
Какие нейроны входят в состав нервных центров
Определение понятия нервного центра
Нервный центр — это совокупность связанных между собой нейронов, совместно выполняющих определённую функцию путём преобразования входящего возбуждения в выходящее с изменёнными характеристиками.
Данное определение содержит 7 критериев нервного центра, найдите и назовите их.
Суперкраткое определение: Нервный центр - это "полисинаптический трансформатор возбуждения".
Нервный центр - это та нервная структура, которая связывает сенсорную систему с эффекторной и преобразует сенсорное возбуждение в эффекторное или модулирующее.
Нервный центр - это многозначное понятие.
Анатомический подход: нервный центр - это совокупность сходных нервных клеток, выполняющих общую функцию и компактно расположенных в определенном участке ЦНС.
Здесь используется морфологический подход, т.е. нервные центры определяются по строению. Нервные клетки, образующие такой нервный центр, соединены в локальные компактные структуры: нервные узлы (ганглии) в периферической нервной системе или мозговые ядра в центральной нервной системе.
Физиологический подход (по деятельности) : нерный центр - это система взаимосвязанных клеток, которые объединяются для выполнения определённой функции, а физически могут находится в различных местах нервной системы.
Такое определение созвучно понятию "функциональной системы", которое было предложено П.К. Анохиным. Но если функциональная система подразумевает временное объединение нейронов, то нервный центр - это обычно устойчивое образование. По П.К. Анохину различные анатомические нервные центры при необходимости могут на время объединяться в функциональную систему для получения определенного полезного результата.
Нервные центры биорегуляции. В функциональном отношении нервный центр также может представлять собой сложное объединение нескольких анатомических нервных центров, расположенных в разных отделах ЦНС и обусловливающих сложные рефлекторные акты. В этом смысле говорят о "пищевом нервной центре", "болевом нервном центре" и т.п. Это центры регуляции функций организма.
Понятие нерный центр может также пересекаться с понятием доминанты. Доминанта - это устойчивый очаг возбуждения, подчиняющий себе другие очаги . Кроме единичного очага возбуждения в состав доминанты также могут входить и другие связанные с ним очаги. Создатель учения о доминанте А.А. Ухтомский называл такие объединения возбуждённых очагов "созвездиями" ("констелляциями").
Эволюция выбирает путь концентрации нервных элементов и увеличение количества связей между ними. Поэтому из рассеянных диффузно нервных клеток образуются компактные нервные центры.
Линия эволюционного развития структуры нервной системы
1) Отдельные нервные клетки соединены примерно равными связями.
2) Нервные цепи - нервные клетки соединены последовательно более сильными связями, образуя предопределённый путь для движения возбуждения от одного пункта к другому.
3) Нервные сети - нервные клетки соединены в виде решеток с неравноценными связями.
4) Нервные узлы (ганглии) - нервные клетки собраны в компактные структуры, соединёнными между собой продолными связями (коннективами) и поперечными (комиссурами) в виде лестницы.
5) Нервная трубка - нервные клетки расположены в виде сплошных слоёв, закрученных в виде трубы.
6) Нервные ядра - обособленные нейронные структуры из тесно связанных сходных между собой нейронов в составе нервной трубки, специализирующиеся на определённых функциях.
7) Нейронные поля — зоны коры головного мозга. Они состоят из вертикальных колонок, в которые сгруппированы нейроны.
Свойства нервных центров
Наличие входов и выходов для возбуждения . В нервном центре можно различить приносящие (афферентные) входы и выносящие (эфферентные) выходы.
Одностороннее проведение возбуждения. Это свойство отдельного синапса и нервной цепи. В нервном центре может быть множество путей между входами и выходами. За счёт обратных связей возможно возвратное движение возбуждения. Но это происходит внутри нервного центра. А если рассматривать нервный центр целиком, то возбуждение приходит внего по приходящим путям, а выходит по эфферентным выходящим. Таким образом, можно говорить об одностороннем проведении возбуждения нервным центром.
З адержка ( замедление ) проведения возбуждения . В нервных центрах возникает задержка в проведении возбуждения, так называемый латентный (скрытый) период. Задержка обусловлена синаптической передачей возбуждения. Чем больше синапсов участвует в проведении возбуждения, тем более длительной получается задержка.
Суммация возбуждения . Если одновременно подавать возбуждение на несколько входов нервного центра, то на выходе можно получить более сильное возбуждение. Свойством суммации обладает и отдельный нейрон за счёт суммации локальных потенциалов.
Трансформация (преобразование) входящего возбуждения в иное - выходящее . Нервный центр осуществляет изменение, перекодирование поступающих в него потоков импульсов. Трансформация возбуждения - это, пожалуй, самое главное свойство нервного центра. Наиболее известное свойство из этого ряда – трансформация ритма. Нервный центр получает на входе один ритм импульсации, а на выходе дает другой (более медленный или более частый).
Последействие (облегчение) . Это означает, что после возбуждения нервного центра он некоторое время ещё сохраняет повышенную возбудимость. Поэтому последующее возбуждение даёт более сильный эффект и получение эффекта от работы нервного центра облегчено.
Утомляемость и низкая лабильность. Лабильность - это предельная частота импульсации, доступная данной нервной структуре. Нервные центры могут пропускать через себя потоки возбуждения с ограниченной частотой импульсации вследствие задержки передачи возбуждения, которая происходит в многочисленных синапсах. Повышенная утомляемость нервных центров объясняется высокой утомляемостью синапсов и ухудшением метаболизма (обменнных процессов) в нейронах после нагрузки.
Тонус . Это означает, что даже без внешнего воздействия нервный центр сохраняет определённый уровень возбудимости и самостоятельно поддерживает у себя определённый уровень возбуждения.
Чувствительность к кислороду и к действию биологически активных веществ (нейротропных). Это создаёт предпосылки к хеморегуляции - химическому управлению деятельностью нервного центра. Например, усиление или ослабление кровоснабжения изменяет работу нервных центров.
В озбудимость ( возбуждение ) . Это способность нервных центров переходить в более возбуждённое состояние, например, при внешнем воздействии на них (стимуляции) или под влиянием других нервных центров.
Торможение ("тормозимость") . Это способность нервных центров переходить в менее возбуждённое состояние, например, при внешнем воздействии на них или под влиянием других нервных центров.
Иррадиация возбуждения . Это "растекание возбуждения" по нервному центру, распространение возбуждения на новые участки от места его первоначального появления.
Конвергенция (схождение) . Это объединение двух или нескольких входящих потоков возбуждения в один выходящий поток. Т.е. в нервный центр входит больше потоков возбуждения, че выходит из него.
Дивергенция (расхождение) . Это разделение входящего потока возбуждения на несколько выходящих потоков. За счёт дивергенции получается, что в нервный центр входит меньше потоков возбуждения, чем выходит из него.
Окклюзия (запирание) . Это блокирование одним из входящих потоков возбуждения другого входящего потока. В результате выходящий поток возбуждения получается слабее, чем сумма этих входящих потоков.
Индукция (отдача) . Это наведение противоволожного (возбуждённого или тормозного) состояния на другие нервные центры или на себя самого. Для понятия индукции очень важно, что данной структурой наводится именно противоположное состояние, а не то, в котором находится она сама. Так, возбуждённая структура индуцирует торможение, а заторможенная - возбуждение.
Автоматия (спонтанная активность, автономность) нервных центров. Это означает, что даже без внешнего воздействия нервный центр может самостоятельно порождать возбуждение на выходе или поддерживать свой тонус (как бы развлекать сам себя). Объясняется это свойство нервного центра существованием в нём специальных нейронов-пейсмекеров (водителей ритма). В них самопроизвольно возникает возбуждение, независимо от работы их афферентных входов. Таким образом, в нервных центрах может происходить периодическая или постоянная генерация (порождение) нервных импульсов, которые возникают даже при отсутствии входящего возбуждения. Самопроизвольная импульсация пейсмекеров обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов.
Реципрокные (взаимоисключающие) отношения. Это означает, что возбуждение одного нейрона (или центра) подавляет работу другого, связанного с ним, нейрона (или центра).
П ластичность . Это способность перестраивать свою структуру и\или деятельность под влиянием предыдущей деятельности. Пластичность - это одно из важнейших свойств биологических систем, которое отличает их от технических систем.
Адаптация. Нервный центр способен приспосабливаться к новой нагрузке и новым условиям работы.
Компенсаторные возможности. При частичном повреждении нервный центр продолжает свою деятельность за счёт сохранившихся нейронов. Для этого он использует свои способности к пластичности и адаптации.
Основные принципы в работе нервных центров
Принцип общего конечного пути ("воронка Шеррингтона"). Как правило, центры имеют больше афферентных входов, чем эфферентных выходов. Поэтому входящие потоки возбуждения конкурируют за выход, имея общий конечный путь. В итоге количество афферентных входов превышает количество эфферентных выходов.
Принцип обратной связи. Это означает, что последующий элемент (нейрон или центр) в последовательной цепи взаимосвязанных элементов влияет на состояние предыдущего элемента. Обратная связь позволяет произвести отладку взаимодействия между элементами и добиться их оптимального взаимодействия для достижения предельно возможного положительного результата в работе системы, состоящей из этих элементов.
Принцип доминанты. Это означает, что нервный путь или нервный центр наиболее активный получает преимущество по отношению к другим путям или центрам и начинает доминировать, господствовать над ними. Он тормозит их деятельность и перехватывает их возбуждение, чтобы усилить своё.
Принцип иерархии (соподчинения). Это означает, что одни элементы (нейроны и/или центры) подчиняются влиянию других элементов. Как правило, вышерасположенные центры подчиняют себе нижерасположенные центры.
Принцип пластичности. Это означает, что нервный центр перестраивает свою деятельность, приспосабливаясь к наилучшему выполнению своей функции для достижения общего конечного системного результата. Пластичность является важнейшей отличительной особенностью биосистем по сравнению с техническими системами.
Низшие нервные центры
Низшие нервные центры играют важную роль в работе любой сенсорной системы. Они являются одним из необходимых элементов сенсорной системы, по которому это понятие отличается от понятия "анализатор". Нервные центры не просто переключают возбуждение с одних нейронов на другие с помощью вставочных нейронов, т.е. выполняют "релейную" функцию, как это считалось ранее. Важно понять, что нервные центры занимаются трансформацией поступающего в них возбуждения, т.е. его преобразованием, или перекодированием. В результате этой трансформации входящее афферентное возбуждение превращается в выходящее эфферентное , отличающееся от входящего.
Работа (функции) низших нервных центров
1. Трансформация возбуждения , т.е.преобразование входящего потока сенсорного возбуждения в новый поток - выходящий. Выходящий поток может сильно отличаться от входящего, например, в том случае, если он должен управлять мышцами, а не строить нервную модель раздражения в виде сенсорного образа.
Виды трансформации возбуждения в нервном центре
1. Усиление.
2. Ослабление.
3. Блокировка.
4. Изменение паттерна (узора, характера).
5. Контрастирование границ в пространстве. Обычно оно достигается с помощью латерального (бокового) торможения. Латеральное торможение усиливает возбуждение по контуру раздражителя и рецептивного поля и ослабляет возбуждение в центральной области рецептивного поля.
6. Контрастирование границ во времени. Происходит за счёт преобразования тонического (постоянного) возбуждения в кратковременное фазическое. Таким способом отмечаются начало и конец действия раздражителя.
2. Распределение входящих потоков сенсорного возбуждения по выходящим потокам, которые направляются в различные нервные структуры. Эта функция нервного центра наглядно показана с помощью нашей схемы "Пути сенсорного возбуждения".
Виды распределения возбуждения в нервном центре
1. Конвергенция (схождение).
2. Дивергенция (расхождение).
3. Окклюзия (запирание).
4. Иррадиация (распространение).
3. Детекция . С помощью детекции выделяются раздражители с определёнными характеристиками за счёт срабатывания специальных нейронов-детекторов с соответствующими рецептивными полями. На любые другие раздражители, неадекватные для них, такие нейроны-детекторы не срабатывают, т.к. просто не возбуждаются такими стимулами.
Нервный центр –
это совокупность нейронов, расположенных на различных этажах ЦНС и регулирующих деятельность определённого органа.
В нервном центре выделяют рабочий отдел и надстройку. Надстройка представлена нейронами, расположенными в выше лежащих отделах ЦНС и управляющими деятельность рабочего отдела. Например: центр ССС деятельности состоит из нейронов, расположенных на уровне продолговатого мозга, гипоталамуса и коры головного мозга. Рабочим отделом являются нейроны в области продолговатого мозга. Нейроны в области гипоталамуса и коры головного мозга входят в состав надстройки.
Структурной частью нервного центра является нейрон (Рис.26). Он имеет тело неправильной формы и 2 вида отростков: многочисленные короткие дендриты и одиночный длинный отросток – аксон. По дендриту возбуждение идёт к телу нейрона. По аксону – от нейрона к рабочему органу или другому нейрону.

По функции нейроны делятся на:
- Чувствительные – афферентные, расположенные в спинномозговых ганглиях, ядрах черепных нервов, спинном и головном мозге.
- Двигательные – эфферентные, находящиеся в коре, подкоркой области, стволе головного мозга, передних рогах спинного мозга.
- Ассоциативные – вставочные. Объединяющие, передающие импульсы с афферентного на афферентные нейроны.
- Нейросекреторные ( например в гипоталамусе). Обладающие свойством вырабатывать и выделять в кровь гормоны, названные нейросекретами.
Место контакта двух нейронов называется СИНАПС.
Его формируют либо разветвление аксона и тело нейрона, либо аксон и дендрит. Строение синапса (Рис.27):

А. Пресинаптичекая мембрана ( мембрана, покрывающая окончание аксона в месте контакта).Пресинаптические окончания образуют бляшки, в которых находятся везикулы, содержащие медиатор. С помощью медиатора возбуждение передаётся через синаптическую щель на постсинаптическую мембрану.
Б. Синаптическая щель (расстояние между пре и постсинаптической мембранами 20 – 40 н/м.). В период возбуждения в синаптической щели появляются адгезивные белки, которые фиксируют положение пре и постсинаптических мембран и способствуют точной передачи медиатора.
В. Постсинаптическая мембрана (часть мембраны нейрона, содержащая рецепторы к медиатору). Медиаторы могут быть различными и возбуждающими и тормозными. Постсинаптическая мембрана является частью постсинаптической плотности, куда входят 1000-1500 белков (арматурные белки, белки цитоскелета, актомиозин).
Механизм передачи возбуждения в синапсе
(на примере медиатора ацетилхолина). Импульс возбуждения, подошедший к пресинаптической мембране, увеличивает её проницаемость для ионов кальция, который входит в синаптическую бляшку, связывается с белком и возникает выброс медиатора из бляшки. Квант медиатора проходит через синаптическую щель, контактирует с рецетором постсинаптической мембраны, увеличивает частично её проницаемость для ионов натрия и вызывает частичную деполяризацию на 3-5 мв. (возбуждающий постсинаптический потенциал или ВПСП). Чтобы получить возбуждение в нервном центре, необходимо суммировать 3-5 таких ВПСП для достижения критического уровня деполяризации (Рис.28). Для этого необходимо к пресинаптической мембране подвести минимум три импульса возбуждения и выбить 3 кванта медиатора.
Потенциал действия возникает в аксонном холмике нейрона.

Генерация потенциала действия в нейроне прекращается ретрограднеой сигнализацией (Рис.29). Во время возбуждения нейрона активируются дополнительно метаботропные рецепторы, которые увеличивают проницаемость постсинаптической мембраны для ионов кальция. Кальций, попав в цитоплазму нейрона, активирует фосфолипазу, которая выщепляет арахидоновую кислоту из мембраны. Из неё образуется медиатор 2-АГ

(2-арахидоноилглицерин), который транспортируется через синаптическую щель назад к пресинаптической мембране, находит для себя рецептор КБ-1 (канабиоидные). Возбуждение этих рецепторов блокирует ток кальция в пресинаптической мембране и медиатор из везикул не выделяется. Генерация потенциала действия в нейроне прекращается. Таким механизмом регуляруется частота нервных импульсов в нейроне.
- Односторонняя передача возбуждения в синапсе ( от пре к постсинаптической мембране ). В обратном направлении передача невозможна, ибо медиатор содержится только в пресинаптических бляшках, а рецептор к нему только на постсинаптической мембране.
- Синаптическая задержка. В синапсе передача возбуждения задерживается на 0,6 – 0,8 сигм ( 1 сигма=0,001 сек.). Время уходит на выделение медиатора, транспорт его через синаптическую щель, контакт с рецептором и суммацию ВПСП.
- Суммация ВПСП.
А. Последовательная, когда последователь по времени суммируются ВПСП.
Б. Пространственная (одновременная). На одном нейроне может контактировать несколько аксонов. Из каждого одновременно выделится по кванту АХ, которые сразу вызовут падение мембранного потенциала в нейроне до критического уровня (Рис.30).

- Высокая утомляемость нервного центра.
При длительном действии раздражителя расходуется АХ в пресинаптической области и не возбуждается постсинаптическая мембрана. Восстанавливается работоспособность утомлённого нервного центра за счёт отдыха. Он может быть активным и пассивным. Пассивный отдых. Когда ничего не делая ожидается время синтеза достаточного количества АХ в пресинаптической бляшке. Активный отдых. Для этого необходимо заняться другим видом работы и подключить к возбуждению параллельную рефлекторную дугу. От неё по коллатерали возбуждение подойдёт к утомлённому нервному центру и выбросит в синапс недостающее количество АХ. ВПСП одновременно будет суммироваться на нейроне из нескольких синапсов. Этот вид отдыха более продуктивен, восстановление трудоспособности наступает быстрее, чем при пассивном отдыхе (Рис.31). 
- ТРАНСФОРМАЦИЯ ритма. Нервный центр может изменять число нервных импульсов, подошедших к нему в область пресинаптичеческих бляшек, увеличивая или уменьшая частоту (трасформируя). Поэтому трансформация может быть как поышающая, так и понижающая. ПОНИЖАЮЩАЯ – возникает за счёт суммации ВПСП
- ПОСЛЕДЕЙСТВИЕ – способность нервного центра генерировать возбуждение после действия раздражителя. Это может быть связано с действием гуморальных раздражителей, способных вызывать деполяризацию или с наличием возвратных коллатералей. По ним импульсы возвращаются к нейрону и его возбуждают (Рис.34).
- ВЫСОКАЯ ЧУВСТВИТЕЛЬНОСТЬ нервного центра к биологически активным веществам (БАВ). В крови существуют соединения, способные увеличивать проницаемость мембраны нейрона к ионам натрия, вызывая деполяризацию. При небольшой их концентрации возникает частичная деполяризация, что определяет тонус нейрона и его готовность к ответной реакции. Это важно для поддержания гомеостаза в организме. При высокой концентрации БАВ в крови может возникнуть самовозбуждение нейронов без действия раздражителей.
Нервный центр обладает высокой чувствительностью к недостатку кислорода. Нейроны коры головного мозга способны существовать без кислорода не более 3-5 минут и этим определяется длительность клинической смерти. Нейроны нижележащих отделов ЦНС могут существовать без кислорода несколько дольше.
- НИЗКАЯ ЛАБИЛЬНОСТЬ нервного центра. Лабильность – это функциональная активность. К нервному центру подходит до 500 импульсов, а он может пропустить 100-120 импульсов. Это связано с последовательной суммацией ВПСП, когда частота импульсов теряется. Низкая лабильность предохраняет рабочий орган от перегрузок.
Нервный центр (НЦ) – объединение нейронов, координирующих и регулирующих в составе функциональной системы (П. К. Анохин) активность различных физиологических эффекторов, деятельность которых обеспечивает стабильность констант внутренней среды организма (рис.1). Например, дыхательный центр – объединение нейронов ствола мозга, регулирующих работу дыхательной мускулатуры для поддержания стабильности газовой константы организма; сердечно-сосудистый центр – объединение нейронов продолговатого мозга, регулирующих работу сердца, тонуса сосудов для обеспечения стабильности константы оптимального кровяного давления; терморегуляторный центр – нейронный комплекс гипоталамической области, координирующий и регулирующий работу множества эффекторов, активность которых поддерживает постоянство температурной константы организма.
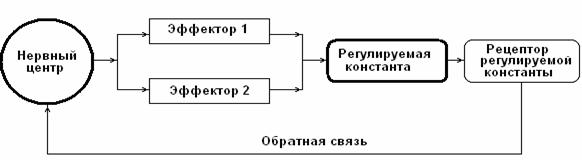
Рис. 1. Принципиальная схема функциональной системы (ФС) организма. В состав ФС входит нервный центр (НЦ), координирующий работу периферических эффекторов, деятельность которых определяет статус жизненно-важной константы. Качество координирующих и регулирующих влияний НЦ на эффекторы зависит от активности каналов обратной связи
Элементарной единицей НЦ является нейронный модуль – совокупность входных и вставочных нейронов, замыкающихся на один выходной (моторный), или конечный нейрон. НЦ обычно состоят из большого числа модулей, функционально связанных друг с другом и работающих параллельно друг с другом. На рис. 2 представлена схема простейшего НЦ, в состав которого входят 2 модуля. Каждый отдельно взятый нейронный модуль демонстрирует активность, характерную для всего НЦ.
Свойства нервных центров. Нервные центры (НЦ) обладают рядом характерных свойств, присущих всем нервным центрам. От чего зависят эти свойства?
1. От свойств одиночных нейронов, входящих в состав НЦ. 2. От свойств химических синапсов, обеспечивающих передачу влияний одних нейронов на другие. 3. От особенностей связей между отдельными нейронами в составе нейронного модуля. В простейшем варианте связи могут быть линейными, однако чаще в составе НЦ нейроны формируют сети сложной конфигурации.
Частные свойства нервных центров (НЦ).
1. НЦ проводит возбуждение только в одну сторону - от входных нейронов к выходным. Данное свойство НЦ определяется как односторонняя проводимость – от входных нейронов НЦ к выходным. Односторонняя проводимость НЦ объясняется тем, что химические синапсы (синапс 1, синапс 2 - рис. 3) проводят возбуждение только в одну сторону – от пресинаптической мембраны к мембране постсинаптической. Одностороннюю проводимость НЦ можно доказать в простом эксперименте с раздражением задних и передних корешков спинного мозга. Если раздражать электрическим током задние корешки, от передних корешков всегда можно зарегистрировать потенциалы действия. Это свидетельствует о проведении возбуждения через НЦ от входных нейронов к выходным. Если раздражать передние корешки, от задних потенциалы действия никогда не регистрируются. Это свидетельствует о том, что от выходных нейронов к входным возбуждение в НЦ не проводится.
2. Возбуждение в нервных центрах (НЦ) распространяется с медленной скоростью. Данное свойство определяется как замедленное распространение возбуждения в НЦ и в целом в ЦНС.
По периферическим нервным волокнам возбуждение распространяется быстро (по аксонам чувствительных нейронов возбуждение распространяется со скоростью 50-70 м/с). Однако, как только возбуждение поступает в НЦ скорость его распространения снижается. Это связано с феноменом синаптической задержки. Известно, что для проведения возбуждения через один химический синапс времени требуется примерно 2 мс (суммарная синаптическая задержка). Следовательно, если возбуждение в НЦ проходит несколько синапсов, средняя скорость проведения возбуждения в НЦ оказывается сниженной по сравнению со скоростью проведения возбуждения в нервных волокнах.

Рис. 2. Принципиальная схема простейшего нервного центра (НЦ), в состав которого входят два модуля (связи между ними – не показаны). Обращает на себя внимание тот факт, что в конечном итоге все входные возбуждения к НЦ сходятся (феномен конвергенции) на одном выходном нейроне модуля НЦ. Выходной нейрон, интегрируя (объединяя) входные потоки, формирует один выходной поток возбуждения, который поступает к периферическому эффектору.
Хорошо известно, что рефлекторные реакции, т. е. изменения активности периферических эффекторов в результате раздражения сенсорных рецепторов и проведения афферентного возбуждения через НЦ, возникают с некоторым скрытым (латентным) периодом. Латентный период – это время от момента нанесения раздражения до появления ответной реакции. Значительную часть латентного периода рефлекторных реакций составляет время проведения возбуждения через НЦ. Последнее определяется как центральное время рефлекторного акта.
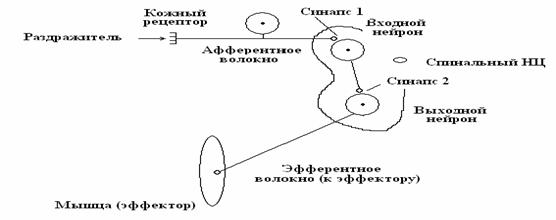
Рис. 3. Схема проведения афферентного возбуждения через спинальный НЦ к скелетной мышце – от входного нейрона к выходному, в обратном направлении возбуждение не проходит. Причина – химические синапсы (1, 2) обладают свойством односторонней проводимости.
3. Возбуждение в ЦНС может широко иррадиировать (перемещаться) от одного нервного центра (НЦ) к другим, часто расположенным всамых отдаленных зонах ЦНС.
Иррадиация возбуждения в ЦНС объясняется ветвлением аксонов выходных нейронов НЦ, связывающих соседние НЦ. При этом создаются возможности для распространения возбуждения по ЦНС (рис. 4). Беспрепятственную иррадиацию возбуждения в ЦНС ограничивают тормозные нейроны.

Рис. 4. Иррадиация возбуждения от НЦ 1 к другим НЦ в составе ЦНС. В целях упрощения нервные центры представлены лишь выходными нейронами.
Примеры макроорганизации иррадиации периферического возбуждения по коре головного мозга.
Горизонтальная иррадиация. Возбуждение от кожных болевых рецепторов поступает по спиноталамическому тракту, формируя первичный очаг в коре головного мозга. От места возникновения первичного очага возбуждение по горизонтальным ассоциативным связям может иррадиировать по коре головного мозга (рис. 5).
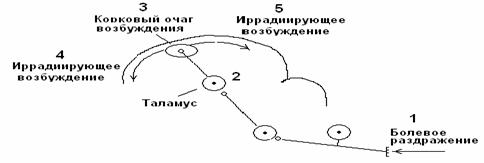
Рис. 5. Механизм горизонтальной иррадиации периферического возбуждения по коре головного мозга.
Вертикальная иррадиация. Возбуждение от кожных болевыхрецепторов по спиноталамическому тракту поступает в кору головного мозга и по стволовым коллатералям – в стволовую ретикулярную формацию. По многочисленным вертикальным ретикуло-кортикальным трактам возбуждение из ретикулярной формации вторично иррадиирует по коре головного мозга. Данный механизм иррадиации периферического возбуждения по коре головного мозга является основным (рис. 6).
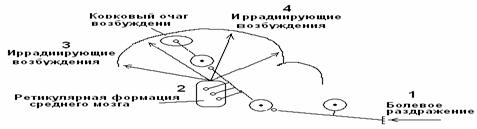
Рис. 6. Механизм вертикальной иррадиации периферического возбуждения по коре головного мозга.
Возвратная иррадиация. Первичное возбуждение в коре головного мозга по корково-ретикулярным связям может активировать нервные центры восходящей ретикулярной формации ствола мозга, при этом возбуждение из ретикулярной формации вторично перемещается в кору по множественным вертикальным ретикуло-корковым ткактам, иррадиируя по различным корковым центрам (рис. 7).

Рис. 7. Механизм возвратной иррадиации возбуждения по коре головного мозга.
На рис. 8 представлена нейронная схема НЦ, обеспечивающая режим усиления, или мультипликации (умножения) входного сигнала.
На вход НЦ поступает 2 нервных импульса, на выходе – регистрируем 6. Феномен усиления входной импульсации осуществляется в результате ее мультипликации (умножения) на нейронах НЦ, связанных в конечном итоге с нейроном выходным.

Рис. 8. Схема нейронной сети НЦ, работающей в режиме усилителя входного сигнала в результате его мультипликации (умножения) на нейронах 1, 2, 3.
На рис. 9 представлена нейронная сеть НЦ, обеспечивающая ослабление (редукцию) входного сигнала. Рассмотрим простейшую ситуацию. Линейная цепь А-В не выполняет преобразовательную работу. Добавим в нее один элемент – тормозный нейрон С, возбуждаемый выходными сигналами клетки В. Аксон тормозного нейрона соединен с выходным нейронов В по схеме возвратного торможения. Что наблюдается на выходе НЦ, если на входной нейрон А непрерывно будет поступать нервная импульсация ?

Рис. 9. Нейронная сеть нервного центра с тормозным нейроном, включенным по схеме возвратного торможения. В целом нервная сеть НЦ работает как аттенюатор – выходной поток нервных импульсов слабее входного.
Клетка А принимает поток входных импульсов, передавая их на нейрон В, который работает в паре с тормозным нейроном С. Что делает клетка В? Она отвечает на 1-й, 2-й, 3-й импульсы, поступающие к ней от нейрона А. Однако параллельно возбуждение идет к тормозному нейрону С. Нейрон С возбуждается, при этом тормозный медиатор в окончаниях аксона С тормозит активность нейрона В. Нейрон В перестает реагировать на импульсацию от нейрона А, следовательно, в структуре выходного потока клетки В появляется пауза. Однако, как только клетка В перестанет работать под влиянием нейрона С, последний сам перестает работать, т. к. к нему перестает поступать возбуждающая импульсация от нейрона В. Это немедленно восстанавливает возбудимость нейрона В, и он вновь начинает реагировать на импульсацию от нейрона А. Далее циклы повторяются, при этом выходной поток НЦ, определяемый активностью нейрона В, будет носит форму, в которой вспышки активности чередуются с тормозными паузами (рис. 9). В целом выходная импульсация НЦ будет редуцирована в сравнении с исходной.

Рис. 10. Схема нейронной сети НЦ с тормозным нейроном, включенным по схеме параллельного торможения.
Еще один пример с участием тормозного нейрона. Что будет на выходе НЦ, если тормозная клетка включается в нейронную сеть по схеме параллельного торможения?
Под влиянием начальной порции возбуждения выходной нейрон В генерирует несколько импульсов. Однако через коллатераль аксона А параллельно активируется тормозный нейрон С, который тормозит клетку В. Ее активность тормозится. В итоге – на выходе наблюдаем всего два импульса в сравнении с шестью ПД на входе (рис. 10).
5. Нервным центрам (НЦ) присуще явление последействия, под которым понимают повышенную активность нейронов НЦ на протяжении некоторого времени после прекращения поступления нервных импульсов на вход НЦ.
Как этот феномен объяснить с позиций нейрофизиологии? Линейная цепь нейронов А-В последействие не обеспечивает. Однако ситуация изменяется, когда между нейронами А и В встраивается замкнутая сеть вставочных нейронов 1, 2, 3 (рис. 11).

Рис. 11. Схема нейронной сети, обеспечивающей феномен последействия в НЦ за счет циркуляции (реверберации) нервных импульсов в цепи вставочных нейронов 1, 2, 3, замыкающихся на нейрон А. Цепь вставочных нейронов загружается импульсацией при поступлении к нейрону А входной (афферентной) импульсации (1). В дальнейшем (2) входная импульсация воспроизводится непрерывно в замкнутой цепи нейронов 1, 2, 3, далее нейрон А и выходной нейрон В.
6. Нервные центры обладают способностью суммировать возбуждения, поступающие к ним по афферентным каналам. Суммация может быть двух видов: временная и пространственная.
Временная, или частотная суммация (рис. 12). К нейрону (А) НЦ в результате воздействия на входной нерв подходят друг за другом два нервных импульса. Каждый из импульсов вызывает генерацию ВПСП, недостаточную для выведения мембранного потенциала нейрона до КУД (В). Однако если второй импульс приходит к нейрону на фоне развития ВПСП от первого, ВПСП от второго импульса, суммируясь с ВПСП от первого, смещает мембранный потенциал до КУД, нейрон НЦ при этом возбуждается. В рассмотренном примере суммация двух входных подпороговых воздействий к НЦ является производным фактора времени между входными воздействиями. При некотором оптимальном временном интервале два подпороговых воздействия могут суммироваться на нейронах НЦ.

Рис. 12. Временная (частотная) суммация входных воздействий на нейроне НЦ.
Пространственная суммация (рис. 13). К нейрону (А) по различным входным нервам поступают два нервных импульса. Каждый вызывает подпороговый ВПСП (В). Однако если нервные импульсы поступают одновременно – подпороговые ВПСП суммируются (С), при этом мембранный потенциал нейрона смещается до КУД, что приводит к возбуждению нейрона НЦ.

Рис. 13. Пространственная суммация входных воздействий на нейроне НЦ, когда два импульса подходят к нейрону одновременно по различных афферентным проводникам (феномен конвергенции).
7. Нервные центры (НЦ) обладают способностью перестраивать свою работу в зависимости от изменения активности сопряженных нервных центров, изменения качества афферентных воздействий на НЦ. Данное свойство НЦ определяется как пластичность.
Как доказать феномен пластичности НЦ? На рис. 14-1 представлена упрощенная схема моторных центров спинного мозга А и В, осуществляющих контроль за активностью периферических мышц конечностей А и В. Перережем эфферентные (моторные) нервы и соединим их по схеме – нерв А – с мышцами В, нерв В с – мышцами А (рис. 14-2). Спустя некоторое время функциональная целостность нервов восстанавливается, однако моторная активность подопытного животного после вмешательства грубо нарушается. Животное теряет способность нормально передвигаться. Тем не менее, после многочисленных попыток перемещения в среде обитания ситуация постепенно начинает улучшаться вплоть до полного восстановления в результате радикальной перестройки нейронной активности спинальных двигательных центров.


Рис. 14. Упрощенная схема моторных центров спинного мозга интактного животного (1) и животного после перекрестного сшивания эфферентных нервов (2).

Рис. 13. Кора головного мозга играет главную роль в обеспечении пластических реакций ЦНС после повреждений.
В рассмотренном примере пластические перестройки в работе спинальных нервных центров происходят преимущественно за счет пластических перестроек нейронной активности на уровне коры головного мозга. Именно нисходящие корковые возбуждения нового качества вносят решающий вклад в изменение работы спинальных нервных центров. Ведущую роль коры головного мозга в обеспечении пластических перестроек спинальных центров можно доказать в эксперименте. Перережьте нисходящие в спинной мозг корковые двигательные тракты у животного после восстановления двигательных функций – ситуация немедленно вернется к исходной.
8. Нервные центры (НЦ) обладают повышенной утомляемостью. Известно, что нервное волокно практически не утомляется (Н. Е. Введенский), напротив, НЦ обладают повышенной утомляемостью. Неслучайно в целом организме утомление раньше всего развивается в НЦ (И. М. Сеченов).
9. Нервные центры обладают повышенной чувствительностью к ядам, биологически активным веществам, лекарственным препаратам. Это связано с обилием синаптических контактов в структуре нейронных сетей НЦ, наличием множества химических механизмов передачи возбуждения через синапсы.
10. Нервные центры обладают повышенной чувствительностью к недостатку кислорода.
11. Нервные центры (НЦ) могут пребывать в трех функциональных состояниях, отражающих совокупный уровень активности всех нейронов НЦ. Исходное состояние НЦ – нормальная (обычная) активность нейронов НЦ; второе состояние – сниженная активность нейронов НЦ; третье состояние – повышенная активность нейронов НЦ. В последнем случае НЦ в составе ЦНС становится доминантным, или господствующим (А. А. Ухтомский).
1. Гормональные причины, например, избыток или недостаток в циркулирующей крови тех или иных гормонов. 2. Патологические причины, например, хроническая болевая импульсация, формирующая доминантное состояние центров, ответственных за переработку болевой импульсации (ядра ретикулярной формации, гипоталамуса, таламуса, лимбической системы). 3. Метаболические причины, например, недостаток воды в организме, формирующий доминантное состояние гипоталамического центра жажды. 4. Информационные причины (применительно к человеку), связанные с восприятием словесных сигналов, письменных сообщений и проч., формирующих доминантное состояние корковых центров, имеющих первостепенное значение для организации целенаправленных поведенческих актов.
Читайте также:
