
Физиологические свойства нерва и мышцы
В этой книге предельно сжато изложен курс лекций по нормальной физиологии. Благодаря четким определениям основных понятий студент может сформулировать ответ, за короткий срок усвоить и переработать важную часть информации, успешно сдать экзамен. Курс лекций будет полезен не только студентам, но и преподавателям.
- ЛЕКЦИЯ № 1. Введение в нормальную физиологию
- ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
- ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
- ЛЕКЦИЯ № 4. Физиология мышц
- ЛЕКЦИЯ № 5. Физиология синапсов
- ЛЕКЦИЯ № 6. Физиология центральной нервной системы
- ЛЕКЦИЯ № 7. Физиология различных разделов ЦНС
Приведённый ознакомительный фрагмент книги Нормальная физиология: конспект лекций (С. С. Фирсова) предоставлен нашим книжным партнёром — компанией ЛитРес.
ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
1. Физиология нервов и нервных волокон. Типы нервных волокон
Физиологические свойства нервных волокон:
1) возбудимость – способность приходить в состояние возбуждения в ответ на раздражение;
2) проводимость – способность передавать нервные возбуждение в виде потенциала действия от места раздражения по всей длине;
3) рефрактерность (устойчивость) – свойство временно резко снижать возбудимость в процессе возбуждения.
Нервная ткань имеет самый короткий рефрактерный период. Значение рефрактерности – предохранять ткань от перевозбуждения, осуществляет ответную реакцию на биологически значимый раздражитель;
4) лабильность – способность реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом импульсов возбуждения за определенный период времени (1 с) в точном соответствии с ритмом наносимых раздражений.
Нервные волокна не являются самостоятельными структурными элементами нервной ткани, они представляют собой комплексное образование, включающее следующие элементы:
1) отростки нервных клеток – осевые цилиндры;
2) глиальные клетки;
3) соединительнотканную (базальную) пластинку.
Главная функция нервных волокон – проведение нервных импульсов. Отростки нервных клеток проводят сами нервные импульсы, а глиальные клетки способствуют этому проведению. По особенностям строения и функциям нервные волокна подразделяются на два вида: безмиелиновые и миелиновые.
Безмиелиновые нервные волокна не имеют миелиновой оболочки. Их диаметр 5–7 мкм, скорость проведения импульса 1–2 м/с. Миелиновые волокна состоят из осевого цилиндра, покрытого миелиновой оболочкой, образованной шванновскими клетками. Осевой цилиндр имеет мембрану и оксоплазму. Миелиновая оболочка состоит на 80 % из липидов, обладающих высоким омическим сопротивлением, и на 20 % из белка. Миелиновая оболочка не покрывает сплошь осевой цилиндр, а прерывается и оставляет открытыми участки осевого цилиндра, которые называются узловыми перехватами (перехваты Ранвье). Длина участков между перехватами различна и зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами. При диаметре 12–20 мкм скорость проведения возбуждения составляет 70—120 м/с.
В зависимости от скорости проведения возбуждения нервные волокна делятся на три типа: А, В, С.
Наибольшей скорость проведения возбуждения обладают волокна типа А, скорость проведения возбуждения которых достигает 120 м/с, В имеет скорость от 3 до 14 м/с, С – от 0,5 до 2 м/с.
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
- ЛЕКЦИЯ № 1. Введение в нормальную физиологию
- ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
- ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
- ЛЕКЦИЯ № 4. Физиология мышц
- ЛЕКЦИЯ № 5. Физиология синапсов
- ЛЕКЦИЯ № 6. Физиология центральной нервной системы
- ЛЕКЦИЯ № 7. Физиология различных разделов ЦНС
Приведённый ознакомительный фрагмент книги Нормальная физиология: конспект лекций (С. С. Фирсова) предоставлен нашим книжным партнёром — компанией ЛитРес.
Согласно современным представлениям, нервная и мышечная ткани могут находиться в трех основных состояниях — физиологическом покое, возбуждении и торможении.
Физиологический покой — это такое состояние, когда ткань или орган не проявляет признаков присущей им деятельности.
Возбуждение — деятельное состояние живой ткани, в которое она приходит под влиянием раздражения.
Торможение — такое состояние, когда деятельность ткани или органа ослабляется или полностью прекращается.
Раздражители.При раздражении мышца приходит в состояние возбуждения, которое распространяется по мышце и вызывает ее сокращение. По биологическому действию раздражители бывают адекватные и неадекватные. Адекватный — это такой раздражитель, к которому данный орган или ткань приспособились в процессе эволюции. Для мышцы адекватным раздражителем является нервный импульс. Неадекватными — будут такие раздражители, действию которых ткань или орган в естественных условиях обычно не подвергаются. Так, сокращение мышцы можно вызвать механическим раздражением (укол, кислота, ток) и т.д.
Все раздражители по своей силе деляг на пороговые, подпороговые и сверхпороговые. Пороговыми называют минимальные раздражители, которые могут вызвать возбуждение. Подпороговые — это раздражители, сила которых меньше пороговой, сверхпороговые — раздражители более сильные, чем пороговые.
Биоэлектрические явления.Возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки (рис. 4.19).
Потенциал покоя.Мембрана нервной или мышечной клетки в состоянии покоя с наружной стороны заряжена положительно, а с внутренней — отрицательно. Разность зарядов между наружной и внутренней поверхностью клеточной мембраны в состоянии физиологического покоя клетки называется потенциалом покоя, или мембранным потенциалом.
Потенциал действия. Под влиянием раздражения проницаемость клеточной мембраны для ионов натрия повышается в сотни
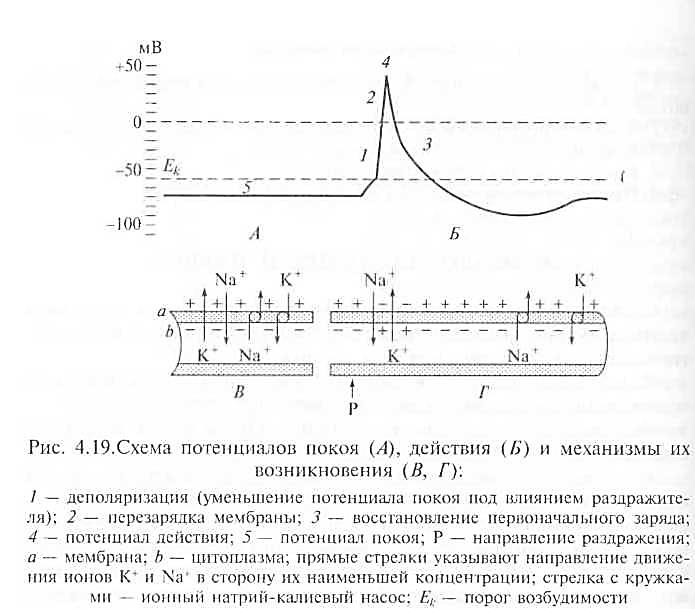 |
Рис. 4.19.Схема потенциалов покоя (А), действия (5) и механизмы их
возникновения (В, Г):
1 — деполяризация (уменьшение потенциала покоя под влиянием раздражите-
ля); 2 — перезарядка мембраны; 3 — восстановление первоначального заряда;
4 — потенциал действия; 5 — потенциал покоя; Р — направление раздражения;
а — мембрана; b — цитоплазма; прямые стрелки указывают направление движе-
ния ионов К + и в сторону их наименьшей концентрации; стрелка с кружка-
ми — ионный натрий-калиевый насос; Ек — порог возбудимости
раз. Происходит перезарядка клеточной мембраны: внутренняя ее сторона заряжается положительно, а наружная — отрицательно. Затем значительно повышается проницаемость мембраны для ионов калия, они начинают быстро перемещаться из клеток во внеклеточную жидкость и вновь восстанавливают первоначальный заряд мембраны. Пикообразное колебание потенциала, возникающее в результате кратковременной перезарядки мембраны и последующего восстановления ее заряда, называется потенциалом действия. Такие колебания потенциала продолжаются 1 — 5 мс.
Проведение возбуждения. Круговой ток, проходя через участок покоя, раздражает его и вызывает появление потенциала действия (рис. 4.20). Когда в одном из участков нервной клетки, нервного или мышечного волокна возникает возбуждение и появляется потенциал действия, между возбужденным и соседним невозбужденным участком возникает разность потенциалов. Эта разность потенциалов является причиной появления круговых (локальных) токов. Внутри клетки ток идет от возбужденного участка к участку покоя, по внешней стороне — от участка покоя к возбужденному.
В участке, который ранее был возбужден, происходит восста
Рис. 4.20. Схема распрост-
ранения возбуждения:
А — волокна мышц и безмякотных нервных волокон; Б —
мякотное нервное волокно; 1 — мембрана; 2 — волокно;
а, Ь, с — перехваты Ранвье; заштрихованы возбужденные
участки; светлые — участки покоя; стрелками показано движение круговых токов

новление потенциала покоя. Данный процесс многократно повторяется и обусловливает распространение импульсов возбуждения. Скорость проведения возбуждения по скелетным мышцам 12 —15 м/с, гладким мышцам — 1 — 18 см/с, в безмякотных нервах — 0,5 — 3 м/с.
Основные свойства живой ткани.Любая живая клетка обладает свойствами раздражимости, возбудимости и лабильности (функциональной подвижности).
Раздражимость. С раздражимостью связаны все процессы роста и размножения клеток и тканей. На разнообразные воздействия внешней или внутренней среды — раздражения протоплазма живой клетки отвечает специфической реакцией, при которой происходят усиление или ослабление обмена веществ, количественное и качественное его изменение. Эта реакция на раздражение получила название раздражимости, или реактивности.
Возбудимость — свойство нервной или мышечной ткани отвечать на действие раздражителя возбуждением. Для измерения величины возбудимости определяют порог возбудимости и хронаксию. Порогом возбудимости называется наименьшая сила раздражителя, способная вызвать возбуждение; его выражают в вольтах постоянного тока.
Лабильность — скорость, с которой в ткани возникает и успевает закончиться полный период отдельного импульса возбуждения. Мера лабильности — это максимальное число импульсов возбуждения за 1 св ответ на такое же максимальное число раздражений. Высокой лабильностью обладают мякотные нерпы, у без- мякотных нервов она низкая. Так, максимальный ритм возбуждений мякотного нервного волокна 500, безммкотного — 200. Максимальный ритм возбуждений скелетной мышцы 200 импульсов в 1 с, а гладкой мышцы — в десятки раз меньше.
При очень частых раздражениях сокращения мышцы уменьшаются или даже совсем прекращаются. Такая частота называется пессимальной, или пессимумом. Пессимум возникает вследствие того, что возбуждение еще не закончилось и ткань находится в состоянии абсолютной или относительной рефрактерное™, а на нее действует новое раздражение. Частые раздражения, превышающие меру лабильности, вызывают не возбуждение, а торможение.
Учение Н. Е. Введенского о парабиозе устанавливает связь между возбуждением и торможением. Возбуждение и торможение — это различные реакции ткани на раздражение, исход которогозависит от лабильности. При высокой лабильности возникает возбуждение, снижение лабильности вызывает торможение.
ФИЗИОЛОГИЯ МЫШЦ
Скелетные мышц, входящие в состав опорно-двигательного аппарата позвоночных животных, состоят из группы мышечных пучков, каждый из них составлен из тысяч мышечных волокон, которые представляют собой клетки цилиндрической формы длиной до 12 см и диаметром 10 — 100 мкм. Каждое волокно окружено оболочкой сарколеммой и содержит тонкие нити — миофибрил-лы (рис. 4.21). Поперечные мембраны делят каждую миофибриллу на отдельные участки — саркомеры. Сократимым веществом мышечного волокна являются миофибриллы, состоящие из множества (около 2500) тонких и толстых белковых нитей — протофибрилл. Толстые протофибриллы образованы из белка миозина, тонкие — из актина. Нити актина прикреплены к мембране саркомера; они образуют светлые участки миофибриллы. В темных участках находятся нити миозина. Акгиновые нити частично входят своими концами в промежутки между миозиновыми нитями. Нити актина и миозина соединяются между собой многочисленными поперечными мостиками, которые образованы скрученными в спираль отростками — мостиками миозиновой нити. Чередование нитей в миофибрилле обусловливает ее поперечную исчерченность.

Рис. 4.21. Схема чередова-
ния нитей в миофибрилле:
А — электронно-микроскопическая фотография мио-
фибриллы; Б — строение миофибриллы; Z — поперечная
мембрана, разделяющая миофибриллу на отдельные уча-
стки — саркомеры; 1 — миозиновая нить, 2 — актиновая нить
Гладкие мышцы, образующие стенки внутренних органов, состоят из клеток различной формы длиной от 30 до 500 мкм и диаметром 2—10 мкм; у клеток имеются боковые отростки, благодаря которым клетки гладких мышц группируются в длинные пучки. Они, в свою очередь, при помощи тяжей соединяются друг с другом, обеспечивая деятельность мышцы как единой системы. Сократительным аппаратом гладких мышц являются миофибриллы, в которых содержатся тонкие актиновьге нити с прикрепленными к ним короткими отростками миозиновых нитей, называемых димерами.
Свойства скелетных мышц.Проведение возбуждения в мышцах происходит изолированно, т. е. потенциалы действия не переходят с одного мышечного волокна на другое. Возбудимость скелетныхмышц меньше возбудимости нервов. Нервное волокно оканчивается в середине мышечного волокна, поэтому возбуждение распространяется в обе стороны со скоростью 4—15 м/с.
Скелетная мышца является упругим телом. Если к мышце подвесить груз, то она растягивается, это свойство называется растяжимостью. Эластичностью мышцы называется возвращение мышцы к первоначальной своей длине после растяжения, когда будет удален груз. Пластичностью мышцы называется свойство сохранять удлиненную форму после удаления груза, вызвавшего ее растяжение. В скелетных мышцах лучше выражено свойство эластичности, чем пластичности.
Виды мышечных сокращений.При нанесении на мышцу одиночного раздражения она отвечает одиночным сокращением. Записывая это сокращение на двигающейся ленте какого-либо самописца (например, в простейшем случае на кимограф), можно отметить три периода: латентный — период от раздражения до начала сокращения, период сокращения и период расслабления. У икроножной мышцы лягушки одиночное сокращение длится 0,1 с, у млекопитающих — 0,04 — 0,1 с. В латентный период в мышце происходят процессы, при которых освобождается энергия для мышечного сокращения.
Если к мышце поступает несколько частых импульсов возбуждения, наступает длительное сокращение мышцы, которое называется тетаническим, или тетанусом. В зависимости от частоты возбуждений тетанус будет зубчатым или гладким (рис. 4.22).
Зубчатый тетанус наблюдается при такой частоте импульсов возбуждений, когда каждый импульс действует на мышцу в тот момент, когда она уже начинает расслабляться. Если же импульсы возбуждения настолько частые, что они воздействуют на мышцу до начала ее расслабления, то получатся длительные непрерывные сокращения мышцы — гладкий тетанус.
 |
Рис. 4.22. Сокращение скелетной мышцы при различной частоте раздра-
жения:
А — одиночное сокращение в ответ на одно раздражение; Б — зубчатый тетанус
(до 15 раздражений в 1 с); В — гладкий тетанус (свыше 25 раздражений); М —
механограмма — запись сокращения мышц; Р — частота раздражений; 1 — фаза
сокращения; 2 — фаза расслабления
Механизм мышечного сокращения (теория скольжения).В состоянии покоя тонкие нити актина лежат частично своими концами в промежутках между толстыми миозиновыми нитями. При возбуждении мышцы под влиянием потенциала действия нити актина сдвигаются в промежутке между нитями миозина, т. е. скользят навстречу друг другу, вызывая сокращение миофибрилл (рис. 4.23). Считают, что скольжение происходит в результате укорочения мостиков — боковых отростков миозиновых нитей под влиянием ионов кальция. Укорачиваясь, мостики тянут актиновую нить навстречу миозиновой.
Работа и сила мышц.При сокращении мышца укорачивается и тем самым совершает работу. Она будет наибольшей при средних нагрузках и среднем ритме сокращений.
Силу мышцы определяют по максимальному сокращению ее при поднятии максимального груза. Наибольшей силой обладают мышцы с косыми волокнами, имеющими перистое строение.
Утомление мышц.В процессе сокращений мышцы утомляются, при этом понижаются их возбудимость, лабильность и величина сокращения. В утомленной мышце понижается содержание гликогена и накапливаются продукты обмена. В опытах на нервно-мышечном препарате Н. Е. Введенский установил, что прежде всего утомляются синапсы в связи с их низкой лабильностью. В целом организме утомление в первую очередь наступает в нервных центрах коры больших полушарий. И. М.Сеченов доказал, что быстрое восстановление работоспособности утомленных мышц наступает не при полном их покое, а при работе других, до этого не сокращавшихся мышц. Импульсы от вновь вовлеченных в работу мышц повышают возбудимость нервных центров, возбуждение одних нервных центров снижает и даже снимает утомление других центров. Утомление зависит от состояния симпатической нервной системы и желез внутренней секреции: утомленная мышца вновь
Рис. 4.23. Схема расслабленной (А) и сократившейся (Б) миофибриллы
 |
начинает сокращаться при раздражении симпатического нерва или введении адреналина, усиливающих обмен. Тренировка мышц позволяет отсрочить наступление утомления. При ней увеличивается объем мышц в результате утолщения мышечных волокон, повышается содержание гликогена, АТФ и креатинфосфата, ускоряются восстановительные процессы.
Тонус мышц. Тонус скелетных мышц играет важную роль для поддержания определенного положения тела в пространстве и деятельности двигательного аппарата. Скелетные мышцы в покое расслабляются не полностью, а находятся в некотором напряжении, т.е. тонусе. Он обусловлен поступлением к мышцам редких нервных импульсов, возбуждающих мышечные волокна не одновременно, а поочередно.
Гладкие мышцы обладают способностью длительно находиться в тонусе. Длительные тонические сокращения этих мышц особенно отчетливо выражены в сфинктерах полых органов, в стенках кровеносных сосудов.
Одним из основных физиологических свойств возбудимых тканей является возбудимость, которая у различных тканей различна. Для характеристики уровня возбудимости служит порог раздражения, то есть минимальная сила раздражителя, при действии которой возникает ответная реакция. В экспериментальных условиях для определения возбудимости мышцы применяют прямой метод ее раздражения, то есть раздражение, наносимое непосредственно на мышцу. Возбудимость нерва исследуют раздражением нерва, иннервирующего данную мышцу, то есть методом непрямого раздражения мышцы.
Для работы необходимо: препаровальный набор инструментов, лоток, салфетки, операционный столик, Раствор Рингера, лягушка, кимограф, универсальный штатив с вертикальным миографом, стимулятор, раздражающие электроды, чернила, бумага.
Ход работы. Готовят нервно-мышечный препарат лягушки. Собрать установку по схеме, представленной на рисунке6.
Рис.6. Схема установки для регистрации мышечных сокращений: 1 – кимограф; 2 – вертикальный миограф; 3 – стимулятор.
Для графической регистрации движений икроножную мышцу укрепляют в миографе, для чего крючком, фиксированным на верхнем кронштейне миографа, прокалывают сумку коленного сустава, а пяточное сухожилие также с помощью крючка укрепляют на нижнем кронштейне миографа. К верхнему кронштейну миографа крепят раздражающие электроды с помощью специальной муфты, на которые помещают нерв нервно-мышечного препарата. Электроды соединены с клеммами стимулятора. Ручку регулировки амплитуды устанавливают на 0. Подавая на нерв одиночные стимулы с постоянной длительностью (например, 0,5 мс), постепенно увеличивают амплитуду и находят ту минимальную силу раздражителя, которая вызывает едва заметное сокращение мышцы – это и будет порог раздражения для нерва.
Для определения порога раздражения мышцы на нее наносят прямое раздражение через проводящую цепь миографа, соединенного со стимулятором. Порог раздражения находят так же, как при непрямом раздражении. Запись производят на ленте остановленного кимографа, поворачивая барабан рукой после каждой стимуляции.
Рекомендации к оформлению работы. Записать ход работы, зарисовать схему установки для прямого и непрямого раздражения мышцы. Записать результаты опыта, вклеить полученную кимограмму. Дать оценку возбудимости нерва и мышцы, сравнив пороги их возбуждения.
Физиология мышц
Вопросы для самоподготовки.1. Общие физиологические свойства мышц. 2. Виды сокращения мышц. 3. Механизм мышечного сокращения. 4. Физиологические особенности скелетных мышц. 5. Топография и физиологические свойства гладких мышц.
По морфологическим признакам выделяют три группы мышц: 1) поперечно-полосатые скелетные мышцы; 2) гладкие мышцы; 3) сердечную мышцу (миокард).
Функции поперечно-полосатых мышц: 1) двигательная (динамическая и статическая); 2) обеспечения дыхания; 3) мимическая; 4) рецепторная; 5) депонирующая; 6) терморегуляторная.
Функции гладких мышц: 1) поддержание давления в полых органах; 2) регуляция давления в кровеносных сосудах; 3) опорожнение полых органов и продвижение их содержимого.
Основная функция сердечной мышцы – насосная, обеспечение движения крови по сосудам.
Физиологические свойства скелетных мышц:
1) возбудимость (ниже, чем в нервном волокне);
2) проводимость (низкая, порядка 10-13 м/с);
3) рефрактерность (занимает по времени больший отрезок, чем у нервного волокна);
4) лабильность (функциональная подвижность, около 250 имп./с);
5) сократимость (способность укорачиваться или (и) развивать напряжение).
6) Эластичность (способность развивать напряжение при растягивании).
Структурно-функциональной единицей скелетных мышц является многоядерное мышечное волокно. Объединяясь в пучки, эти волокна образуют мышцу. Специфическим свойством мышц является сократимость. Мышечное сокращение проявляется в укорочении мышцы и (или) развитии ею механического напряжения. В зависимости от условий стимуляции и функционального состояния мышцы может возникнуть одиночное, слитное (тетаническое) сокращение или контрактура мышцы (стойкое длительное, иногда необратимое сокращение мышц, сохраняющееся после прекращения действия раздражителя – при отравлении некоторыми ядами, лекарственными средствами, нарушении обмена веществ, повышении температуры тела).
Мышечному сокращению предшествует процесс возбуждения, электрографическим проявлением которого является биопотенциал. По времени своего развития биопотенциал совпадает с латентным периодом мышечного сокращения. Амплитуда одиночного сокращения мышцы зависит от количества сократившихся миофибрилл. Возбудимость отдельных групп волокон, составляющих целую мышцу, различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения последовательно вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращений.

Рис.7 Скелетная мышца состоит из поперечнополосaтых мышечных волокон. Значительный объём МВ занимают миофибриллы. Расположение светлых и тёмных дисков в параллельных друг другу миофибриллах совпадает, что приводит к появлению поперечной исчерченности. Структурная единица миофибрилл — саркомер, сформированный из толстых (миозиновых) и тонких (актиновых) нитей. Расположение тонких и толстых нитей в саркомере показано слева и слева внизу. G‑актин — глобулярный, F‑актин — фибриллярный актин.
Физиологические особенности гладких мышц.Гладкие мышцы имеют те же физиологические свойства, что и скелетные, но имеют и свои особенности:
1) нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного частичного сокращения – тонуса;
2) самопроизвольную автоматическую активность;
3) сокращение в ответ на растяжение;
5) высокую чувствительность к химическим, в том числе фармакологическим веществам.
Дата добавления: 2018-02-28 ; просмотров: 860 ;
Теория по нормальной физиологии на тему: Синапс. Физиология мышечных волокон. В данной статье рассматривается механизм синаптической передачи, НМС…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Синапс — это специфическое место контакта двух возбудимых систем (клеток) для передачи возбуждения.
По способу передачи сигналов:
- механические,
- химические,
- электрические.
По виду медиатора: холинэргические и др.
Нервно-мышечный синапс (НМС) — химический, передача с помощью медиатора ацетилхолина.
Синонимы к слову НМС:
- Нервно-мышечное соединение;
- Моторная концевая пластинка.

Аксоны нервных клеток на своих окончаниях теряют миелиновую оболочку , ветвятся, и концевые веточки аксона утолщаются. Это пресинаптическая терминаль или бляшка или пуговка , которая погружается в углубление на поверхности мышечного волокна.
Покрывающая концевую веточку аксона поверхностная мембрана называется пресинаптической мембраной , т.е. это мембрана, покрывающая поверхность синаптической бляшки (терминали аксона).
Мембрана, покрывающая мышечное волокно в области синапса, называется постсинаптической мембраной , или концевой пластинкой. Она имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна, за счет чего увеличивается площадь контакта.

На постсинаптической мембране находятся белковые структуры — рецепторы , способные связывать медиатор . В одном синапсе количество рецепторов достигает 10-20 млн.
Между пре- и постсинаптическими мембранами находится синаптическая щель , размеры ее в среднем 50 нм, она открывается в межклеточное пространство и заполнена межклеточной жидкостью .
В синаптической щели находится мукополисахаридное плотное вещество в виде полосок, мостиков и содержится фермент ацетилхолинэстераза .
В пресинаптической терминали находится большое количество пузырьков или везикул , заполненных медиатором — химическим веществом посредником, осуществляющим передачу возбуждения.
В нервно-мышечном синапсе медиатор — ацетилхолин (АХ).

АХ синтезируется из холина и уксусной кислоты (ацетил-коэнзима А) с помощью фермента холинэстеразы. Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование АХ.
3 фракции медиатора:
- Первая фракция — доступная — располагается рядом с пресинаптической мембраной.
- Вторая фракция — депонированная — располагается над первой фракцией.
- Третья фракция — диффузно рассеянная — наиболее удаленная от пресинаптической мембраны.
Механизм синаптической передачи



Ионы Ca вызывают образование специального белкового комплекса , который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома .



Излитию содержимого пузырька в щель способствует белок синаптопорин , формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.





На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает ! Он формируется на соседней мембране мышечного волокна.
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе :
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП) .
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Физиология мышечных волокон
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл , расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин .
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации . Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
Сравнительная характеристика скелетных и гладких мышц:
Читайте также:
