
Химический состав нервной ткани миелиновые мембраны
Особенности структуры и функции основных элементов НС – нейрона и нейроглии
Нервная ткань состоит из многих типов клеток, обладающих разнообразными специфическими функциями. Это вносит определенные трудности в знакомство с основами молекулярных процессов, протекающих в нервной ткани. Можно выделить несколько основных задач, решение которых лежит в области биохимии нервной ткани:
а) изучение молекулярных основ проведения возбуждения по аксону;
б) изучение молекулярных основ синаптической передачи;
в) изучение особенностей метаболизма белков, липидов, углеводов, обеспечивающих выполнение основных функций нервной ткани;
г) изучение молекулярных основ обработки информации и ее хранения;
д) изучение молекулярных основ взаимодействия между клетками нервной ткани и других тканей организма;
На рис.18.1,18.2 и 18.3 представлен химический состав мозга. Основным его компонентом является вода, причем в сером веществе её больше, чем в белом. Сухой остаток представлен белками, липидами и минеральными веществами. Среди них в белом веществе мозга особенно много липидов - 17%. Доля белков в сером и белом веществе мозга приблизительно одинакова, она составляет, соответственно, 8 и 9%.
Таблица 18.1. Химический состав мозга человека(в %)
| Компоненты | Серое вещество | Белое вещество |
| Вода | ||
| Белки | ||
| Липиды | ||
| Минеральные вещества |
Таблица 18.2. Липиды серого и белого вещества мозга (в % от общего количества липидов)
| Название липида | Серое вещество | Белое вещество | Название липида | Серое вещество | Белое вещество |
| Холестерол | 22.0 | 27.5 | Фосфатидилэтаноламины | 22.7 | 14.9 |
| Галактолипиды | 7.3 | 26.4 | Фосфатидилхолины | 26.7 | 12.8 |
| Цереброзиды | 5.4 | 19.8 | Фосфатидилсерины | 8.7 | 7.9 |
| Сульфатиды | 1.7 | 5.4 | Фосфатидилинозиты | 2.7 | 0.9 |
| Общие фосфолипиды | 69.5 | 45.9 | Плазмалогены | 8.8 | 11.2 |
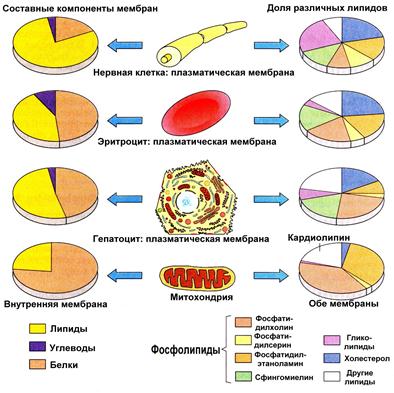
Рис.18.1. Сравнительный химический состав плазматической мембраны нейронов, эритроцитов, гепатоцитов и митохондрий
Миелиновая оболочка- производное плазматической мембраны глиальных клеток (олигодендроглиальных). В расчете на сухую массу содержание липидов в миелине 70-80%, белков - 20-30%. Вода составляет 40% от массы свежей ткани. То есть по сравнению с другими плазматическими мембранами в миелине содержание липидов в
Таблица 18.3. Липиды миелина человека(в %)
| Холестерол | 27,7 | Цереброзиды | 22,7 |
| Фосфатидилэтаноламины | 15,6 | Фосфатидилхолины | 11,2 |
| Сфингомиелины | 7,9 | Фосфатидилсерины | 4,8 |
| Плазмалогены (фосфатидальэтаноламин) | 12,3 | Свободные жирные кислоты | в следовых количествах |
Липидов, специфичных для миелина, как таковых, не существует. Но миелин заметно отличается от других мембран по количественному содержанию липидов. Следует отметить высокое содержание цереброзида и пониженное количество ганглиозидов и фосфолипидов. Ганглиозиды почти полностью представлены Gm1 (структуру см. в гл.7). Для миелиновой мембраны типично значительное содержание цереброзидов, причем увеличение их количества во время развития эмбриона коррелирует с процессом миелинизации. И наоборот, содержание цереброзидов оказывается явно меньше, если в процессе миелинизации происходят патологические нарушения. К примеру, мутантные мыши с генетическими нарушениями миелиновой оболочки (визуально проявляются в виде "дрожания" или "подпрыгивания") содержат только 10% цереброзидов мозга нормальных мышей.
Миелин периферических нервов имеет подобное строение, но содержит меньше лецитина и больше сфингомиелинов. Период полураспада липидов колеблется от 5 недель (фосфатидилинозитол) до 2-4 месяцев (фосфатидилхолин, фосфатидилсерин) или до года (фосфатидилэтаноламин, холестерол, цереброзиды, сульфатиды, сфингомиелин).
Следствием высокого содержания липидов является малый процент белка. Время полураспада белков миелина составляет около 1 месяца. Белковый состав относительно прост:
· Водорастворимый щелочной белок составляет 1/3 всех белков миелина мозга. Обладает основными свойствами (ИЭТ 10.6), молекулярная масса 18 кД. Состоит из 170 аминокислот. Введение этого белка или его части (от 114 по 121 аминокислоты), животным вызывает энцефаломиелит, сопровождающийся демиелинизацией.
· Протеолипиды. Не растворяются в воде, но растворяются в смеси хлороформ-метанол. Эти белки составляют почти половину всех белков миелина мозга. Представляют смесь родственных белков с молекулярной массой 12500-30000 Да.
· Кислые белки. Эта группа белков называется белками Вольфграма, функция которых пока неизвестна.
В миелине обнаружены несколько видов ферментативной активности. Были выделены в чистом виде протеинкиназы, фосфодиэстеразы, фосфолипазы.
В нейроглии и нейронах обнаружены уникальные белки, которых нет в других тканях. К ним относятся:
· Белок S-100. Выделен из глии, назван так из-за его способности растворяться в насыщенных растворах сульфата аммония. Состоит из трех субъединиц с молекулярной массой 7000 Да, богат аспарагиновой и глутаминовой кислотами. Каждая молекула связывает два иона кальция.
· Белок 14-3-2. Выделен из серого вещества мозга. Он входит в нейроны и медленно перемещается из тела клетки по аксонам.
Название работы: Химический состав нервной ткани. Миелиновые мембраны: особенности состава и структуры
Предметная область: Биология и генетика
Описание: Данилевский впервые разделил белки мозговой ткани на растворимые в воде и солевых растворах белки и нерастворимые белки. которые разделили белки нервной ткани на 4 фракции: извлекаемые водой 45 раствором КСl 01 раствором NOH и нерастворимый остаток. В настоящее время сочетая методы экстракции буферными растворами хроматографии на колонках с ДЭАЭцеллюлозой и дискэлектрофореза в полиакриламидном геле удалось выделить из ткани мозга около 100 различных растворимых белковых фракций.
Дата добавления: 2015-02-20
Размер файла: 152.07 KB
Работу скачали: 13 чел.
Химический состав нервной ткани. Миелиновые мембраны: особенности состава и структуры.

На долю белков приходится примерно 40% от сухой массы головного мозга. Мозговая ткань является трудным объектом для изучения белкового состава вследствие большого содержания липидов и наличия белково-ли-пидных комплексов. А.Я. Данилевский впервые разделил белки мозговой ткани на растворимые в воде и солевых растворах белки и нерастворимые белки . Обширные исследования в этой области были проведены также А.В. Палладиным и сотр., которые разделили белки нервной ткани на 4 фракции: извлекаемые водой , 4,5% раствором КСl, 0,1% раствором NaOH и нерастворимый остаток. Установлено, что серое вещество богаче белками , растворимыми в воде , чем белое вещество , соответственно 30 и 19%. Белое вещество , напротив, содержит гораздо больше (22%) нерастворимого белкового остатка, чем серое вещество (5%). В дальнейшем было выделено 510 фракций растворимых белков мозга, различающихся по своей электрофоретической подвижности. В настоящее время, сочетая методы экстракции буферными растворами , хроматографии на колонках с ДЭАЭ-целлюлозой и диск-электрофореза в полиакриламидном геле , удалось выделить из ткани мозга около 100 различных растворимых белковых фракций. В нервной ткани содержатся как простые, так и сложные белки . Простые белки это альбумины (нейроальбумины), глобулины (нейроглобулины), катионные белки ( гистоны и др.) и опорные белки (нейросклеропротеины). Альбумины и глобулины по своим физико-химическим свойствам несколько отличаются от аналогичных белков сыворотки крови , поэтому их называют нейроальбуминами и нейроглобулинами. Количество нейрогло-булинов в головном мозге относительно велико в среднем 5% по отношению ко всем растворимым белкам . Нейроальбумины являются основным белковым компонентом фосфопротеинов нервной ткани , на их долю приходится основная масса растворимых белков (8990%). В свободном состоянии нейроальбумины встречаются редко. В частности, они легко соединяются с липидами , нуклеиновыми кислотами , углеводами и другими небелковыми компонентами. Белки , которые в процессе электрофоретического разделения при рН 10,512,0 движутся к катоду , получили название катионных. Главнейшими представителями этой группы белков в нервной ткани являются гистоны , которые делятся на пять основных фракций в зависимости от содержания в их полипептидных цепях остатков лизина , аргинина и глицина .
Нейросклеропротеины можно охарактеризовать как структурно-опорные белки . Основные представители этих белков нейроколлагены, нейроэлас-тины, нейростромины и др. Они составляют примерно 810% от общего количества простых белков нервной ткани и локализованы в основном в белом веществе головного мозга и в периферической нервной системе.
Сложные белки нервной ткани представлены нуклеопротеинами , липопротеинами, протеолипидами, фосфопротеинами , гликопротеинами и т.д. В мозговой ткани содержатся в значительном количестве еще более сложные надмолекулярные образования, такие, как липонуклеопротеины, липогликопротеины и, возможно, липогликонуклеопротеиновые комплексы.
Нуклеопротеины белки , которые принадлежат либо к дезоксирибонук-леопротеинам, либо к рибонуклеопротеинам . Часть этих белков из мозговой ткани извлекается водой , другая часть солевыми средами, а третья 0,1 М раствором щелочи .
Липопротеины составляют значительную часть водорастворимых белков мозговой ткани . Их липидный компонент это в основном фосфоглицериды и холестерин .
Протеолипиды это белково-липидные соединения, экстрагируемые органическими растворителями из ткани мозга. Отличаются от водорастворимых липопротеинов тем, что они нерастворимы в воде , но растворимы в смеси хлороформметанол. Белки , освобожденные от липидов , растворимы в воде , а также (благодаря высокому содержанию гидрофобных аминокислот ) в смеси хлороформметанол. Наибольшее количество про-теолипидов сосредоточено в миелине, в небольших количествах они входят в состав синаптических мембран и синаптических пузырьков.
Фосфопротеины в головном мозге содержатся в большем количестве, чем в других органах и тканях , около 2% от общего количества всех сложных белков мозга. Фосфопротеины обнаружены в мембранах различных морфологических структур нервной ткани .
Гликопротеины представляют собой чрезвычайно гетерогенную группу белков . По количеству белка и углеводов , входящих в состав гликопро-теинов, их можно разделить на две основные группы. Первая группа это гликопротеины , содержащие от 5 до 40% углеводов и их производных; белковая часть состоит преимущественно из альбуминов и глобулинов . В гликопротеинах , составляющих вторую группу, содержится 4085% углеводов , часто обнаруживается липидный компонент; по своему составу они могут быть отнесены к гликолипопротеинам.
В нервной ткани обнаружен ряд специфических белков , в частности белок S-100 и белок 14-3-2. Белок S-100, или белок Мура, называют также кислым белком , так как он содержит большое количество остатков глутаминовой и аспарагиновой кислот . Этот белок сосредоточен в основном в нейроглии (85-90%), в нейронах его не более 10-15% от общего количества белка в головном мозге. Установлено, что концентрация белка S-100 возрастает при обучении (тренировках) животных. Пока нет оснований считать, что белок S-100 непосредственно участвует в формировании и хранении памяти. Не исключено, что его участие в этих процессах опосредованно. Белок 14-3-2 также относится к кислым белкам . В отличие от белка S-100 он локализован в основном в нейронах ; в нейроглиальных клетках его содержание невелико. Пока неясна роль белка 14-3-2 в выполнении специфических функций нервной ткани .
Ферменты . В мозговой ткани содержится большое количество ферментов , катализирующих обмен углеводов , липидов и белков . До сих пор в кристаллическом виде из ЦНС млекопитающих выделены лишь некоторые ферменты , в частности ацетилхолинэстераза и креатинкиназа.
Значительное количество ферментов в мозговой ткани находится в нескольких молекулярных формах ( изоферменты ): ЛДГ, альдолаза , креатинкиназа, гексокиназа , малатдегидрогеназа , глутаматдегидрогеназа , холинэстераза, кислая фосфатаза , моноаминоксидаза и др.
Среди химических компонентов головного мозга особое место занимают липиды , высокое содержание и специфическая природа которых придают мозговой ткани характерные особенности. В группу липидов головного мозга входят фосфоглицериды , холестерин , сфингомиелины , цереброзиды , ганглиозиды и очень небольшое количество нейтрального жира . Многие липиды нервной ткани находятся в тесной взаимосвязи с белками , образуя сложные системы типа протеолипидов.
В сером веществе головного мозга фосфоглицериды составляют более 60% от всех липидов , а в белом веществе около 40%. Напротив, в белом веществе содержание холестерина , сфингомиелинов и особенно церебрози-дов больше, чем в сером веществ е.
В мозговой ткани имеются гликоген и глюкоза , но по сравнению с другими тканями ткань мозга бедна углеводами . Общее содержание глюкозы в головном мозге разных животных составляет в среднем 14 мкмоль на 1 г ткани , а гликогена 2,54,5 мкмоль на 1 г ткани . Интересно отметить, что общее содержание гликогена в мозге эмбрионов и новорожденных животных значительно выше, чем в мозге взрослых. Например, у новорожденных мышей в отличие от взрослых особей уровень гликогена в 3 раза выше. По мере роста и дифференцировки мозга концентрация гликогена быстро снижается и остается относительно постоянной у взрослого животного.
В мозговой ткани имеются также промежуточные продукты обмена углеводов : гексозо- и триозофосфаты, молочная, пировиноградная и другие кислоты .
В мозговой ткани на долю адениновых нуклеотидов приходится около 84% от всех свободных нуклеотидов . Большую часть оставшихся нуклеотидов составляют производные гуанина . В целом количество высокоэргических соединений в нервной ткани невелико. Содержание нуклеотидов и креатин-фосфата в головном мозге крыс составляет в среднем (в мкмоль на 1 г сырой массы): АТФ 2,302,90; АДФ 0,300,50; АМФ 0,030,05; ГТФ 0,200,30; ГДФ 0,150,20; УТФ 0,170,25; креатинфосфат 3 , 5 0 4,75. Распределение основных макроэргических соединений примерно одинаково во всех отделах мозга. Содержание циклических нуклеотидов ( цАМФ и цГМФ) в головном мозге значительно выше, чем во многих других тканях . Уровень цАМФ в мозге в среднем 12, а цГМФ до 0,2 нмоль на 1 г ткани . Для мозга характерна также высокая активность ферментов метаболизма циклических нуклеотидов . Большинство исследователей считают, что циклические нуклеотиды участвуют в синаптической передаче нервного импульса.
Ионы Na+, K+, Cu2+, Fe3+, Ca2+, Mg2+и Мn2+ распределены в головном мозге относительно равномерно в сером и белом веществе . Содержание фосфатов в белом веществе выше, чем в сером . Количественное соотношение неорганических анионов и катионов в мозговой ткани свидетельствует о дефиците анионов . Расчет показывает, что для покрытия дефицита анионов потребовалось бы в 2 раза больше белков , чем их имеется в мозговой ткани . Принято считать, что остающийся дефицит анионов покрывается за счет липидов . Вполне возможно, что участие липидов в ионном балансе одна из функций головного мозга.
Миелиновое вещество понятие морфологическое. По сути миелин это система, образованная многократно наслаивающимися мембранами клеток нейроглии вокруг нервных отростков (в периферических нервных стволах нейроглия представлена леммоцитами, или шванновскими клетками , а в белом веществе ЦНС астроцитами).
По химическому составу миелиновое вещество является сложным белково-липидным комплексом.
На долю липидов приходится до 80% плотного осадка; 90% всех липидов миелина представлено холестерином , фосфолипидами и церебро-зидами. Считают, что в липоидных слоях миелиновых оболочек молекулы различных липидов имеют строго определенное расположение.
На долю белков приходится примерно 40% от сухой массы головного мозга. Мозговая ткань является трудным объектом для изучения белкового состава вследствие большого содержания липидов и наличия белково-ли-пидных комплексов. А.Я. Данилевский впервые разделил белки мозговой ткани на растворимые в воде и солевых растворах белки и нерастворимые белки. Обширные исследования в этой области были проведены также А.В. Палладиным и сотр., которые разделили белки нервной ткани на 4 фракции: извлекаемые водой, 4,5% раствором КСl, 0,1% раствором NaOH и нерастворимый остаток. Установлено, что серое веществобогаче белками, растворимыми в воде, чем белое вещество, соответственно 30 и 19%. Белое вещество, напротив, содержит гораздо больше (22%) нерастворимого белкового остатка, чем серое вещество (5%). В дальнейшем было выделено 510 фракций растворимых белков мозга, различающихся по своей электрофоретической подвижности. В настоящее время, сочетая методы экстракции буферными растворами, хроматографии на колонках с ДЭАЭ-целлюлозой и диск-электрофореза в полиакриламидном геле, удалось выделить из ткани мозга около 100 различных растворимых белковых фракций. В нервной ткани содержатся как простые, так и сложные белки. Простые белки это альбумины (нейроальбумины), глобулины(нейроглобулины), катионные белки (гистоны и др.) и опорные белки (нейросклеропротеины). Альбумины и глобулины по своим физико-химическим свойствам несколько отличаются от аналогичных белков сыворотки крови, поэтому их называют нейроальбуминами и нейроглобулинами. Количество нейрогло-булинов в головном мозге относительно велико в среднем 5% по отношению ко всем растворимым белкам. Нейроальбумины являются основным белковым компонентомфосфопротеинов нервной ткани, на их долю приходится основная масса растворимых белков (8990%). В свободном состоянии нейроальбумины встречаются редко. В частности, они легко соединяются с липидами, нуклеиновыми кислотами, углеводами и другими небелковыми компонентами. Белки, которые в процессе электрофоретического разделения при рН 10,512,0 движутся к катоду, получили название катионных. Главнейшими представителями этой группы белков в нервной ткани являются гистоны, которые делятся на пять основных фракций в зависимости от содержания в их полипептидных цепях остатков лизина, аргинина и глицина.
Нейросклеропротеины можно охарактеризовать как структурно-опорные белки. Основные представители этих белков нейроколлагены, нейроэлас-тины, нейростромины и др. Они составляют примерно 810% от общего количества простых белковнервной ткани и локализованы в основном в белом веществе головного мозга и в периферической нервной системе.
Сложные белки нервной ткани представлены нуклеопротеинами, липопротеинами, протеолипидами, фосфопротеинами,гликопротеинами и т.д. В мозговой ткани содержатся в значительном количестве еще более сложные надмолекулярные образования, такие, как липонуклеопротеины, липогликопротеины и, возможно, липогликонуклеопротеиновые комплексы.
Нуклеопротеины белки, которые принадлежат либо к дезоксирибонук-леопротеинам, либо к рибонуклеопротеинам. Часть этих белковиз мозговой ткани извлекается водой, другая часть солевыми средами, а третья 0,1 М раствором щелочи.
Липопротеины составляют значительную часть водорастворимых белков мозговой ткани. Их липидный компонент это в основномфосфоглицериды и холестерин.
Протеолипиды это белково-липидные соединения, экстрагируемые органическими растворителями из ткани мозга. Отличаются от водорастворимых липопротеинов тем, что они нерастворимы в воде, но растворимы в смеси хлороформметанол. Белки, освобожденные от липидов, растворимы в воде, а также (благодаря высокому содержанию гидрофобных аминокислот) в смеси хлороформметанол. Наибольшее количество про-теолипидов сосредоточено в миелине, в небольших количествах они входят в состав синаптических мембран и синаптических пузырьков.
Фосфопротеины в головном мозге содержатся в большем количестве, чем в других органах и тканях, около 2% от общего количества всех сложных белков мозга. Фосфопротеины обнаружены в мембранах различных морфологических структур нервной ткани.
Гликопротеины представляют собой чрезвычайно гетерогенную группу белков. По количеству белка и углеводов, входящих в состав гликопро-теинов, их можно разделить на две основные группы. Первая группа это гликопротеины, содержащие от 5 до 40% углеводови их производных; белковая часть состоит преимущественно из альбуминов и глобулинов. В гликопротеинах, составляющих вторую группу, содержится 4085% углеводов, часто обнаруживается липидный компонент; по своему составу они могут быть отнесены к гликолипопротеинам.
В нервной ткани обнаружен ряд специфических белков, в частности белок S-100 и белок 14-3-2. Белок S-100, или белок Мура, называют также кислым белком, так как он содержит большое количество остатков глутаминовой и аспарагиновой кислот. Этот белоксосредоточен в основном в нейроглии (85-90%), в нейронах его не более 10-15% от общего количества белка в головном мозге. Установлено, что концентрация белка S-100 возрастает при обучении (тренировках) животных. Пока нет оснований считать, что белокS-100 непосредственно участвует в формировании и хранении памяти. Не исключено, что его участие в этих процессах опосредованно.Белок 14-3-2 также относится к кислым белкам. В отличие от белка S-100 он локализован в основном в нейронах; в нейроглиальныхклетках его содержание невелико. Пока неясна роль белка 14-3-2 в выполнении специфических функций нервной ткани.
Ферменты. В мозговой ткани содержится большое количество ферментов, катализирующих обмен углеводов, липидов и белков. До сихпор в кристаллическом виде из ЦНС млекопитающих выделены лишь некоторые ферменты, в частности ацетилхолинэстераза и креатинкиназа.
Значительное количество ферментов в мозговой ткани находится в нескольких молекулярных формах (изоферменты): ЛДГ, альдолаза, креатинкиназа, гексокиназа, малатдегидрогеназа, глутаматдегидрогеназа, холинэстераза, кислая фосфатаза, моноаминоксидаза и др.
Среди химических компонентов головного мозга особое место занимают липиды, высокое содержание и специфическая природа которых придают мозговой ткани характерные особенности. В группу липидов головного мозга входят фосфоглицериды, холестерин,сфингомиелины, цереброзиды, ганглиозиды и очень небольшое количество нейтрального жира. Многие липиды нервной ткани находятся в тесной взаимосвязи с белками, образуя сложные системы типа протеолипидов.
В сером веществе головного мозга фосфоглицериды составляют более 60% от всех липидов, а в белом веществе около 40%. Напротив, в белом веществе содержание холестерина, сфингомиелинов и особенно церебрози-дов больше, чем в сером веществе.
В мозговой ткани имеются гликоген и глюкоза, но по сравнению с другими тканями ткань мозга бедна углеводами. Общее содержаниеглюкозы в головном мозге разных животных составляет в среднем 14 мкмоль на 1 г ткани, а гликогена 2,54,5 мкмоль на 1 г ткани. Интересно отметить, что общее содержание гликогена в мозге эмбрионов и новорожденных животных значительно выше, чем в мозге взрослых. Например, у новорожденных мышей в отличие от взрослых особей уровень гликогена в 3 раза выше. По мере роста и дифференцировки мозга концентрация гликогена быстро снижается и остается относительно постоянной у взрослого животного.
В мозговой ткани имеются также промежуточные продукты обмена углеводов: гексозо- и триозофосфаты, молочная, пировиноградная и другие кислоты .
В мозговой ткани на долю адениновых нуклеотидов приходится около 84% от всех свободных нуклеотидов. Большую часть оставшихсянуклеотидов составляют производные гуанина. В целом количество высокоэргических соединений в нервной ткани невелико. Содержание нуклеотидов и креатин-фосфата в головном мозге крыс составляет в среднем (в мкмоль на 1 г сырой массы): АТФ 2,302,90; АДФ 0,300,50; АМФ 0,030,05; ГТФ 0,200,30; ГДФ 0,150,20; УТФ 0,170,25; креатинфосфат 3 , 5 0 4,75. Распределение основных макроэргических соединений примерно одинаково во всех отделах мозга. Содержание циклических нуклеотидов (цАМФ и цГМФ) в головном мозге значительно выше, чем во многих других тканях. Уровень цАМФв мозге в среднем 12, а цГМФ до 0,2 нмоль на 1 г ткани. Для мозга характерна также высокая активность ферментов метаболизмациклических нуклеотидов. Большинство исследователей считают, что циклические нуклеотиды участвуют в синаптической передаче нервного импульса.
Ионы Na+, K+, Cu2+, Fe3+, Ca2+, Mg2+и Мn2+ распределены в головном мозге относительно равномерно в сером и белом веществе. Содержание фосфатов в белом веществе выше, чем в сером. Количественное соотношение неорганических анионов и катионов в мозговой ткани свидетельствует о дефиците анионов. Расчет показывает, что для покрытия дефицита анионов потребовалось бы в 2 раза больше белков, чем их имеется в мозговой ткани. Принято считать, что остающийся дефицит анионов покрывается за счет липидов. Вполне возможно, что участие липидов в ионном балансе одна из функций головного мозга.
Миелиновое вещество понятие морфологическое. По сути миелин это система, образованная многократно наслаивающимисямембранами клеток нейроглии вокруг нервных отростков (в периферических нервных стволах нейроглия представлена леммоцитами, или шванновскими клетками, а в белом веществе ЦНС астроцитами).
По химическому составу миелиновое вещество является сложным белково-липидным комплексом.
На долю липидов приходится до 80% плотного осадка; 90% всех липидов миелина представлено холестерином, фосфолипидами и церебро-зидами. Считают, что в липоидных слоях миелиновых оболочек молекулы различных липидов имеют строго определенное расположение.
Белки сост-ют 50% на сухой вес сер.в-ва и 30% белого. Более интенсивное включение АК в белки идет в КБП. Классификация белков мозга: 1.Опорные(нейросклеропротеиды,нейроглобулины); 2.Мембранные. Ф-ии: а)участие в форм-ии и контроле за работой ион.каналов, б)рецепторные, в)участие в создании потенциалов покоя и действия; 3.Специфические(обучение-белок Мура; память,диф-ка нейронов-ф-ры роста нервов). АК: 1.уч-ют в синтезе специфич.белков мозга 2.вып-ие медиаторной ф-ии 3.явл-ся медиаторами: глутаминовая, аспарагиновая, глицин 4.из АК обр-ся биоген.амины-медиаторы:АХ,катехоламины,ГАМК,серотонин 5.из АК обр-ся нейропептиды: глутатион; специфич.:эндорфины, энкефалины,гипофизарные г-ны 6.вып-ют дезинтоксикац.ф-ю: глутаминовая к-та обезвреж-ет избыток NH3. 75%пула АК-глутам.к-та; она влияет на настроение, внимание. Ф-ии глутаминовой к-ты: 1.-CO2→ГАМК(в сер.в-ве) 2.-NH3→NH3(при необ-ти-возб-ль ф-ии нейронов) 3.+NH3→связывание избытка NH3 4.обр-ие α-кетоглутарата→ЦТК→энергетич.ф-я.
Липиды мозга. Хар-но: 1.высокое сод-ие(до 50% от сух.массы) 2.разнообразие стр-ры 3.Постоянство состава 4.Разнооб-е ЖК состава,высокое содер-ие ПНЖК(арахидоновая) 5.специфичность ЖК состава разн.отделов мозга, разн.типов клеток, субклет.структур; в миелине высокое сод-ие олеиновой. Изм-ие ЖК состава ведет к нарушению функц.деят-ти мозга. Ф-ии: 1.структурная-участие в построении мембран 2.ф-ия диэлектриков-электрич.изоляция 3.защитная: ганглиозиды активные антиоксиданты,ингибиторы ПОЛ 4.регул-ая: фосфоинозиты имеют отн-ие к проведению нерв.сигнала. Холестерин.У взрослого 25г, у новорож-2г; в первый год ↑в 3р. Мозг способен синтезировать ХС из ацетил-КоА.
Миелиновая об-ка обладает изолирующими св-ми. 70-80% состава миелина-это липиды; ХС-0,775 мкмоль/мг,фосфотидилхолин-0,116.Белки миелина гидрофобны.
Энергетика мозга. Энергия необ-ма для: 1.поддер-ия ион.потенциалов ионов по обе стороны нейрон.мем-ны 2.хранения и переработки поступающей инф-ии 3.поддер-ия архитектоники мозга. Фонд макроэргов мозга: 1.АТФ 2.креатинфосфат, стабилизирующий уровень АТФ; его в 2р больше чем АТФ. Энерг.ист-ом для мозга явл-ся аэробный путь ок-ия глюкозы.След-но энергетика мозга зависит от поступления О2 и глюкозы. Потреб-ть в О2-мозг погл-ет 20% поступающ.О2, больше всего потр-ет КБП. Потреб-ть в глюкозе-самый большой потребитель энергии из всех органов.85-90% глюкозы полностью ок-ся аэробно(до Н2О и СО2), 5-7% исп-ся на синтез гликогена, гликолипидов,нейротрансмиттеров. Депо глюкозы-750мг(10мин жизни). Ист-ки Г. для мозга: гликоген мозга и кровь(пища,гликоген печени,глюконеогенез). Недост.поступление Г. в мозг сопр-ся голов.болью,↓внимания,наруш-ем умств.трудоспос-ти, сост-ем подавленности и т.д. При длит.голодании ист-ом энергии явл-ся кетоновые тела.
110. Медиаторы: ацетилхолин, биогенные амины, аминокислоты, пептиды; образование и инактивация. Связь нарушений обмена медиаторов с заболеваниями цнс.
Нейромедиатор(нейротрансмиттер)-в-во,кот.синтез-ся в нейроне, сод-ся в пресин.окончаниях,высвобождается в синап.щель в ответ на нерв.импульс,и действует на спец.участки постсинап.клетки,вызывая изменения мембр.потенциала и метаболизма клетки.
1группа-Амины(АХ,А,НА,дофамин,серотонин);2гр-АК;3гр-пуриновые нуклеотиды;4гр-нейропептиды(самая болш.группа). Ацетилхолин. Холинэргич.с-мы уч-ют в таких ф-ях как память, рег-ция движения,двиг.стереотип, уровень бодрствования. Синт-ся из Ацетил-КоА(актив.форма ацетата) и холина при помощи ацетилхолинтрансферазы,с участием АТФ,ионов Mg,Ca,K,Na. Для обр-ия холина необх-мы метионин и фолиевая к-та. Инактивация АХ: 1.при пом. ацетилхолинэстеразы до холина и ацетата. 2.энергозависимый транспорт в нейрон и повтор.использование(реутилизация). Нарушение в с-ме инактивации-миастении при превышает АХ.Лечение-антихолинэстеразные препараты. Норадреналин(НА) синт-ся в нерв.концевых пластинках из тирозина при уч-ии НАДФН2,В6,аскорб.к-ты и ионов Са. Рецепция.Сущ-ет 2 типа рецепторов α и β.Передача импульса с α-адренорецепторов связана с цГМФ,а с β-рецепторов с цАМФ. Агонистом НА явл-ся адреналин. Инактивация: 1.под действием МАО с участием НАД или ФАД; 2.путем метилирования с участием S-аденозилметионина и катехол-о-метилтрансферазы(превращ-е в адреналин-типичный гормон) и последующим ок-ем в печени и выделением с мочой. НА нейроны причастны к поддержанию бодрствования,с-ме поощрения(центр удовольствия), к сновидениям и рег-ии настроения. В стволе мозга(преим.голубое пятно)-ф-ии-концентрация внимания;активация НА имеет место при панических атаках. Дофамин(ДФА) Нейроны,сод-ие ДФА-в сред.мозге черн.субстанции.Многие нейроны посылают свои аксоны в перед.мозг,где уч-ют в поддержании эмоц.реакций. Другие ДФА волокна окан-ся в области полос.тела,где играет решающую роль в рег-ии слож.движений. Дегенерация ДФА волокон приводит к регидности мышц,тремору(бол.Паркенсона). Для обр-ия: 1.АК:тирозин,метионин; 2.Вит:В6,С,ФК,РР. Глутаминовая к-та(Глу к-та): 1.-СО2→ГАМК(торм.м-р); 2.+NH3→глутамин связ-е NH3; 3.-NH3→α-кетоглутаровая к-та(энерг.материал). Ф-ии. ГАМК. Ф-ии: 1.рег-ия мотор.деят-ти; 2.поддержание судорожной акт-ти; 3.форм-е поведенческих реакций(агрессия,защитное поведение); 4.контроль высвобождения ряда гипофизарных гормонов; 5.осущ-ие высших интегральных ф-й(усл.р-сы,память,обучение). Серотонин производное АК триптофана; из него обр-ся метионин. В обр-ии серотонина уч-ют:триптофан,В6,В2,РР,С. Медиаторы-пептиды: энкефалины,эндорфины, холецистокинин, вещ-во Р,брадикинин, ангиотензин||, тиролиберин, соматостатин и др. Энкефалины и эндорфины имеются в спин.мозге-в сенс.нейронах, воспринимающих чув-во боли,и в нейронах лимб.с-мы,регул-их эмоции.Эти пептиды обр-ся путем частичного гидролиза белков-предшественников.
Читайте также:
