
Филогенез покровов тела и нервной системы хордовых
Филогенез покровов тела хордовых животных
Наружные покровы, или кожа, подразделяются на 2 слоя: эпидермис (эктодермального происхождения) и кориум – дерма, или собственно кожа (мезодермального происхождения). Покровы тела выполняют следующие функции:
- защитная (от воздействия факторов внешней среды);
- участие в выделении продуктов обмена (мочевины, минеральных солей);
- орган чувств (содержит рецепторы осязания, температурные, болевые);
- участие в дыхании (амфибии).
У ланцетника покровы тела слабо развиты: эпидермис однослойный с отдельными железистыми клетками; кориум – слой студенистой соединительной ткани (рисунок 52).

Рисунок 52 – Строение кожи ланцетника
1 – кутикула, 2 – эпидермис, 3 – кориум
Направления эволюции покровов тела:
- дифференцировка на эпидермис и дерму, увеличение толщины дермы;
- от однослойного эпидермиса к многослойному;
- дифференцировка производных кожи;
- от одноклеточных кожных желез к многоклеточным;
- развитие подкожной жировой клетчатки и совершенствование механизмов терморегуляции.
- чешуя рептилий, перья птиц, волосы млекопитающих;
- железистые клетки рыб, многоклеточные железы наземных позвоночных (сальные, потовые, молочные);
- когти, ногти, рога, копыта.
Онтофилогенетически обусловленные аномалии развития кожных покровов у человека (рекапитуляции и параллелизмы): отсутствие потовых желез, ихтиоз (усиленное развитие рогового слоя), многососковость (в норме у человека два соска, число сосков у животных от 1-8 до 22-25).
Филогенез скелета хордовых
Осевой скелет ланцетника – хорда(энтодермального происхождения) и плотные тяжи, которые дают опору плавникам и жаберным щелям. Хорда – это упругий тяж из особых вакуолизированных клеток.
Скелет позвоночных мезодермального происхождения. Он состоит из трех частей: осевой скелет, скелет головы, скелет конечностей и их поясов.
- защитная (для внутренних органов):
- участие в обмене веществ (депо солей кальция и фосфора).
Направления эволюции осевого скелета:
- замена хорды позвоночником, хрящевой ткани костной тканью;
- дифференцировка позвоночника на отделы, увеличение числа позвонков в каждом отделе;
- формирование грудной клетки.
Круглоротые и низшие (хрящевые) рыбы сохраняют хорду, в течение всей жизни. Над хордой и под хордой в виде парных хрящей появляются дуги позвонков.
У высших (костных) рыб позвоночник имеет два отдела – туловищный и хвостовой. Позвонки имеют тела, дуги и отростки. В туловищном отделе есть ребра, которые свободно заканчиваются на брюшной стороне тела.
Отделы позвоночника амфибий: шейный и крестцовый отделы имеют по 1 позвонку, грудной отдел – 5 позвонков, хвостовой отдел содержит разное число позвонков. Шейный и грудной отделы несут короткие ребра, которые заканчиваются свободно. Грудина хрящевая.
Рептилии имеют 5 отделов позвоночника. Появляется поясничный отдел. Количество позвонков в отделах позвоночника увеличивается до 8-12. В шейном отделе ребра редуцируются, в поясничном отделе имеются рудиментарные ребра. Хорошо развитые ребра грудного отдела соединяются с грудиной, образуя грудную клетку. Хвостовой отдел рептилий содержит различное число позвонков.
У млекопитающих позвоночник состоит из 5 отделов: шейный отдел (7 позвонков), грудной отдел (9 – 24 позвонка), поясничный отдел (2 – 9 позвонков), крестцовый отдел (4 – 10 и более позвонков), хвостовой отдел (различное число позвонков). Ребра редуцированы в шейном и поясничном отделах. 10 пар ребер вместе с костной грудиной образуют грудную клетку.
Онтофилогенетические аномалии осевого скелета у человека:
- дополнительные (шейные) ребра;
- увеличение позвонков хвостового отдела (длина хвоста до 20 см);
- увеличение числа крестцовых позвонков;
- расщепление задних отростков позвонков.
Эволюция скелета конечностей. Основой для образования конечностей позвоночных животных послужили метаплевральные (кожные) складки, расположенные по бокам туловища у ланцетника.
Метаплевральные складки ланцетника → парные (грудные и брюшные) плавники рыб → пятипалая конечность наземного типа.
- уменьшение числа костных элементов плавников и укрупнение их путем слияния;
- увеличение подвижности в сочленении с плечевым поясом.
Конечности рыб – это грудные и брюшные плавники. Внутренний скелет плавников составляют хрящевые или костные лучи, неподвижно соединенные между собой и с плечевым поясом. У большинства рыб плавники служат для изменения направления движения. У кистеперых рыб плавники стали использоваться для передвижения по грунту, и сочленение с поясом конечностей стало подвижным.
Конечности пятипалого типа появились у амфибий. Прочное соединение элементов в плавниках заменилось подвижным сочленением в виде суставов. Конечность превратилась в сложный рычаг. Удлиняются кости плеча, предплечья и пальцев. Пояса конечностей (лопатки, вороньи кости, ключицы) имеют вид дуги, которая охватывает тело с боков и снизу. Хорошо развит спинной отдел поясов.
Онтофилогенетически обусловленные аномалии развития скелета конечностей у человека:
- добавочные кости запястья и предплюсны;
- полидактилия (более 5 пальцев);
- синдактилия (срастание фаланг пальцев);
- брахидактилия (укорочение фаланг пальцев).
Скелет головы (череп) позвоночных состоит из мозгового и лицевого (висцерального) отделов. Черепная коробка защищает головной мозг и органы чувств. Лицевой череп образует опору для передней части пищеварительного тракта. Закладка мозгового отдела у зародышей всех позвоночных происходит под головным мозгом в виде двух пар хрящей. По бокам переднего конца хорды расположены удлиненные пластинки – парахордалии, впереди от хорды лежит пара хрящей – трабекулы. Парахордалии и трабекулы сливаются, образуя дно и боковые части черепной коробки. К ней прирастают обонятельные и слуховые капсулы, с боков – глазничные хрящи (рисунок 53).

Рисунок 53 – Распределение хрящевых закладок черепа акулы
(вид со спинной стороны). 1 – хорда, 2 – парахордалии, 3 – трабекулы, 4 – обонятельные капсулы, 5 – слуховая капсула, 6 – глазничные хрящи, 7 – глаз.
Направления эволюции черепа:
- замена хрящевых элементов костными;
- уменьшение числа костей за счет их слияния;
- изменение формы (преобладание мозгового отдела над лицевым).
Висцеральный отдел закладывается из метамерно расположенных вокруг переднего конца пищеварительной трубки хрящевых дуг.
Направления эволюции висцерального отдела – дифференциация висцеральных дуг.
У рыб 1-я дуга (челюстная) состоит из двух хрящей – небно-квадратного и меккелева, которые срастаются и выполняют функции челюстей (рисунок 54). 2-я дуга (подъязычная) содержит гиомандибулярный хрящ, выполняющий функцию прикрепления к черепной коробке (гиостильный череп). Остальные дуги – жаберные – являются опорой для дыхательного аппарата.

Рисунок 54 – Хрящевой скелет акулы
1 – осевой череп, 2 – небноквадратный хрящ, 3 –нижний хрящ I – й дуги, 4 –гиомандибулярный хрящ, 5 – гиоид, 6 – VI – X жаберные дуги
У наземных позвоночных:
- челюстная дуга срастается с дном черепа;
- подъязычная дуга дает начало слуховым косточкам;
- 3-6-я жаберные дуги дают начало щитовидному хрящу и другим хрящам гортани.
Для всех наземных позвоночных характерно подвижное соединение мозгового черепа с позвоночником (аутостильный тип черепа).
У млекопитающих уменьшается число костей мозгового черепа за счет их слияния и увеличивается его объем. Нижняя челюсть соединяется с мозговым черепом с помощью сустава. Размеры лицевого черепа значительно меньше размеров мозгового черепа.
Онтофилогенетически обусловленные аномалии развития скелета головы у человека:
- увеличение числа костных элементов;
- одна или две слуховые косточки;
ФИЛОГЕНЕЗ ПОКРОВОВ ТЕЛА ХОРДОВЫХ
Кожные покровы хордовых имеют эктодермальное (эпидермис) и мезодермальное (дерма) происхождение. В связи с разной средой обитания и образом жизни кожные покровы хордовых очень разнообразны по строению и отличаются многообразием функций.
• защищает организм от вредных воздействий факторов внешней среды (механических, химических, проникновения микроорганизмов);
• участвует в терморегуляции;
• является органом осязания (осязательные, температурные и болевые рецепторы);
• у некоторых животных участвует в дыхании.
В связи с разной средой обитания и образом жизни, кожа и ее производные у хордовых разнообразны по строению и функциям.
Основные направления эволюции покровов хордовых:
1) дифференцировка на два слоя: наружный – эпидермис, внутренний – дерму и увеличение толщины дермы;
1) от однослойного эпидермиса к многослойному;
2) дифференцировка дермы на 2 слоя – сосочковый и сетчатый;
3) появление подкожно-жировой клетчатки и совершенствование механизмов терморегуляции;
4) от одноклеточных желез к многоклеточным;
6) дифференцировка различных производных кожи.
У низших хордовых (ланцетник) эпидермис однослойный, цилиндрический, имеет железистые клетки, выделяющие слизь. Дерма (кориум) представлена тонким слоем неоформленной соединительной ткани.
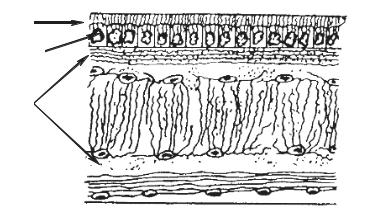
Рис. 1. Разрез через кожу ланцетника. 1 – кутикула, 2 –
эпидермис, 3 – кориум
У низших позвоночных эпидермис становится многослойным. Нижний его слой – ростковый (базальный), клетки его делятся и восполняют клетки вышележащих слоев. Дерма имеет правильно расположенные волокна, сосуды и нервы.
Производными кожи являются : одноклеточные (у круглоротых, рыб) и многоклеточные (у земноводных) слизистые железы; чешуя: а) плакоидная у хрящевых рыб, в развитии которой принимают участие эпидермис и дерма; б) костная у костных рыб, которая развивается за счет дермы.
Плакоидная чешуя снаружи покрыта слоем эмали (эктодермального происхождения), под которым находятся дентин и пульпа (мезодермального происхождения). Чешуя и слизь выполняют защитную функцию.
У земноводных кожа тонкая гладкая, без чешуй. Кожа содержит большое количество многоклеточных слизистых желез, секрет которых увлажняет покровы и обладает бактерицидными свойствами. Кожа принимает участие в газообмене.
У высших позвоночных в связи с выходом на сушу эпидермис становится сухим, имеет роговой слой.
У рептилий развиваютсяроговыечешуи, отсутствуюткожныежелезы.
У млекопитающих : хорошо развиты эпидермис и дерма, появляется подкожно-жировая клетчатка.
Эпидермис состоит из 5 слоев. Каждый слой характеризуется особенностями строения и выполняет свои функции. В коже находится большое количество желез: потовые, сальные, млечные, пахучие и др. Все они
– производные эпидермиса вторично погруженные в дерму. Имеются также разнообразные производные рогового слоя: волосы, рога, когти, ногти, копыта. Волос состоит из стержня и корня. Корень расположен в волосяной сумке, в которую открываются протоки сальных желез. В дерме выделяют сетчатый и сосочковый слои. Сосочковый слой содержит нервные рецепторы, кровеносные и лимфатические сосуды.
3
5 
Рис. 2. Строение кожи млекопитающих. 1 – эпидермис, 2 – дерма, 3 –
ороговевающий слой, 4 – ростковый слой, 5 – сосочковый слой, 6 – сетчатый слой, 7 – потовая железа, 8 – нервные окончания, 9 – подкожная жировая клетчатка, 10 – волос, 11 – сальная железа, 12 – мышца, поднимающая волос
У человека встречаются онтофилогенетически обусловленные аномалии кожного покрова (механизм возникновения – рекапитуляции): отсутствие потовых желез, ихтиоз (чрезмерное развитие рогового слоя), избыточная обволошенность лица и тела, дополнительные млечные железы (многососковость) и др.
Филогенез систем органов хордовых рассмотрен в соответствии с прогрессивным направлением эволюции этого типа животных от подтипа Бесчерепные до класса Млекопитающие. Организация систем органов класса Птицы не описана в связи с тем, что птицы произошли от пресмыкающихся значительно позже млекопитающих и являются боковой ветвью эволюции хордовых.
Наружные покровы
Покровы любых животных всегда выполняют функцию восприятия внешних раздражений, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции сопровождается дифференцировкой. Характерно также расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное — экто- и мезодермальное — происхождение. Из эктодермы развивается эпидермис, из мезодермы — дерма. Для бесчерепных характерна слабая степень дифференцировки обоих слоев кожи. Эпидермис — однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях — дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
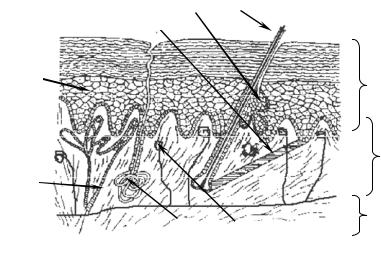
У рыб в эпидермисе железы одноклеточные. Они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной. Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью (рис. 15.1). Дентин — мезодермального происхождения, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся снаружи в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.

Рис. 15.1. Закладка плакоидной чешуи.
1 — клетки-эмалеобразователи, 2 — эпидермис, 3 — эмаль, 4 — склеробласты — ден- тинообразователи, 5 — дентин, 6 — сосочек дермы
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуй в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных — стегоцефалов — соответствовала покровам рыб и тоже была покрыта чешуей. Современные амфибии имеют тонкую гладкую кожу без чешуй, принимающую участие в газообмене. Этому способствует наличие большого количества многоклеточных слизистых желез, секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы — продуценты токсинов, защищающих их от врагов (см. разд. 23.1).
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает. Роговые чешуйки у некоторых рептилий тонки и эластичны, у других — сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие — насекомоядные, грызуны и некоторые другие — наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3—7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос (рис. 15.2), покрывающих практически все тело, кроме некоторых участков, например подошв и ладоней у приматов.

Рис. 15.2. Расположение волос у млекопитающих. А — на хвосте грызунов;
Б — на других участках тела.
1 — роговые чешуйки, 2 — группы волос, расположенные в шахматном порядке
Волосы многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы, основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функции передних конечностей, у многих яйцекладущих и сумчатых — разбросаны по всему телу. Этот факт может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.

Рис. 15.3. Строение развивающегося соска млекопитающего.
Потовые железы млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы.
Виден постепенный переход от потовых (2) к млечным (2) железам
У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам (рис. 15.3). Количество млечных

А — зародыш в возрасте 5 недель (млечные линии); Б —дифференцировка пяти пар сосков; В — зародыш в возрасте 7 недель
Рис. 15.4. Эмбриогенез передней брюшной стенки человека.


Рис. 15.5. Атавистические аномалии развития кожи.
А — гипертрихоз; Б — полимастия
Сальные железы образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуй, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез) (рис. 15.5). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами — млекопитающими. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.
1 Лекция 19 Тема: Филогенез систем органов хордовых животных (I) План лекции 1. Связь онтогенеза и филогенеза. Биогенетический закон. 2. Филэмбриогенезы. 3. Способы органогенеза. Корреляции. 4. Филогенез покровов и скелета хордовых животных. Связь онтогенеза и филогенеза. Биогенетический закон Онтогенез это индивидуальное развитие, комплекс процессов развития отдельной особи от образования зиготы и до смерти. Развитие происходит благодаря реализации генетической информации, полученной от родителей. Существенное влияние на ее реализацию оказывают условия окружающей среды. Филогенез это историческое развитие вида, эволюционное развитие организмов. Онтогенез и филогенез тесно связаны. Зная направления и преобразования органов и их систем в процессе исторического развития, можно понять и объяснить возникающие в процессе эмбриогенеза аномалии развития. Связь онтогенеза и филогенеза получила отражение в ряде биологических законов и закономерностей. В 1828 году Карл Бэр сформулировал три закона. Первый закон закон зародышевого сходства зародыш какого-либо высшего животного не бывает похож на другое животное, но похож на его эмбрион. Второй закон закон последовательного появления признаков более общие признаки, характерные для данной крупной группы животных, выявляются у их зародышей раньше, чем признаки более специальные. Третий закон закон эмбриональной дивергенции каждый зародыш данной формы животных не проходит через другие формы, а постепенно обособляется от них. Объяснить эти законы можно так. На ранних этапах эмбриогенеза зародыши животных разных классов позвоночных (например, рыбы, птицы,
2 млекопитающие) похожи между собой. Со временем между ними появляются различия в пределах классов, а дальше в пределах отрядов (пример: зародыш свиньи и зародыш человека). Закон параллелизма Меккеля-Серре говорит о том, что каждое существо в своем эмбриональном развитии повторяет взрослые формы животных более низких ступеней развития. Ч. Дарвин, подтверждая связь онтогенеза и филогенеза, создал учение о рекапитуляциях. Рекапитуляция это повторение у зародышей признаков их предков. Например, в процессе закладки и развития дыхательной системы у зародыша млекопитающего проходят этапы жаберных отверстий на глотке, затем образуются жабры, а затем органы наземного дыхания легкие. В 1866 году Э.Геккель сформулировал биогенетический закон: онтогенез есть краткое и быстрое повторение филогенеза, но повторяется строение не взрослых стадий предков, а их эмбриональных стадий. Оказалось, что рекапитуляции в онтогенезе могут быть не всегда. На этом основании Геккель выделил две группы признаков палингенезы и ценогенезы. Палингенезы это повторение у зародышей признаков предков (закладка хорды и жаберных щелей на глотке, двухкамерного сердца у наземных позвоночных). Ценогенезы это эмбриональные адаптации, которые имеются только у зародышей и отсутствуют у взрослых организмов (зародышевые оболочки у высших позвоночных амнион, хорион, аллантоис). Филэмбриогенезы Важное значение для объяснения связи онтогенеза и филогенеза имеет учение А.Н.Северцова о филэбриогенезах. Филэмбриогенезы это эмбриональные новообразования, которые имеют филогенетическое значение. Время их появления и способы могут быть различными. Пример: из чешуи акулы развиваются: а) роговые щитки рептилий путем девиации; б) перо птицы путем анаболии; в) волос млекопитающих путем архаллаксиса. Эволюция чаще идет путем анаболий, поэтому наблюдаются рекапитуляции.
3 Время появления филэмбриогенезов начальные стадии морфогенеза средние стадии морфогенеза конечные стадии морфогенеза Архаллаксис полностью изменяется развитие органа (рекапитуляция отсутствует) Девиация отклонение в развитии органа (рекапитуляция частичная в начальных стадиях) Анаболия надставка в развитии органа имеет место рекапитуляция, новый признак закладывается на поздней стадии развития) Способы органогенеза. Корреляции. Главные принципы эволюционных преобразований органов: Дифференциация Интеграция Подразделение первоначально однородной структуры Усиление взаимной зависимости частей организма. Например: пищеварительная трубка Например: взаимозависимость дыхательной млекопитающих подразделяется на: ротовую и кровеносной систем млекопитающих. полость, глотку, пищевод, желудок, кишечник. А.Н.Северцов описывает основные способы морфофункциональных изменений органов в филогенезе. Наиболее часто в процессе эволюции наблюдается смена функций органов. Одновременно со сменой функций происходит изменение строения органа. Например: плавательный пузырь кистеперых рыб преобразовался в орган наземного дыхания легкие; кожная чешуя рыб в процессе эволюции преобразовалась в зубы млекопитающих; из яйцеклада членистоногих образовалось жало. Следующим способом является расширение функций в связи с изменяющимися условиями. Грудные плавники рыб вначале выполняли функции обеспечения устойчивости тела в воде на определенном уровне, затем они стали определять направление движения. У донных рыб они выполняют функции опоры и передвижения по дну водоема. В некоторых случаях отмечается усиление функций органа. Увеличиваются размеры органа, и изменяется его гистологическая структура. Пример: развитие переднего мозга у позвоночных животных.
4 Филогенетические преобразования могут происходить путем замещения органов и функций. Один орган исчезает или становится рудиментарным, а замещает его другой орган. Пример: замена хорды позвоночником, замена головной почки у позвоночных туловищной, а потом тазовой. Принцип компенсации функций: мускулистый (мышечный) желудок птиц компенсирует отсутствие у них зубов; участие кожи амфибий в дыхании компенсирует недостаточное легочное дыхание. Взаимоотношения между частями организма, согласованность филогенетических преобразований его органов и частей называются корреляциями. Различают корреляции геномные, морфогенетические и эргонтические, или функциональные. Корреляции геномные определяются взаимодействием генов, плейотропией, сцеплением генов. Например: сочетание волос светлых и гладких, темных и курчавых; редукция крыльев у дрозофилы и одновременное укорочение задней пары конечностей. Корреляции морфогенетические определяются внутренними факторами на ранних стадиях эмбрионального развития, когда еще не установились функциональные связи между частями зародыша. В основе этих корреляций лежит эмбриональная индукция зачатки структур, образующиеся раньше, определяют направление и характер изменений зачатков, образующихся позже. Например: дорзальная губа бластопора индуцирует развитие нервной трубки, хорды и сомитов. Корреляции эргонтические определяются взаимозависимостью между функциями определенных органов. Например, связь между гонадотропной функцией гипофиза и развитием половых желез. Филогенез покровов тела хордовых животных Наружные покровы, или кожа, подразделяются на 2 слоя: эпидермис (эктодермального происхождения) и кориум дерма, или собственно кожа (мезодермального происхождения). Покровы тела выполняют следующие функции: защитная (от воздействия факторов внешней среды);
5 участие в выделении продуктов обмена (мочевины, минеральных солей); орган чувств (содержит рецепторы осязания, температурные, болевые); депо крови; участие в дыхании (амфибии). У ланцетника покровы тела слабо развиты: эпидермис однослойный с отдельными железистыми клетками; кориум слой студенистой соединительной ткани (рис. 48) Рис.48. Строение кожи ланцетника 1 кутикула, 2 эпидермис, 3 кориум Направления эволюции покровов тела: дифференцировка на эпидермис и дерму, увеличение толщины дермы; от однослойного эпидермиса к многослойному; дифференцировка производных кожи; от одноклеточных кожных желез к многоклеточным; развитие подкожной жировой клетчатки и совершенствование механизмов терморегуляции. Производные кожи: чешуя рептилий, перья птиц, волосы млекопитающих; железистые клетки рыб, многоклеточные железы наземных позвоночных (сальные, потовые, молочные); когти, ногти, рога, копыта. Онтофилогенетически обусловленные аномалии развития кожных покровов у человека (рекапитуляции и параллелизмы): отсутствие потовых
6 желез, ихтиоз (усиленное развитие рогового слоя), многососковость (в норме у человека два соска, число сосков у животных от 1-8 до 22-25). Филогенез скелета хордовых Осевой скелет ланцетника хорда (энтодермального происхождения) и плотные тяжи, которые дают опору плавникам и жаберным щелям. Хорда это упругий тяж из особых вакуолизированных клеток. Скелет позвоночных мезодермального происхождения. Он состоит из трех частей: осевой скелет, скелет головы, скелет конечностей и их поясов. Функции скелета: опорно-двигательная; защитная (для внутренних органов): кроветворная; участие в обмене веществ (депо солей кальция и фосфора). Направления эволюции осевого скелета: замена хорды позвоночником, хрящевой ткани костной тканью; дифференцировка позвоночника на отделы, увеличение числа позвонков в каждом отделе; формирование грудной клетки. Круглоротые и низшие (хрящевые) рыбы сохраняют хорду, в течение всей жизни. Над хордой и под хордой в виде парных хрящей появляются дуги позвонков. У высших (костных) рыб позвоночник имеет два отдела туловищный и хвостовой. Позвонки имеют тела, дуги и отростки. В туловищном отделе есть ребра, которые свободно заканчиваются на брюшной стороне тела. Отделы позвоночника амфибий: шейный и крестцовый отделы имеют по 1 позвонку, грудной отдел 5 позвонков, хвостовой отдел содержит разное число позвонков. Шейный и грудной отделы несут короткие ребра, которые заканчиваются свободно. Грудина хрящевая. Рептилии имеют 5 отделов позвоночника. Появляется поясничный отдел. Количество позвонков в отделах позвоночника увеличивается до В шейном отделе ребра редуцируются, в поясничном отделе имеются
7 рудиментарные ребра. Хорошо развитые ребра грудного отдела соединяются с грудиной, образуя грудную клетку. Хвостовой отдел рептилий содержит различное число позвонков. У млекопитающих позвоночник состоит из 5 отделов: шейный отдел (7 позвонков), грудной отдел (9-24 позвонка), поясничный отдел (2-9 позвонков), крестцовый отдел (4-10 и более позвонков), хвостовой отдел (различное число позвонков). Ребра редуцированы в шейном и поясничном отделах. 10 пар ребер вместе с костной грудиной образуют грудную клетку. Онтофилогенетические аномалии осевого скелета у человека: дополнительные (шейные) ребра; увеличение позвонков хвостового отдела (длина хвоста до 20 см); увеличение числа крестцовых позвонков; расщепление задних отростков позвонков. Эволюция скелета конечностей. Основой для образования конечностей позвоночных животных послужили метаплевральные (кожные) складки, расположенные по бокам туловища у ланцетника. Метаплевральные складки ланцетника парные (грудные и брюшные) плавники рыб пятипалая конечность наземного типа. Направления эволюции: уменьшение числа костных элементов плавников и укрупнение их путем слияния; увеличение подвижности в сочленении с плечевым поясом. Конечности рыб это грудные и брюшные плавники. Внутренний скелет плавников составляют хрящевые или костные лучи, неподвижно соединенные между собой и с плечевым поясом. У большинства рыб плавники служат для изменения направления движения. У кистеперых рыб плавники стали использоваться для передвижения по грунту, и сочленение с поясом конечностей стало подвижным. Конечности пятипалого типа появились у амфибий. Прочное соединение элементов в плавниках заменилось подвижным сочленением в виде суставов. Конечность превратилась в сложный рычаг. Удлиняются кости плеча, предплечья и пальцев. Пояса конечностей (лопатки, вороньи
8 кости, ключицы) имеют вид дуги, которая охватывает тело с боков и снизу. Хорошо развит спинной отдел поясов. Онтофилогенетически обусловленные аномалии развития скелета конечностей у человека: добавочные кости запястья и предплюсны; полидактилия (более 5 пальцев); синдактилия (срастание фаланг пальцев); брахидактилия (укорочение фаланг пальцев). Скелет головы (череп) позвоночных состоит из мозгового и лицевого (висцерального) отделов. Черепная коробка защищает головной мозг и органы чувств. Лицевой череп образует опору для передней части пищеварительного тракта. Закладка мозгового отдела у зародышей всех позвоночных происходит под головным мозгом в виде двух пар хрящей. По бокам переднего конца хорды расположены удлиненные пластинки парахордалии, впереди от хорды лежит пара хрящей трабекулы. Парахордалии и трабекулы сливаются, образуя дно и боковые части черепной коробки. К ней прирастают обонятельные и слуховые капсулы, с боков глазничные хрящи (рис. 49) Рис. 49. Распределение хрящевых закладок черепа акулы (вид со спинной стороны) 1 - хорда, 2 - парахордалии, 3 - трабекулы, 4 - обонятельные капсулы, 5 - слуховая капсула, 6 - глазничные хрящи, 7 глаз. Направления эволюции черепа: замена хрящевых элементов костными; уменьшение числа костей за счет их слияния;
9 увеличение объема; изменение формы (преобладание мозгового отдела над лицевым). Висцеральный отдел закладывается из метамерно расположенных вокруг переднего конца пищеварительной трубки хрящевых дуг. Направления эволюции висцерального отдела дифференциация висцеральных дуг. У рыб 1-я дуга (челюстная) состоит из двух хрящей небно-квадратного и меккелева, которые срастаются и выполняют функции челюстей. 2-я дуга (подъязычная) содержит гиомандибулярный хрящ, выполняющий функцию прикрепления к черепной коробке (гиостильный череп). Остальные дуги жаберные являются опорой для дыхательного аппарата Рис. 50. Хрящевой скелет акулы 1 осевой череп, 2 небноквадратный хрящ, 3 нижний хрящ I й дуги, 4 гиомандибулярный хрящ, 5 гиоид, 6 VI X жаберные дуги У наземных позвоночных: челюстная дуга срастается с дном черепа; подъязычная дуга дает начало слуховым косточкам; 3-6-я жаберные дуги дают начало щитовидному хрящу и другим хрящам гортани. Для всех наземных позвоночных характерно подвижное соединение мозгового черепа с позвоночником (аутостильный тип черепа). У млекопитающих уменьшается число костей мозгового черепа за счет их слияния и увеличивается его объем. Нижняя челюсть соединяется с мозговым черепом с помощью сустава. Размеры лицевого черепа значительно меньше размеров мозгового черепа. Онтофилогенетически обусловленные аномалии развития скелета головы у человека:
Читайте также:
