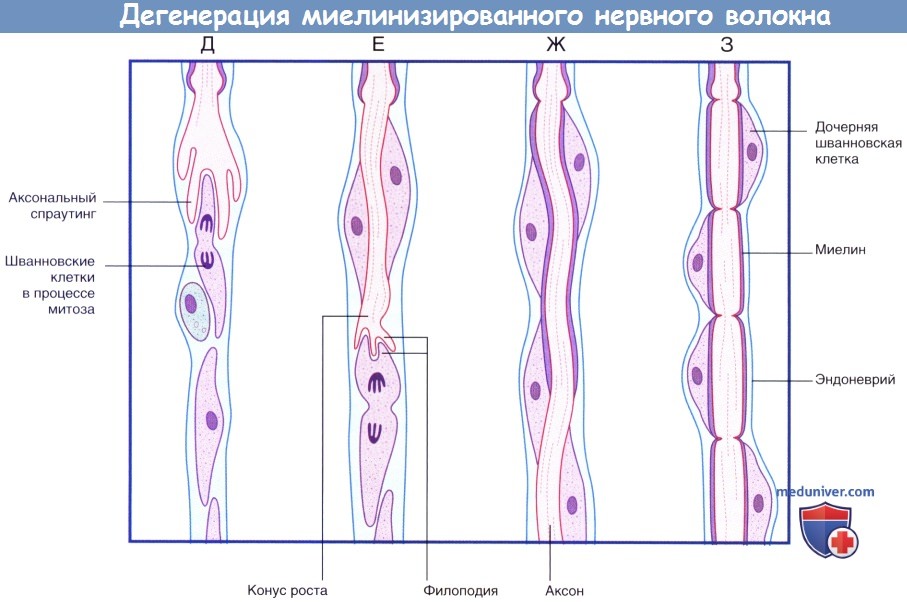
(продолжение) Последовательность процессов при дегенерации миелинизированного нервного волокна.
(Д) Аксональный спраутинг начинается с дистального участка аксона. Спраутинг оказывает митогенное дейсвтие на окружающие шванновские клетки.
(Е) Конус роста продвигается дистально вдоль поверхности шванновских клеток.
(Ж) Миелинизация начинается вдоль проксимального участка регенерирующего аксона.
(З) Общее строение вновь миелинизированного нервного волокна соответствует тем же принципам, однако миелинизированные сегменты характеризуются меньшей длиной.
б) Регенеративные процессы в периферической нервной системе. Основные этапы процесса регенерации представлены на рисунке ниже. После ровного среза нерва спраутинг (разрастание новых ветвей) на конце проксимального отрезка аксона начинается уже спустя несколько часов. Однако в клинической практике повреждения нерва часто происходят при раздавливании или разрыве. В этих случаях происходит отмирание участка нерва длиной 1 см и более, за счет чего спраутинг может продолжаться в течение недели. В случае удачной регенерации происходит тесное соприкосновение проксимального конца аксона со шванновской клеткой дистального конца пересеченного нейрона.
При нарушении формирования этой связи в месте первичного повреждения образуется псевдоневрома, представляющая собой извитые регенерирующие аксоны, погруженные в рубцовую ткань. Ампутационные псевдоневромы — источники сильных болей после ампутаций конечностей.
Регенерация нейронов при повреждении происходит двумя путями в течение нескольких часов после повреждения. На проксимальном конце пересеченного аксона появляются множественные отростки, на конце которых образуются утолщения — конусы роста. На дистальном конце шванновские клетки формируют отростки, направляющиеся навстречу конусам роста. На концах конусов роста формируются напоминающие антенны филоподии, где располагаются поверхностные рецепторы, временно связывающиеся с соответствующими поверхностными молекулами адгезии базальных мембран шванновских клеток. Актиновые филаменты филоподий прикрепляются к поверхностным рецепторам и относительно этих соединений осуществляют дальнейшее продвижение конусов роста.
Конусы роста стимулируют митотическую активность шванновских клеток. Шванновские клетки делятся и миелинизируют наиболее крупные аксоны.
При полном разрыве нервных стволов перед попыткой их восстановления, как правило, выжидают около трех недель, поскольку сразу после повреждения их соединительнотканные оболочки отечны, а в течение этого промежутка времени они становятся немного толще, что позволяет шовному материалу лучше закрепиться. Кроме того, обрезание нервов перед наложением швов приводит к развитию вторичной аксотомии проксимальной части пересеченного аксона. В ходе экспериментальных исследований на животных показано, что вторичная аксотомия стимулирует более интенсивную и длительную регенерацию.
Влияния тела нейрона на пересеченный участок нерва заключаются в следующем:
• Вследствие осмотических изменений перикариона ядро ориентируется эксцентрически.
• Клетки нейроглии заполняют все синаптические щели, изолируя исходный двигательный нейрон от синаптических контактов в сером веществе.
• В исследованиях, проведенных на обезьянах, показано, что после перерезки чувствительных нейронов 30-40 % окончаний дорсальных нервных корешков подвергается валлеровской дегенерации. Регенерация этих нейронов не происходит, поскольку их окончания располагаются в сером веществе головного мозга. Однако некоторые рецепторы восстанавливаются за счет спраутинга сохранных прилежащих нейронов. Данное наблюдение позволяет объяснить явление неполного восстановления чувствительности у таких пациентов.
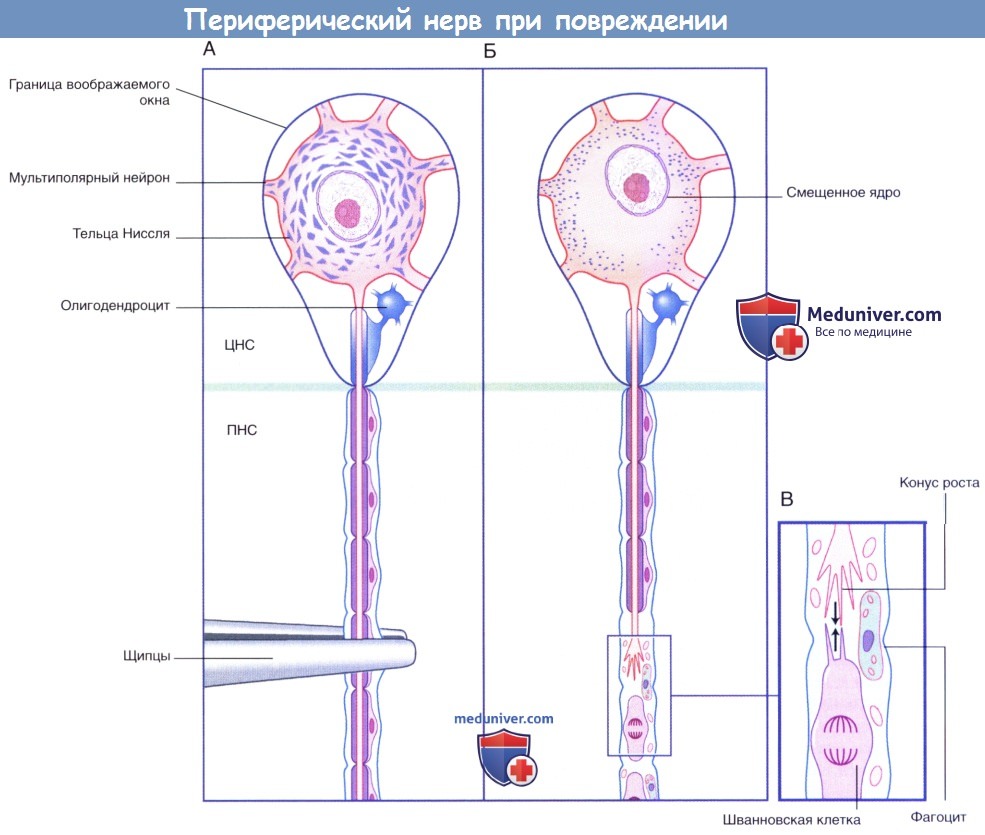
Схематическое изображение процессов, происходящих в периферическом нерве после повреждения.
(А) Двигательный нейрон ЦНС, видимый через воображаемое окно.
(Б) Хроматолизис проявляется фрагментированием и рассеиванием телец Ниссля, а также смещением ядра.
(В) В зоне повреждения в условиях удаления продуктов деградации происходит образование контакта филоподий конуса роста с проксимальными выростами шванновских клеток (указано стрелками).
ЦНС — центральная нервная система; ПНС — периферическая нервная система.
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Нейроны являются несменяемой клеточной популяцией. Им свойственна только внутриклеточная физиологическая регенерация, заключающаяся в непрерывной смене структурных белков цитоплазмы. Отростки нейронов и соответственно периферические нервы обладают способностью к регенерации в случае их повреждения. При этом регенерации нервных волокон предшествуют явления дегенерации.
Дегенерация
Дегенерация - это патологический процесс, характеризующийся распадом миелиновой оболочки нервных волокон на ограниченных участках при относительной сохранности осевых цилиндров.
При дегенерации нерва происходят биохимические и биофизические изменения. Вес нерва и содержание в нем воды увеличиваются, двоякопреломляемость нарушается, понижается концентрация липоидов мякотной оболочки (сфигномиелина, цереброзидов, свободного холестерина). Изменяется активность различных ферментов.
Шванновская оболочка при дегенерации нерва также изменяется. Уже через 48 часов после травматизации нерва в шванновских клетках происходит набухание ядра, увеличение хроматина, разрастание цитоплазмы. Через 5—6 дней становятся заметны митозы, образуются синцитиальные протоплазматические тяжи с несколькими ядрами, так называемые бюнгеровские ленты. В конце первой недели полибласты скапливаются на поверхности дегенерирующих нервных волокон, затем внедряются в них, превращаясь в макрофаги.
При валлеровской дегенерации распад периферического отрезка поврежденного нерва начинается одновременно на всем протяжении (Б. С. Дойников, Р. Кахаль). Мнения авторов, изучавших процессы дегенерации в периферическом отрезке нерва, о непосредственной причине распада различны.
При повреждении тела нервной клетки она обычно погибает и фагоцитируется микроглиальными элементами. Фагоциты – клетки микроглии, которые обладают способностью поглощать погибшие части нейронов. Если повреждается (в результате перетяжки, травмы и пр.) аксоннервной клетки, то в теле соответствующего нейрона наступает ряд характерных изменений. Во-первых, наблюдаетсяхроматолиз, т.е. разрушение и растворение субстанции Ниссля, представляющей собой шероховатую эндоплазматическую сеть со скоплением рибосом в теле нейрона. Одновременно вследствие потери воды размеры тела нервной клетки и её ядра могут уменьшаться, цитоплазма вакуолизируется, ядро занимает краевое положение и меняет форму. Число нейрофибрилл в клетке уменьшается, они делаются тоньше и плохо различимыми. Центральный и периферический отрезки перерезанного аксона, его мякотная и безмякотная оболочки претерпевают распад; на некотором расстоянии от места поврежедения миелин растворяется. Вся эта картина получила название “первичной реакции Ниссля”, или ретроградной клеточной дегенерации, а для центрального и периферического отрезка аксона – травматической дегенерации. Особенно сложно протекают изменения в периферическом отрезке перерезанного аксона или, если речь идёт о нерве, в периферическом отрезке нерва. Эти изменения называются вторичной, или валлеровской, дегенерацией нервных волокон. Во время валлеровской дегенерации периферические отрезки аксонов, потерявшие связь с телом нервной клетки, являющейся их трофическим центром, распадаются и полностью дегенерируют. Миелиновая оболочка распадается; миелин собирается в капли, в которых иногда ещё можно проследить обломки периферических аксонов.
Аксональная дегенерация возникает, если в теле клетки нарушается синтез необходимых аксону веществ или при нарушении выработки энергии в митахондриях, вследствие чего прекращается быстрый антероградный аксоплазматический ток в аксоне. При аксональной дегенерации в нейроне наблюдается повышенный уровень кальция, который активизирует систему кальпаинов, способных запускать различные патологические процессы.
При аксональной дегенерации, прежде всего, страдает утолщенная часть аксона, затем дегенерация распространяется в проксимальном направлении. Сильнее всего повреждаются самые длинные аксоны, поэтому симптомы аксональной дегенерации ощущаются, прежде всего, в стопах и в кистях.
Дегенеративный процесс, распространившийся на тело клетки, приводит к ее гибели. Если же воздействие факторов, приведших к аксональной дегенерации, прекратилось до гибели клетки, то происходит восстановление аксонов. Регенерация происходит медленно и может занять от нескольких месяцев до года.
В отличие от кожной раны, которая заживает по первичному натяжению, если ее края привести в соприкосновение, место пореза нерва первичным натяжением не срастается, даже в тех случаях, когда наложены немедленно эпиневральные швы.
Срастание разорванных частей нерва и восстановление его функций происходит путем длительного процесса регенерации нервных волокон, причем новообразованные нервные волокна растут из центрального участка поврежденного нерва. Такой точки зрения придерживаются отечественные и зарубежные нейрогистологи (Б. С. Дойников, Б. И. Лаврентьев, Р. Кахаль, И. Ф. Иванов, А. Н. Голиков). Немногочисленные представители теории клеточных цепей допускают возможность аутогенной регенерации за счет дифференцировки элементов шванновского синцития периферического отрезка.
Экспериментальные исследования процесса регенерации у некоторых лабораторных животных дали более обнадеживающие результаты. Было установлено, что у крыс после полной перерезки спинного мозга со временем восстанавливаются двигательные рефлексы в газовых конечностях (Л. Фримен, 1955). При дегенерации нерва происходят биохимические и биофизические изменения. Вес нерва и содержание в нем воды увеличиваются, двоякопреломляемость нарушается, понижается концентрация липоидов мякотной оболочки (сфигномиелина, цереброзидов, свободного холестерина). Изменяется активность различных ферментов.
Шванновская оболочка при дегенерации нерва также изменяется. Уже через 48 часов после травматизации нерва в шванновских клетках происходит набухание ядра, увеличение хроматина, разрастание цитоплазмы. Через 5—6 дней становятся заметны митозы, образуются синцитиальные протоплазматические тяжи с несколькими ядрами, так называемые бюнгеровские ленты. В конце первой недели полибласты скапливаются на поверхности дегенерирующих нервных волокон, затем внедряются в них, превращаясь в макрофаги.
При валлеровской дегенерации распад периферического отрезка поврежденного нерва начинается одновременно на всем протяжении (Б. С. Дойников, Р. Кахаль). Мнения авторов, изучавших процессы дегенерации в периферическом отрезке нерва, о непосредственной причине распада различны.
Патогистологические изменения в нервных волокнах изучены наиболее детально, они проявляются в двух основных формах: валлеровская, или вторичная, дегенерация, характеризующаяся распадом осевого цилиндра и мякотной оболочки, и периаксональная, или сегментарная, дегенерация (демиелинизация), когда преимущественно поражается мякотная оболочка, в то время как в осевом цилиндре патологических изменений нет.
Характер действующей причины, ее сила и длительность в одних случаях могут привести к одновременному сочетанию обеих описанных форм изменений нервного волокна или только преимущественно к одной из них, поэтому представляет большой интерес изучение изменений как в мякотной оболочке, так и в осевом цилиндре.
Изменения мякотной оболочки характеризуются распадом ее (демиелинизация), увеличением количества эльцгольцевых телец. Уже через 1—2 суток в мякотной оболочке появляются различной глубины поперечные зазубршш, в течение 3—5-го дня оболочка распадается на фрагменты, принимающие элипсоидную, шаровидную или овоидную формы, в дальнейшем распадаясь на мелкие зерна.
В осевых цилиндрах (аксоны) обычно происходит истончение или, напротив, утолщение, набухание, вакуолизация, разволокнение, варикозные вздутия, распад на фрагменты и зерна. Фрагменты распадающегося осевого цилиндра сильнее, чем обычно, начинают импрегнироваться серебром и нередко закручиваются в виде отдельных спиралей. Если в результате травмы нерва произошел перерыв аксона, то обычно на проксимальном его конце образуется шаровидное утолщение или он разволакнивается и отрывается от нервного волокна. В некоторых случаях при поражении периферических нервов в осевых цилиндрах появляются амилоидные тельца (А. В. Романов, В. В. Семенова-Тяншанская).
При валлеровском перерождении (Валлер, Б. С. Дойников), возникшем в результате травмы, изменения можно отметить уже в течение первых суток. Очертания осевого цилиндра становятся неровными, импрегнация серебром неравномерной, появляются продольная исчерченность и разволокнение отдельных аксонов их, варикозные вздутия. При исследовании в поляризационном свете изменения в осевом цилиндре обнаруживаются уже через 3 часа. Через 1—2 суток происходит распад на отдельные фрагменты различной длины, резкая аргентофилия, изменения калибра. Безмякотные нервные волокна распадаются на мелкие зерна уже в первые дни, но часть их остается в неизменном виде иногда до двух недель. При механическом повреждении нерва следует различать: изменения центрального отрезка нерва (ретроградное перерождение); изменения периферического отрезка (вторичное, или валлеровское, перерождение); местный процесс в зоне повреждения нерва, сопровождающийся перерождением осевых цилиндров и оболочек (первичная дегенерация).
В зависимости от тяжести ранения первичная дегенерация может проявляться только распадом миелиновой оболочки. При ранениях, нарушающих целостность осевых цилиндров, все поврежденные волокна, отделенные от своих нервных центров, подвергаются вторичному, или валлеровскому, перерождению. Сущность валлеройского перерождения сводится к разрушению осевых цилиндров; контуры их становятся неравномерными, изменяется калибр, нейрофибриллы варикозно утолщаются и разволакниваются. Через 5—6 дней осевые цилиндры распадаются на фрагменты, образуя зернистую массу; параллельно идет распад миелина на глыбки. Дегенерация начинается не во всех волокнах одновременно, несколько позже она происходит в мякотных волокнах, еще позднее в безмякотных, являющихся проводниками вегетативной нервной системы (Б. С. Дойников). В то время как аксоны и мякотные оболочки распадаются, шванновские клетки разрастаются. Из их протоплазмы образуются синцитиальные тяжи с ядрами. Разрастающиеся шванновские синцитии прорастают в рубцовую ткань, образовавшуюся на месте ранения, и проникают в центральный отрезок нерва. Дефект между центральным и периферическим отрезками заполняется тяжами из шванновских клеток, последние вместе с макрофагами фагируют обломки осевых цилиндров и миелина. Длительность процесса уборки зависит от калибра мякотных волокон; чем крупнее нервный ствол, тем дольше идет очищение бывшего ложа от продуктов распада. В крупных нервных стволах этот процесс иногда длится несколько месяцев.
Восстановление функции нервных волокон происходит путем длительного и сложного процесса регенерации. Срастания нервных волокон первичным натяжением не происходит. Непременным условием нормальной регенерации является соединение или приближение друг к другу поврежденных участков нерва наложением эпиневральных швов и отсутствие гнойного воспаления в зоне повреждения.
Новообразованные нервные волокна вначале не имеют миелиновой оболочки., Одновременно с миелинизацией начинается восстановление функции нерва. Первой восстанавливается болевая и грубая температурная чувствительность, позже тактильная, тонкая температурная и поверхностная болевая чувствительность. Последняя часто вовсе не восстанавливается. В некоторых случаях невротизация идет за счет нервных образований мышц и сосудов окружающих тканей. В таких случаях даже при выраженной регенерации функция нерва не восстанавливается. Собственные исследования показали, что у телят после перерезки шейного симпатического ствола с наложением эпиневральных швов исчезновение признаков птоза и восстановление двигательной функции верхнего века — симптомы регенерации нерва — наблюдаются через три недели.
Клиникофизиологическими признаками прорастания нервных волокон в периферический отрезок и восстановления биоэлектрического контакта с нервным центром являются: исчезновение анестезии, появление болевой чувствительности ниже места повреждения, восстановление мышечного тонуса, прекращение секреторных и трофических расстройств.
При открытых повреждениях нервных стволов регенеративный процесс может резко нарушаться. Разрыв нерва со значительным расхождением концов или препятствий между этими концами в виде грубых рубцов, осколков костей, инородных тел и других образований обычно влекут за собой образование травматической невромы. Она может возникнуть и при сохранении целости нерва, если имеются препятствия для растущих нервных волокон. В таких случаях происходит избыточное образование нервных волокон, растущих из центрального отрезка; они формируют запутанный клубок, обрастающий соединительной тканью (неврома).
Поскольку новообразование нервных волокон происходит длительное время, невромы могут значительно увеличиваться в объеме. Они создают патологическую болевую импульсацию, резко нарушающую двигательную и трофическую функции (ирритатинный фокус). После ампутации конечности, осложнившейся нагноением культи или развитием раневой инфекции, образование невром не составляет большой редкости (ампутационная неврома). Чтобы избежать развития ампутационной невромы, необходима тщательная асептика при операции и технически правильная обработка места пересечения нервного ствола. Рекомендуется накладывать лигатуру на предварительно оттянутый к периферии эпинервий. Это препятствует росту аксонов, и неврома не образуется.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Глия.
Каждая клетка ЦНС окружается протоплазматическими астроцитами с цитоплазмой, содержащей малое количество фибриллярных нитей. Волокна нервных клеток в белом веществе окружены фиброзными астроцитами, в цитоплазме которых присутствует большое количество фибриллярного материала. Фиброзные астроциты заполняют пространство между пучками миелизированных нервных волокон. Эти крупные клетки в составе глии похожи на раскрывшиеся бутоны астр, отсюда и их название – астроциты.
Олигодендроциты родственны астроцитам, но отличаются меньшими размерами и более мелкими ядрами, а так же более слаборазвитыми ветвистыми отростками. Они связаны непосредственно с телами нейронов и нервными волокнами, поэтому их часто рассматривают в качестве центральных гомологов шванновских клеток. Мелкие микроглиальныеклетки похожи на паучков. Они отличаются характером своих отростков и очень небольшими темными ядрами. Эти клетки равномерно рассеяны как в головном, так и спинном мозге.
Особое место в нервной ткани занимает эпендимный призматический эпителий – нейроэпителий, выстилающий спинномозговой канал и желудочки головного мозга. У эмбрионов и новорожденных он несёт мерцательные реснички.
Что касается крупных сосудов, которые находятся в нервной ткани, то они всём своём протяжении сопровождаются соединительной тканью и покрыты глиальными, образованными астроцитами пограничными мембранами, которые некоторыми исследователями рассматриваются в качестве одного из субстратов гематоэнцефалического барьера, обеспечивающего избирательную проницаемость сосудов мозга. Лимфатические сосуды нервной ткани отсутствуют.
СТРУКТУРА ГЛИАЛЬНЫХ КЛЕТОК.
Глиальные клетки были впервые выделены в определенную группу элементов нервной системы в 1871 г. А.Вирховым, который рассматривал своеобразную соединительную ткань мозга. Назвал эти клетки нейроглией, т.е. нервным клеем.
Выделяют 4 типа глиальных клеток: астроциты, олигодендроциты, клетки эпиндемы и микроглии Первые три разновидности глиальных клеток образуются в эмбриогенезе, как и нейрон из нейроэктодермы, микроглия занимает несколько обособленное положение.
1. Астроцитарная глия – это крупные клетки со светлым овальным ядром, многочисленными отростками небольшим числом органоидов.
2. Олигодендроциты – это глиальные клетки. К ней относятся: олигодендроциты серого и белого вещества мозга, шванновские клетки, клетки-спутники /сателлитная глия/. Характеризуются более плотной цитоплазмой, с хорошо развитым ЭПР, ап. Гольджи, множеством митохондрии, лизосом.
3. Эпендимная глия является разновидностью глиальных клеток. Она образует выстилку полостей мозговых желудочков и центрального канала спинного мозга. Представлена цилиндрическими и кубовидными клетками. Хорошо развиты органоиды клеток.
4. Микроглия это мелкие отростчатые клетки с очень плотной цитоплазмой. Характерен фагоцитоз. До сих пор окончательно не решен вопрос о происхождении микроглии в эмбриогенезе. С одной стороны ее рассматривают как своеобразные макрофаги, и таким образом, относят к элементам тканей внутренней среды мезенхимного происхождения. С другой стороны имеются данные, позволяющие рассматривать часть микроглии как недифференцированные /покоящееся/ астроциты, которые при определенных условиях начинают активно размножаться и превращаться в зрелые фиброзные астроциты.
Глия выполняет следующие функции.
1. Обеспечивают нормальную деятельность определенных нейронов и всего мозга.
2. Обеспечение надежной элементарной изоляции тел нейронов, их отростков, синапсов для исключение неадекватного взаимодействие между нейронами при распространении возбуждении по нейронным цепям мозга.
3. 3.Астроциты и олигодендроциты обладают способностью активно захватывать из синаптической щели медиаторы или их составные части после прекращение синаптической передачи. В частности, целиком захватывается глией такие медиаторы как КА, аминокислотные пептиды.
4. Трофическая функция глий. В глиальных клетках сосредоточен основной запас гликогене /главного энергетического субстрата мозга/ и липидов. Они контролируют ионный состав межклеточной жидкости, обеспечивает стабильность внутренней среды мозга – необходимое условие нормального функционирования нервной ткани.
Преобразовавшись в пучки усиленно растущих волокон, регенерирующие центральные отрезки аксонов в конце концов проникают в бюнгнеровы тяжи и начинают расти в них, как по готовому руслу, достигая старых периферических нервных чувствительных или двигательных окончаний. Шванновский синцитий распадается на отдельные клетки, в которых появляется миелин с характерными перехватами Ранвье и т.п. Аналогичным путём идёт врастание регенерирующих волокон и в безмякотных нервах, но без образования миелина. Одновременно восстанавливаются и функции регенерировавших нервов.
Регенеративные процессы в ЦНС во многих отношениях остаются ещё не изученными, хотя частичное или полное функциональное восстановление при травмах центральной нервной системы имеет место в ряде случаев. В ЦНС к регенерации отрезанных отростков способны клетки Гольджи 1-го типа с длинными аксонами. Клетки Гольджи 2-го типа с короткими отростками, по- видимому, не способны к восстановлению утраченных отростков. Однако и в случае регенерации последняя носит абортивный характер, т.к. полному восстановлению перерезанных аксонов мешает сложный соединительнотканноглиальный рубец, возникающий на месте травмы или пере резки. В последнее время в опытах на млекопитающих, задерживая рост глиальной части рубца подкожным введением животному пиромена, удавалось наблюдать регенерацию некоторых перерезанных нервных пучков спинного и головного мозга.
Особый интерес представляет проблема образования раковых опухолей в нервной системе. Содержательно этот процесс представляет патологические изменения, происходящие в клетках нервной ткани, приводящие к непрерывному их делению. Никаких других своих функций такая клетка не выполняет, только делится. Причём скорость деления раковых клеток потрясающая. Вновь образованные клетки заполняют собой все нервные пути, ткани и органы, препятствуя их нормальному функционированию, и сами продолжают процесс деления. Что служит толчком к началу процесса непрерывного деления нервных клеток, пока точно не известно, как и то, что может остановить этот уже начавшийся процесс. Имя того человека, кто даст ответы на эти вопросы и решит проблему борьбы с раковыми клетками, будет золотыми буквами записано в истории человечества и на доске почёта в каждом медицинском учреждении, работающим в этом направлении. Может это будет Ваше имя? Будем надеяться!
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Хотя установлено, что нейроны могут делиться в головном мозгу взрослых птиц, нейроны у млекопитающих обычно не делятся, а их дегенерация приводит к необратимым потерям. Отростки нейронов в центральной нервной системе в очень ограниченных пределах могут замещаться благодаря синтетической активности их перикарионов.
Нервные волокна периферической нервной системы также способны регенерировать, если их перикарионы не разрушены.
Гибель нервной клетки ограничена ее перика-рионом и отростками. Нейроны, функционально связанные с погибшим нейроном, не погибают, за исключением тех, которые имеют только одну связь. В последнем случае изолированный нейрон претерпевает изменения, описанные как транснейронная дегенерация.
В отличие от нервных клеток, нейроглия центральной нервной системы, а также шванновские клетки и сателлитные клетки узлов периферической нервной системы способны делиться митозом. В центральной нервной системе в пространства, которые остаются после нервных клеток, погибших в результате заболевания или повреждения, внедряется нейроглия.
Так как нервы широко распределены по всему телу, они часто повреждаются. При перерезке аксона происходят дегенеративные изменения, после которых наступает фаза репарации. В поврежденном нервном волокне важно различать изменения,происходящие в проксимальном и дистальном сегментах. Проксимальный сегмент сохраняет свою связь с трофическим центром (перикарионом) и часто регенерирует. Дисталь-ный сегмент, отделенный оттела нервной клетки, дегенерирует.
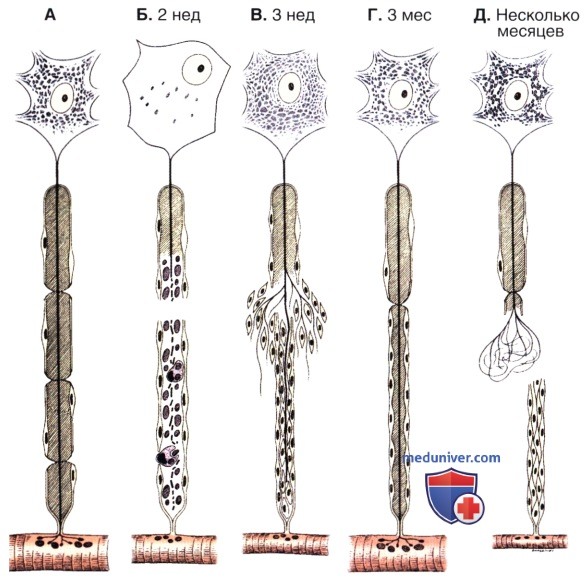
Важнейшие изменения, происходящие в поврежденном нервном волокне.
А — нормальное нервное волокно с его перикарионом и эффекторным органом (поперечнополосатой скелетной мышцей). Обратите внимание на расположение ядра нейрона, а также на количество и распределение телец Ниссля.
Б — при повреждении волокна ядро нейрона смещается к периферии клетки, а количество телец Ниссля резко сокращается. Нервное волокно дистальнее участка повреждения дегенерирует вместе со своей миелиновой оболочкой. Детрит фагоцитируется макрофагами.
В — мышечное волокно подвергается выраженной атрофии вследствие денервации. Шванновские клетки пролиферируют, образуя компактный тяж, в который внедряется растущий аксон. Аксон растет со скоростью 0,5—3 мм/сут.
Г — в данном случае регенерация нервного волокна прошла успешно. Обратите внимание, что мышечное волокно также регенерировало после возобновления поступления нервной стимуляции.
Д — в том случае, если аксон не проникает сквозь тяж шванновских клеток, его рост не имеет организованного характера.
Повреждение аксона вызывает ряд изменений в перикарионе: хроматолиз, т.е. растворение субстанции Ниссля с последующим уменьшением базофилии цитоплазмы, увеличение объема перикариона и миграцию ядра к его периферии. Проксимальный сегмент аксона вблизи раны дегенерирует на небольшом протяжении, однако как только макрофаги удалят детрит, начинается его рост. Макрофаги вырабатывают интерлейкин-1, стимулирующий шванновские клетки, которые секретируют вещества, обеспечивающие рост нерва.
В оставшейся части нервного волокна, дистальнее места повреждения, как аксон (отделенный от своего трофического центра), так и миелиновая оболочка полностью дегенерируют, а их остатки, за исключением соединительнотканной и периневральной оболочек, удаляются макрофагами. В то время как происходят эти регрессивные изменения, шванновские клетки пролиферируют внутри оставшегося тяжа из соединительной ткани, давая начало солидным клеточным колонкам.
Эти ряды шванновских клеток служат в качестве направляющих элементов для пускающих ростки аксонов, которые формируются в течение репаративной фазы.
После регрессивных изменений проксимальный сегмент аксона растет и ветвится, образуя несколько филаментов, которые продолжают двигаться в направлении колонок из шванновских клеток. Только те волокна, которые проникнут сквозь эти колонки шванновских клеток, будут расти и достигнут эффекторного органа.
В том случае, если между дистальным и проксимальным сегментами имеется значительный промежуток или когда дистальный сегмент исчезает полностью (как, например, при ампутации конечности), из появившихся в результате роста новых нервных волокон может образоваться опухоль, или неврома, которая способна стать источником спонтанных болей.
Регенерация функционально эффективна только тогда, когда нервные волокна и колонки шванновских клеток растут в правильном направлении. Вероятность этого, однако, достаточно велика, поскольку каждое регенерирующее волокно дает начало нескольким отросткам, а к каждой колонке шванновских клеток направляются отростки от нескольких регенерирующих волокон. В поврежденном смешанном нерве, однако, если регенерирующие чувствительные волокна врастут в колонки, связанные с моторными бляшками, которые ранее занимали двигательные волокна, восстановления функции мышцы не произойдет.
Читайте также:
