
Какие медиаторы выделяются в симпатической нервной системе
а) Ганглионарная передача нервных импульсов. Преганглионарные нейроны симпатической и парасимпатической систем — холинергические: при образовании аксодендритических синапсов с ганглионарными клетками из этих нейронов высвобождается ацетилхолин (АХ). Рецепторы на поверхности ганглионарных клеток называют никотиновыми, так как их возбуждение может происходить при местном действии никотина.
б) Передача нервных импульсов в нейроэффекторном синапсе. Постганглионарные нервные волокна симпатической и парасимпатической систем образуют нейроэффекторные соединения (синапсы)с эффекторными тканями (тканями-мишенями). За высвобождение нейромедиаторов отвечают многочисленные пресинаптические утолщения, расположенные по ходу нервных волокон.
Главный нейромедиатор в симпатических нейроэффекторных соединениях — норадреналин (норэпинефрин), который высвобождается из гранулярных везикул. Постганглионарные симпатические волокна — преимущественно адренергические; исключение составляют холинергические волокна, отвечающие за симпатическую иннервацию расположенных по всему телу эккриновых потовых желез. Главный нейромедиатор в парасимпатических нейроэффекторных соединениях — ацетилхолин (АХ). Постганглионарные парасимпатические волокна преимущественно холинергические.
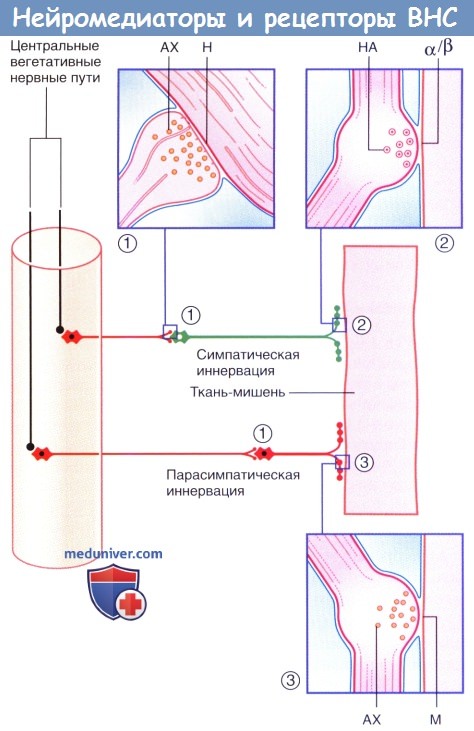
Нейромедиаторы и рецепторы вегетативной нервной системы.
(1) Аксодендритические синапсы с никотиновыми рецепторами.
(2) Нейроэффекторные синапсы с адренергическими рецепторами.
(3) Нейроэффекторные синапсы с мускариновыми рецепторами. Ганглионарные нейроны и постганглионарные волокна выделены красным цветом.
АХ — ацетилхолин; М — мускариновые рецепторы; Н — никотиновые рецепторы; НА — норадреналин.
в) Рецепторы в нейрозффекторном синапсе. Физиологические эффекты вегетативной нервной системы (ВНС) зависят от вида рецепторов на постсинаптической мембране (плазматической мембране эффекторных клеток). На высвобождение нейромедиаторов также влияют рецепторы пресинаптической мембраны (аксолеммы).
- Синаптические рецепторы симпатической системы (адренорецепторы). Для норадреналина существуют два вида а-адренорецепторов и два вида β-адренорецепторов.
1. Постсинаптические α1-адренорецеторы, активация которых вызывает сокращение гладких мышц мелких периферических артерий и крупных артериол, дилататора зрачка, а также мышц семявыносящего протока, сфинктеров ЖКТ и шейки мочевого пузыря.
2. Пресинаптические α2-адренорецеторы локализуются как на симпатических, так и на парасимпатических нервных окончаниях и ингибируют в них высвобождение нейромедиаторов. Пресинаптические α2-адренорецеторы симпатической системы называют ауторецепторами.
3. Постсинаптические адренорецепторы, возбуждение которых приводит к увеличению пейсмекерной активности клеток сердца, а также к повышению силы сердечных (желудочковых) сокращений. При резком падении артериального давления за счет активации симпатических β1-адренорецепторов юкстагломерулярных клеток почек происходит секреция ренина, который, в свою очередь, вызывает секрецию мощного вазоконстриктора ангиотензина II.
4. β2-Адренорецепторы, которые реагируют как на норадреналин, так и на адреналин (эпинефрин).
Активация постсинаптических β2-адренорецепторов вызывает расслабление гладких мышц, наиболее ярко выраженное в трахеобронхиальном дереве и мышцах глаза, участвующих в аккомодации. Часть постсинаптических β2-адренорецепторов располагается на поверхности гепатоцитов. При возрастании энергетических затрат организма их активация вызывает распад гликогена, необходимый для поддержания уровня глюкозы в крови.
Пресинаптические β2-адренорецепторы адренергических нервных окончаний отвечают за высвобождение норадреналина.
В симпатических нервных окончаниях большая часть высвобождаемого норадреналина подвергается обратному захвату моноаминным ферментным насосом. После обратного захвата часть норадреналина разрушается митохондриальным ферментом — моноаминоксидазой (МАО). Влияние лекарственных средств на симпатическую систему подробно рассмотрено в блоке клинической информации ниже.
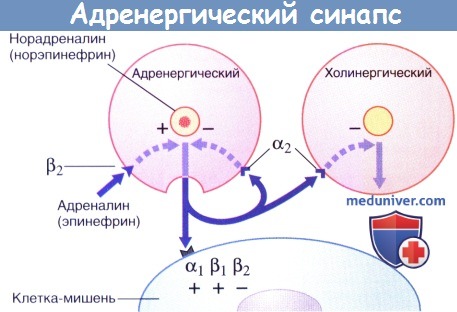
Передача возбуждения в адренергическом нейроэффекторном синапсе.
Секреция норадреналина стимулируется адреналином и ингибируется путем активации пресинаптических α2-рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих парасимпатических синапсов, где также ингибируют секрецию).
- Синаптические рецепторы парасимпатической системы. Парасимпатические синаптические рецепторы называют мускариновыми, так как их активацию, помимо АХ, может вызывать другое химическое вещество — мускарин. Стимуляция парасимпатической системы приводит к следующим М-холинергическим эффектам.
• Замедление (в ответ на стимуляцию блуждающего нерва) частоты сердечных сокращений, а также уменьшение силы желудочковых сокращений.
• Сокращение гладких мышц, которое обусловливает такие процессы, как перистальтика кишечника, опустошение мочевого пузыря, а также рефлекс аккомодации при взгляде на близкое расстояние.
• Секреция желез.
Помимо представленных выше эффектов, пресинаптические мускариновые рецепторы симпатических нервных окончаний также отвечают за ингибирование высвобождения норадреналина.
Влияние лекарственных средств на парасимпатическую систему подробно рассмотрено в Блоке клинической информации. Лекарственные средства, обладающие мускариноподобным действием, называют холинергическими (холиномиметическими). Лекарственные средства, блокирующие доступ АХ к постсинаптической мембране, называют антихолинергическими (холинолитическими).
Главный фактор, который необходимо учитывать при назначении препаратов, активирующих или подавляющих деятельность симпатической и парасимпатической систем,— наличие рецепторов к этим лекарственным веществам α-, β- и мускариновых) в ЦНС. Так, в частности, применение данных лекарственных средств в психиатрии обусловлено их действием на центральные, а не на периферические рецепторы.
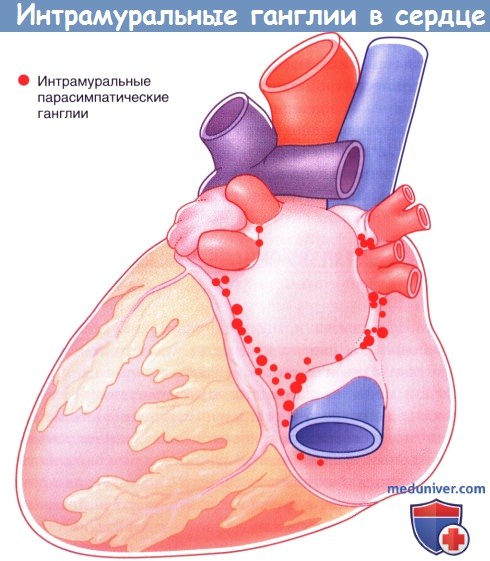
Расположение парасимпатических интрамуральных ганглиев в сердце. 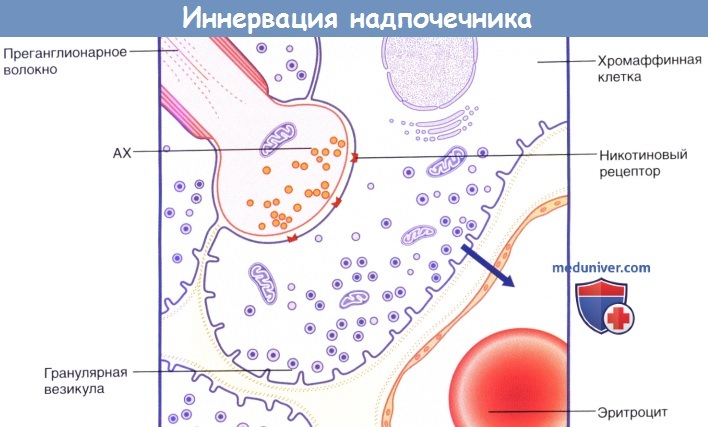
Синаптическое соединение преганглионарного волокна грудного внутренностного нерва и хромаффинной клетки мозгового вещества надпочечников.
Активация никотиновых рецепторов ацетилхолином (АХ).
8% клеток содержат крупные гранулярные везикулы (представлены на рисунке), высвобождающие адреналин; его секрекция в капиллярное русло обозначена стрелкой.
20% клеток содержат маленькие гранулярные везикулы, высвобождающие норадреналин. 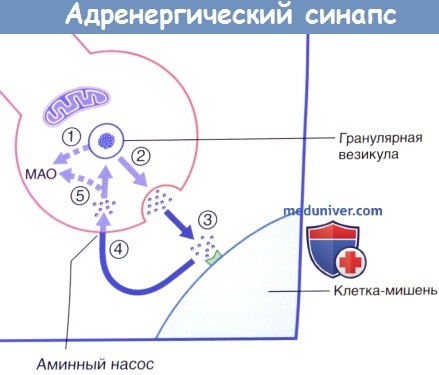
Высвобождение и обратный захват нейромедиатора в адренергическом нервном окончании.
МАО — моноаминоксидаза. 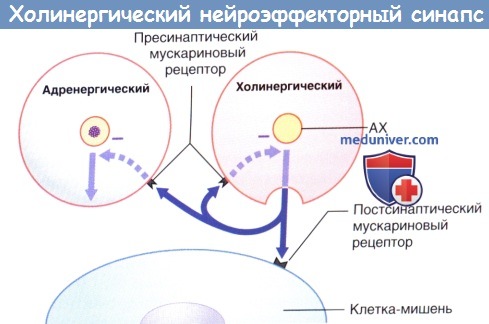
Передача возбуждения в холинергическом нейроэффекторном синапсе.
Избыточная секреция ацетилхолина (АХ) ингибируется путем активации пресинаптических мускариновых рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих симпатических синапсов, где также ингибируют секрецию).
г) Другие типы нейронов. И в симпатическом, и в парасимпатическом отделах ВНС часть нейронов представлена неадренергическими, нехолинергическими нейронами (NANC-нейронами). Находящиеся в симпатических ганглиях небольшие вставочные нейроны отвечают за высвобождение дофамина — предшественника норадреналина. Часть дофамина выделяется в капиллярное русло, другая часть связывается с дофаминовыми рецепторами на основных (адренергических) нейронах, оказывая на них слабое тормозное действие.
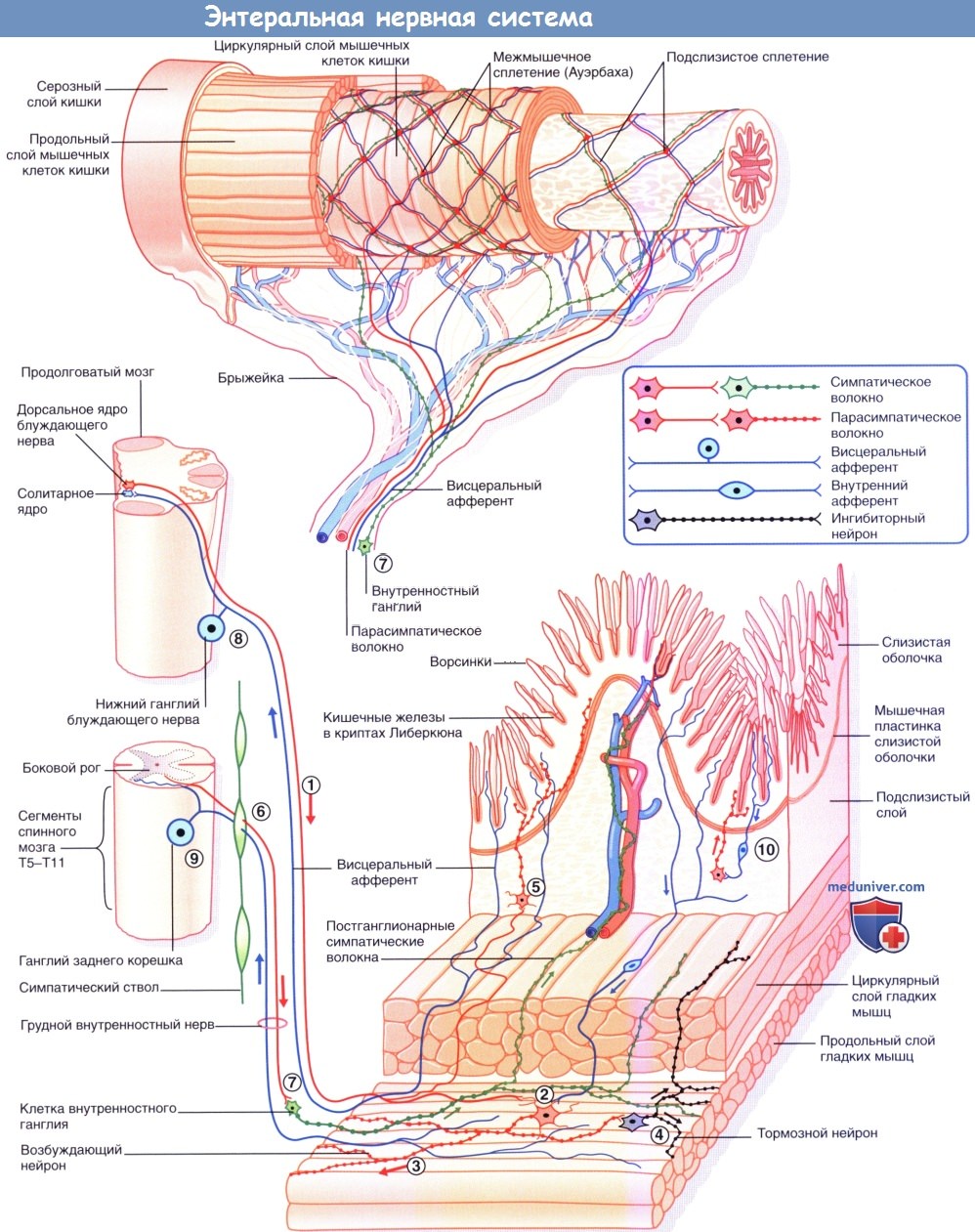
Самое большое количество NANC-нейронов находится в области ганглионарных клеток стенки ЖКТ и в тазовых ганглиях. Эти нейроны содержат более 50 разнообразных белковых субстанций, находящихся в несвязанном состоянии или в различных комбинациях. Функция большей части этих веществ — регуляторная: действуя на пресинаптическую или постсинаптическую мембрану, они влияют на продолжительность работы основных нейромедиаторов. Другие белковые субстанции (вещества, выделяющиеся совместно с ацетилхолином (АХ)) представляют собой комедиаторы (сопутствующие медиаторы).
Редактор: Искандер Милевски. Дата публикации: 14.11.2018
Медиаторы вегетативной нервной системы - это трансмиттеры, которые используются в синапсах вегетативной нервной системы для передачи возбуждения или наведения торможения на клетки-мишени.
Следует принять во внимание, что деление трансмиттеров на медиаторы и модуляторы, на возбуждающие и тормозные медиаторы, является условным и очень относительным. Дело в том, что один и тот же биолиганд может выполнять в организме разные функции: и медиатора, и модулятора, и возбуждающего медиатора, и тормозного, и гормона, и гистогормона.
В симпатическом отделе вегетативной нервной системы используется норадреналин, а в парасимпатическом - ацетилхолин.
| № | Название | Локализация синапсов | Рецепторы | Эффекты |
| 1 | Ацетилхолин (АХ), низкомолекулярное вещество с зарядом +1 | а) вегетативные ганглии, б) моторные пластинки скелетных мышц | н-холинорецепторы (n-холинорецепторы, никотиновые) - молекулярные рецепторы ионотропного типа, для них ацетилхолин является медиатором | Возбуждение постсинаптической мембраны. |
| Постганглионарные окончания парасимпатической вегетативной нервной системы. | м-холинорецепторы (m-холинорецепторы, мускариновые) - молекулярные рецепторы метаботропного типа, для них ацетилхолин является модулятором | а) в сосудах и сердце - тормозящий эффект: расширение сосудов, замедление и ослабление сокращений сердца; б) в бронхах, ЖКТ, радужке - стимуляция эффекторных клеток: сужение бронхов, усиление перистальтики и секреции ЖКТ, сужение зрачка. | ||
| 2 | Норадреналин (НА), из группы катехоламинов, производное аминокислоты тирозина | Постганглионарные окончания симпатической вегетативной нервной системы. | α1- , α2-, β1-, β2-адренорецепторы (все метаботропного типа) | Возбуждение α1- и β1- рецепторов оказывает возбуждающее действие, а α2- и β2-рецепторв - тормозящее. Отсюда - набор симпатических реакций: 1) усиление сокращений сердца, 2) сужение сосудов мышц, 3) расширение бронхов, 4) ослабленние перистальтики кишечника, 5) расширение зрачков. |
| Обычно в одном органе содержится несколько типов адренорецепторов. При этом α-рецепторы преобладают в сосудах кожи и сфинктерах ЖКТ, β1-рецепторы - в сердце, β2-рецепторы - в бронхах, кишечнике, матке, сосудах скелетных мышц. | ||||
Мушкамбаров Н.Н., Кузнецов С.Л. Молекулярная биология. Учебное пособие для студентов медицинских вузов. М.: ООО "Медицинское информационное агентство", 2003. 544 с.
Симпатическая часть вегетативной нервной системы состоит из центрального и периферического отделов. Центральный отдел образуют клетки боковых рогов спинного мозга на уровне всех его грудных и верхних трех поясничных сегментов. Периферический отдел представлен нервными волокнами и симпатическими нервными узлами (ганглиями). Нервные узлы подразделяются на две группы:
околопозвоночные (паравертебральные), расположенные двумя цепочками по бокам от позвоночника и образующие правый и левый симпатические стволы, и предпозвоночные (превертебральные) — узлы периферических нервных сплетений, лежащие в грудной и брюшной полостях
Основным медиатором симпатических синапсов является норадреналин и такие синапсы называютадренергическими.Рецепторы, связывающие адренергический медиатор получили названиеадренорецепторов.Различают два типа адренорецепторов — альфаи бета,каждый из которых делят на два подтипа — 1 и 2. Небольшая часть симпатических синапсов использует медиатор ацетилхолин и такие синапсы называют холинергическими,а рецепторы — холинорецепторами.Холинергические синапсы симпатической нервной системы обнаружены в потовых железах. В адренергических синапсах кроме норадреналина в существенно меньших количествах содержатся адреналин и дофамин, также относящиеся к катехоламинам, поэтому медиаторное вещество в виде смеси трех соединений раньше называли симпатином.
Норадреналин (НА), из группы катехоламинов, производное аминокислоты тирозина Постганглионарные окончаниясимпатическойвегетативной нервной системы. α1- , α2-, β1-, β2-адренорецепторы (все метаботропного типа) Возбуждение α1- и β1- рецепторов оказывает возбуждающее действие, а α2- и β2-рецепторв - тормозящее.
Отсюда - набор симпатических реакций:
1) усиление сокращений сердца,
2) сужение сосудов мышц,
3) расширение бронхов,
4) ослабленние перистальтики кишечника,
5) расширение зрачков.
Обычно в одном органе содержится несколько типов адренорецепторов. При этом α-рецепторы преобладают в сосудах кожи и сфинктерах ЖКТ, β1-рецепторы - в сердце, β2-рецепторы - в бронхах, кишечнике, матке, сосудах скелетных мышц.
6. Характеристика парасимпатического отдела вегетативной нервной системы его медиаторы
Центрами парасимпатического отдела автономной нервной системы являются ядра, находящиеся в среднем мозге (IIIпара черепно-мозговых нервов), продолговатом мозге (VII, IXи Х пары черепно-мозговых нервов) и крестцовом отделе спинного мозга (ядра тазовых внутренних нервов). От среднего мозга отходят Преганглионарные волокна парасимпатических нервов, которые входят в состав глазодвигательного нерва. Из продолговатого мозга выходят Преганглионарные волокна, идущие в составе лицевого, языкоглоточного и блуждающих нервов. От крестцового отдела спинного мозга отходят Преганглионарные парасимпатические волокна, которые входят в состав тазового нерва.
При возбуждении парасимпатических нервов тормозится работа сердца отрицательные хроно-, ино-, дромо- и батмотропное действия),повышается тонус гладкой мускулатуры бронхов, в результате чего уменьшается их просвет, сужается зрачок, стимулируются процессы пищеварения (моторика и секреция), обеспечивая тем самым восстановление уровня питательных веществ в организме, происходит опорожнение желчного пузыря, мочевого пузыря, прямой кишки. Действие парасимпатической нервной системы направлено на восстановление и поддержание постоянства состава внутренней среды организма, нарушенного в результате возбуждения симпатической нервной системы. Парасимпатическая нервная система выполняет в организметрофотропную функцию.
К этому отделу относятся интрамуральные системы всех полых висцеральных органов, обладающих собственной автоматической двигательной активностью: сердце, бронхи, мочевой пузырь, пищеварительный тракт, матка, желчный пузырь и желчные пути.
Внутриорганный отдел имеет все звенья рефлекторной дуги: афферентный, вставочный и эфферентный нейроны, которые полностью находятся в органе и нервных сплетениях внутренних органов (например, ауэрбаховском и мейснеровском). Преганглионарные волокна выделяютацетилхолининорадреналин,постганглионарные —АТФиаденозин, ацетилхолин, норадреналин, серотонип, дофамин, адреналин, гистамини т.д. Главная роль принадлежит АТФ и аденозину.
Ацетилхолин (АХ), низкомолекулярное вещество с зарядом +1
Дата добавления: 2019-11-25 ; просмотров: 461 ;
МЕДИАТОРЫ нервной системы (лат. mediator посредник; син.: нейротрансмиттеры, синаптические передатчики) — химические передатчики нервного импульса с нервного окончания на клетки периферических органов или на нервные клетки. Чаще всего в роли Медтаторов выступают низкомолекулярные (150—300 дальтон) вещества, выполняющие в организме человека и животных и другие функции. К Медиаторам относят ацетилхолин (см.), различные катехоламины (см.), в частности норадреналин (см.), некоторые аминокислоты (см.), пептиды (см.) и другие биологически активные вещества. Исследования Медиаторов дали важные практические результаты для клиники. Оказалось, что при ряде заболеваний нервной системы, некоторых видах отравлений нарушаются образование М., механизм их действия и распада. Знание особенностей хим. превращений М. в норме и при патологии позволило рекомендовать и ввести новые методы медикаментозного лечения.
Гипотеза о существовании веществ—посредников нервных влияний возникла в начале 20 в. Первоначально она основывалась на опыте фармакологии (имитация симпатических нервных влияний одними экзогенными веществами, а парасимпатических — другими) и относилась только к периферическим нейроэффекторным соединениям. Эллиотт (Th. R. Elliott, 1904) назвал адреналин веществом, к-рое могло бы опосредовать действие симпатических нервов на эффекторные органы. Экспериментально существование ОД. вегетативных нервов доказал в 1921 г. австр. фармаколог О. Леви, установивший, что перфузат сердца после раздражения блуждающего нерва способен оказывать вагоподобное действие. На этом основании М. первоначально называли гуморальными факторами нервного возбуждения. Впоследствии от этого названия отказались, т. к. стало ясно, что попадание М. в кровь — побочное и необязательное следствие процесса синаптической передачи.
Тип медиаторов вегетативных нейроэффекторных соединений не во всех случаях определяется их принадлежностью к тому или иному отделу в. н. с. В связи с этим возникло предположение, что он специфичен для клеточных, а не для анатомических единиц нервной системы. Г. Дейл (1933) предложил называть нервные волокна, выделяющие ацетилхолин, холинергическими, а волокна, выделяющие адреналин (фактически норадреналин),— адренергическими.
Значительный вклад в понимание механизмов действия М. внесли также советские ученые А. Г. Гинецинский, X. С. Коштоянц, М. Я. Михельсон, В. Н. Черниговский, С. В. Аничков и др. Уже в 30-х гг. в СССР развернулась работа по применению М. для лечения нервных болезней.
Участие М. в проведении возбуждения представляется следующим образом. Местом приложения М. является синапс (см.). Пресинаптическим звеном его может быть нейрон (см. Нервная клетка), либо рецепторная клетка (напр., палочки и колбочки сетчатки, волосковые клетки органов слуха и равновесия). Пресинаптической клетке, по-видимому, присуща медиаторная специфичность, т. е. способность синтезировать, запасать, секретировать и реутилизировать строго определенный М. Цитоплазматическими органеллами, в которых запасаются и посредством которых выделяются из клетки М., служат, по везикулярной гипотезе секреции М., особые окруженные мембраной пузырьки. Специализированный для секреции участок пресинаптической клетки (в нейроне — терминальные части аксона, а иногда и дендриты) имеет особую наружную так наз. секреторную мембрану, для к-рой характерно наличие потенциалзависимых кальциевых каналов. Секрецию вызывает входящий ток ионов кальция, возникающий при деполяризации пресинаптической клетки (т. е. при ее возбуждении). Тонкие механизмы действия ионов кальция на синаптические пузырьки еще не изучены; по-видимому, секреция протекает по типу экзоцитоза: мембрана пузырька соединяется с наружной клеточной мембраной так, что образуется отверстие, через к-рое содержимое пузырька выходит в межклеточную среду.
Выйдя в синаптическую щель, М. диффундирует к постсинаптической клетке и взаимодействует с ее специфическими рецепторами, вследствие чего происходит то или иное изменение в состоянии клетки. В основе этого регуляторного эффекта чаще всего лежит изменение ионной проводимости постсинаптической мембраны.
Для каждого М. стараются найти наиболее характерные признаки, позволяющие обнаруживать клеточные системы с данным М. С помощью различных гистохим, методов, в частности формальдегидной конденсации, удалось детально картировать системы моноаминергических нейронов мозга. Однако прямая гистохим. локализация пока возможна лишь для немногих М. Более перспективным и универсальным методом считается иммуногистохим, выявление фермента, участвующего в синтезе данного М. или другого специфического белка, связанного с определенным М. С этой целью используют также способность нейронов реутилизировать собственный М. Для этого в окружающую нейрон среду вводят М. или его метаболический предшественник с какой-либо (напр;, радиоактивной) меткой и изучают последующее распределение метки. О принадлежности к тому или иному известному типу М. помогает судить также изучение морфологии секреторных пузырьков пресинаптической клетки с помощью электронного микроскопа.
Распределение нейронов, обладающих одним и тем же М., и их функции сходны у систематически близких организмов. Это сходство прослеживается в пределах только одного зоол, типа и не наблюдается при сравнении разных типов (напр., позвоночных, членистоногих и моллюсков). Однако организмы, относящиеся к разным зоол, типам, имеют одни и те же медиаторные вещества, т. е. сходный клеточный состав нервных систем. Это указывает на глубокую древность медиаторных различий между нейронами и на консерватизм специфического секреторного химизма нервных клеток.
Значительная часть известных М. относится к группе биогенных аминов (см.), к-рую составляют декарбоксилированные производные ароматических аминокислот (так наз. арилэтиламины). В эту группу входят катехоламиновые М. Древнейшим (с эволюционной точки зрения) из них является, по-видимому, дофамин, представленный в специальной категории нейронов у большинства организмов, обладающих нервной системой. Медиаторная функция дофамина доказана на гигантском интернейроне педального ганглия некоторых водных улиток. У млекопитающих системы дофаминергических нейронов находятся преимущественно в среднем мозге — нигронеостриарная система (см. Лимбическая система). Кроме того, нейроны этого типа имеются в гипоталамической области, в сетчатке. Предполагают, что дофамин выступает в качестве М. интернейронов симпатических ганглиев (нейронный вариант хромаффинных клеток). Функция норадреналина в качестве М. наиболее изучена в нейроэффекторных окончаниях симпатических нервов. Группы нор адренергических нейронов имеются также в среднем мозге, мозговом варолиевом мосту, продолговатом и промежуточном мозге. Адреналин, являющийся метилированным производным норадреналина, служит М. симпатических нейронов у бесхвостых амфибий. В продолговатом мозге млекопитающих найдены небольшие группы нейронов, синтезирующих адреналин, однако вопрос о медиаторной функции адреналина у них изучен еще недостаточно.
Широко распространенный биогенный амин серотонин (см.) является производным триптофана. Медиаторная функция серотонина была впервые показана на моллюсках. Серотонинергические нейроны некоторых ядер ствола головного мозга иннервируют обширные области ц. н. с. млекопитающих, включая новую кору, гиппокамп, подбугровую область, спинной мозг. Содержащие серотонин нейроны найдены также в кишечном нервном сплетении у некоторых позвоночных животных.
Ацетилхолин — единственный известный Медиатор, относящийся к простым эфирам (уксуснокислый эфир холина). Медиаторная функция ацетилхолина детально исследована на некоторых нейроэффекторных соединениях и межнейронных синапсах периферической нервной системы у позвоночных животных. Их периферические секреторные терминали происходят из следующих групп холинергических нейронов: клеток моторных ядер, иннервирующих скелетные мышцы; спинномозговых нейронов, иннервирующих хромаффинную ткань; преганглионарных нейронов, иннервирующих клетки интрамуральных и экстрамуральных ганглиев; значительной части периферических нейронов, в особенности интрамуральных ганглиев. Холинергические нейроны найдены у многих беспозвоночных, часть из них хорошо изучена (моторные нейроны стоматогастрической системы и некоторые афферентные нейроны ракообразных, интернейроны центральных ганглиев моллюсков, мотонейроны соматических мышц круглых и кольчатых червей и др.). Значительно хуже в связи с методическими трудностями идентификации холинергических нейронов исследованы интернейроны головного и спинного мозга. Данные, на основании которых идентификация холинергических нейронов базировалась на гистохим, выявлении ацетилхолинэстеразы, следует считать в основном ошибочными.
(лат. mediator посредник: синоним нейромедиаторы)
Участие М. в передаче нервного импульса представляется следующим образом. Специализированный для секреции М. участок пресинаптической клетки имеет особую наружную так называемую секреторную мембрану, которая при возбуждении пресинаптической клетки формирует мембранный пузырек, содержащий М. Содержимое пузырька изливается затем в синаптическую щель, диффундирует к постсинаптической мембране, где взаимодействует с ее специфическими рецепторами. При изучении действия М. на рецепторы периферических органов и ц.н.с. выявлены различные типы рецепторов к одному и тому же медиатору (м-, н-холинорецепторы, α-, β-адренорецепторы и др.). Их разделение основано на особенностях биохимических реакций, протекающих в системе медиатор — рецептор. Например, в м-рецепторах реакция носит мускариноподобный характер (они не чувствительны к яду кураре), в н-рецепторах — никотиноподобный (чувствительны к яду кураре). Взаимодействие медиаторов с α-рецепторами вызывает эффект возбуждения (сужение сосудов, сокращение матки и т.д.): с β-рецепторами — тормозные эффекты (расширение сосудов, расслабление бронхов). Вместе с тем α- и β-рецепторы, расположенные в различных органах, могут по-разному реагировать на медиаторы. В зависимости от характера взаимодействия α- и β-рецепторов с различными М. эти рецепторы соответственно разделяют на α1-, α2-, β1- и β2-адренорецепторы.
Норадреналин (см. Симпатоадреналовая система) осуществляет медиаторную функцию в периферических нервных окончаниях и ц.н.с. Группы норадренергических нейронов имеются в среднем мозге, мосту мозга, продолговатом и промежуточном мозге. Норадреналин как медиатор воздействует на α-адренорецепторы (преимущественно на α1-адренорецепторы) и вызывает сильные сосудосуживающие прессорные и бронхолитические эффекты. Норадреналин в определенной степени взаимодействует и с β-адренорецепторами сердца, что сопровождается увеличением сердечного выброса, возрастанием потребности миокарда в кислороде.
Адреналин длительное время относили к М. Однако доказано, что нейромедиатором является его предшественник — норадреналин, а сам адреналин играет главным образом роль гормона, влияющего на обмен веществ. Вместе с тем адреналин способен оказывать смешанное действие, влиять одновременно на α- и β-адренорецепторы.
Несмотря на то, что дофамин, норадреналин и адреналин обладают самостоятельной медиаторной функцией, их действие взаимосвязано. что обусловлено общностью происхождения. Исходным продуктом этих М. является аминокислота тирозин. Из триптофана образуется другой М. — серотонин. Нервные клетки, содержащие его (серотонинергические нейроны), найдены в коре головного мозга, гиппокампе, гипоталамусе, стволе мозга, спинном мозге. Помимо структур центральной и периферической нервной системы серотонин содержится в различных органах, тканях и клетках, в т.ч. тромбоцитах, клетках кишечника, мозгового вещества надпочечников. В нервной системе серотонин накапливается в цитоплазме нервных окончаний, выделяясь из синаптических пузырьков под влиянием нервных импульсов и взаимодействуя со специфическими рецепторами (серотонинергические рецепторы). Различают три основных типа серотониновых рецепторов — М-, D- и Т-рецепторы. М-рецепторы локализованы в ц.н.с., D-рецепторы — в ц.н.с. и гладких мышцах, Т-рецепторы — в окончаниях афферентных нервов. Периферическое действие серотонина характеризуется стимуляцией сокращения гладкой мускулатуры (матки, кишечника, бронхов), сужением кровеносных сосудов. Серотонин является также одним из медиаторов воспаления.
Ацетилхолин принимает участие в передаче нервного возбуждения в ц.н.с., вегетативных узлах, окончаниях парасимпатических и двигательных нервов. Относясь также к биогенным аминам и являясь уксуснокислым эфиром холина, ацетилхолин быстро (через 1—2 мс) разрушается с образованием холина и уксусной кислоты. Образовавшиеся продукты распада вновь поступают внутрь нервных окончаний, ресинтезируются и снова способны участвовать в проведении нервного импульса. Холинорецепторы локализованы на внешней стороне постсинаптической мембраны. Периферическое мускариноподобное действие ацетилхолина проявляется замедлением сердечных сокращений, расширением кровеносных сосудов и понижением АД, усилением перистальтики желудка и кишечника, желчного и мочевого пузыря, матки, возрастанием секреции пищеварительных, потовых и слезных желез и т.д.
Периферическое никотиноподобное действие ацетилхолина обусловлено его участием в передаче нервных импульсов с преганглионарных волокон на постганглионарные в вегетативных узлах, а также с двигательных нервов на поперечнополосатую мускулатуру, В больших дозах ацетилхолин способен блокировать передачу возбуждения. Аналогично и центральное действие этого М. В малых дозах ацетилхолин участвует в проведении возбуждения в разные отделы мозга, в больших — тормозит синаптическую передачу.
Функции М. выполняет ряд аминокислот. В частности, глутаминовая кислота является самым распространенным М. в центральной нервной системе, особенно в передних отделах головного мозга; в зависимости от типа рецепторов может оказывать либо возбуждающее, либо тормозящее действие. Предшественник глутаминовой кислоты — гамма-аминомасляная кислота (ГАМК) также обладает свойствами М. Специфические ГАМК-ерические рецепторы тесно связаны с дофаминовыми и другими рецепторами мозга.
Большую группу М. составляют нейропептиды (см. Регуляторные пептиды), представляющие собой короткие цепочки из аминокислотных остатков (от двух до 39). Многие из них ранее рассматривались только как гормоны гипофиза, местные гормоны кишечника или гормоны гипоталамуса, регулирующие выделение других гормонов (см. Гипоталамические нейрогормоны).
Наряду с прямым возбуждением или торможением клетки-мишени медиаторы в ряде случаев воздействуют на нервное окончание, усиливая и уменьшая выход из него других медиаторов. Было принято считать, что отдельная нервная клетка секретирует только один М. (принцип Дейла). Однако обнаружена способность одних и тех же клеток синтезировать М. разных типов. Чаще всего отмечаются Следующие сочетания секреций одной и той же клеткой: классические медиаторы и нейропептиды (серотонин + вещество Р, серотонин + тиротропин, норадреналин + соматостатин, норадреналин + энкефалин, норадреналин + панкреатический полипептид, дофамин + холецистокинин, ацетилхолин + вазоактивный кишечный полипептид).
Многие лекарственные средства оказывают лечебное действие, влияя специфическим образом на передачу возбуждения в окончаниях периферических нервов или в центральной нервной системе (см. Адреноблокирующие средства, Адреномиметические средства, Холиномиметические средства).
Для изучения М. вначале была разработана методика выделения из тканей мозга так называемым синаптосом. Это дало возможность изучать механизмы синаптической передачи вне организма. Для изучения функции М. используют также методы избирательного окрашивания М. в нервных клетках. Много ценных данных дает изучение вводимых извне в мозг или отдельную клетку М. с радиоактивной меткой. Перспективны также методы выработки антител против М. или ферментов, участвующих в синтезе или распаде медиаторов.
Медиаторы аллергических реакций — биологически активные вещества, освобождающиеся или создающиеся в результате образования комплекса аллергена с соответствующими антителами или с антигенсенсибилизированными Т-лимфоцитами и имеющие важное значение в патогенезе аллергии. Характер действия М. зависит от типа реакции, ее иммунологических механизмов, вида аллергена.
При аллергической реакции немедленного типа (см. Аллергия) клетками-мишенями являются тучные клетки (лаброциты) и базофильные лейкоциты, имеющие F-рецепторы к антителам классов lgE и lgG4. При соединении аллергена с этими антителами, фиксированными на клетках, наступает дегрануляция и высвобождение из них М. Регуляторами являются внутриклеточные нуклеотиды — циклический аденозинмонофосфат (цАМФ), гуанозинмонофосфат (цГМФ) и ионы кальция. Образование цАМФ и цГМФ происходит под влиянием ферментов — аденилциклазы (АЦ) и гуанилциклазы (ГЦ), а инактивация осуществляется фосфодиэстеразами (ФДЭ). цАМФ препятствует образованию и выделению медиаторов, а цГМФ способствует им. Эти процессы идут с участием адренергических и холинергических рецепторов кортикостероидных гормонов (Кортикостероидные гормоны) и простагландинов (Простагландины) (ПГ). Из тучных клеток и базофилов выделяется гистамин — продукт превращения гистидина. Действуя через гистаминовые рецепторы I типа (Н—1), он вызывает сокращение гладкомышечных органов, расширение сосудов и повышение их проницаемости, снимает кровяное давление, вызывает отек, зуд. При этом повышается содержание гастамина в крови, а также чувствительность к нему клеток органов-эффекторов. В дальнейшем гистамин препятствует освобождению М. из тучных клеток, действуя через гистаминовые рецепторы II типа (Н—2). Естественная способность белков сыворотки связывать гистамин (гистаминопексия) при аллергии пропадает. Для определения гистаминопексии устанавливают гистаминопектический индекс (ГПИ). В норме он составляет 30—40%, при аллергии — ниже 10%. Гистаминопексия является неспецифическим показателем и имеет большое значение для оценки эффективности лечения в динамике.
Медиаторы аллергических реакций цитотоксического типа образуются при участии антител классов ИГ ГИГ М, способных связывать комплемент. Продукты его активации вызывают повышение проницаемости сосудов, хемотаксис нейтрофилов, повреждение мембран, лизис клеток.
Медиаторы аллергических реакций типа Артюса высвобождаются при образовании циркулирующих иммунных комплексов (ИК) с участием lgG и lgM и их последующей фиксацией в тканях. ИК проявляют повреждающее действие при участии комплемента. В реакции принимают участие полиморфно-ядерные лейкоциты, освобождающие лизосомальные ферменты. В процесс вовлекается также калликреин-кининовая система, т.к. при действии ИК активируется фактор Хагемана (XII фактор свертывания крови) — один из активаторов калликреина. Калликреин образуется в крови и тканях из прекалликреина, активация которого может быть вызвана комплексом антиген — антитело. Калликреин обнаруживают также в тучных клетках. В плазме содержится вазоактивный полипептид брадикинин. Он вызывает сокращение гладкомышечных органов, расширение мельчайших артериол и капилляров, увеличение их проницаемости, диапедез лейкоцитов, развитие аллергического воспаления, осложнений и пролонгирование астматических приступов. Основные медиаторные влияния — активация комплемента, калликреин-кининовой системы, действие лизосомальных ферментов.
Медиаторы аллергических реакций замедленного типа (ГЗТ) — лимфокины, высвобождаемые сенсибилизированными Т-лимфоцитами при контакте с антигеном. Лимфокины способствуют концентрации в очаге аллергической реакции различных клеточных элементов, развитию инфильтрации и аллергического воспаления. Кожно-реактивный фактор увеличивает проницаемость сосудов, усиливает миграцию лейкоцитов и мононуклеаров. Сходное действие оказывает фактор проницаемости. Хемотаксический фактор притягивает в очаг реакции нейтрофилы, эозинофилы, несенсибилизированные лимфоциты, моноциты. Миграцию ингибирующий фактор (МИФ) вызывает задержку и накопление макрофагов в зоне аллергической альтерации, участвует в образовании гранулем при инфекционно-аллергических заболеваниях. Выделяется фактор, ингибирующий миграцию лейкоцитов. К митогенным факторам относят кроме лимфоцитарного фактора интерлейкины, выделяемые макрофагами и Т-хелперами; лимфотоксический фактор оказывает цитотоксическое действие на клетки-мишени, способствует развитию некроза при ГЗТ. Фактор цитотоксичности увеличивает цитотоксичность макрофагов. Фактор переноса обеспечивает перенос специфической активности на несенсибилизированные Т-лимфоциты, вовлекая их в реакцию ГЗТ. Лимфоцитами выделяется иммунный интерферон, который не только проявляет антивирусные свойства, но и влияет на активность естественных киллеров. В механизме образования лимфокинов имеет значение соотношение цАМФ и цГМФ. В реакции ГЗТ участвуют лизосомальные ферменты, освобождаемые при разрушении клеток в очаге воспаления, и кинины.
Принципы фармакотерапии патохимической стадии аллергических реакций основаны на подавлении синтеза медиаторов, процессов их высвобождения из клеток, угнетении влияния на органы-эффекторы (см. Противоаллергические средства).
Библиогр.: Адо А.Д. Общая аллергология, М., 1978; Гущин И. С Немедленная аллергия клетки, М., 1976: Иммунология, под ред. У. Пола, пер. с англ., т. 1, с. 437. М., 1987; Либерман Ф.Л. и Кроуфорд Г.В. Лечение больных аллергией, пер. с англ., с. 103. М., 1986; Медуницын Н.В. Повышенная чувствительность замедленного типа, с. 41, М., 1983; Мозг, пер. с англ., под ред. П.В. Симонова, с. 148, М., 1984; Пыцкий В.И., Адрианова Н.В. и Артомасова А.В. Аллергические заболевания, с. 29. М., 1984; Физиология человека, под ред. Р. Шмидта и Г. Тевса, пер. с англ., т. 1, с. 99, М., 1985; Ялкут С.И. и Котова С.А. Циклические нуклеотиды и особенности гомеостаза при аллергии, с. 47, Киев, 1987.
Читайте также:
