
Какой нервной системой иннервируются скелетные мышцы
Виды мышц:
-скелетные: произвольные, поперечно-полосатые
-сердечные: непроизвольные, поперечно-полосатые
-гладкие: непроизвольные, нет поперечной исчерченности
Типы мышечных волокон:
- медленные (красные) – много миоглобина ( связывает О2) и митохондрий; поддержание позы; утомление наступает очень медленно, быстро проходит.
- быстрые (окислительного типа) – много митохондрий; образует АТФ путём окислительного фосфорилирования; быстрые энергичные движения без заметного утомления.
- сокращение и расслабление происходят медленно; входят в состав наружных мышц глаза.
Иннервация скелетной мышцы:
Соматическая нервная система (произвольная); один двигательный нейрон (тело в ЦНС); осуществляет возбуждение. Двигательную (эфферентную) иннервацию скелетные мышцы туловища и конечностей получают от мотонейронов передних рогов спинного мозга, а мышцы лица и головы - от двигательных нейронов определенных черепных нервов. При этом к каждому мышечному волокну подходит или ответвление от аксона мотонейрона, или же весь аксон. В мышцах, обеспечивающих тонкие координированные движения (мышцы кистей, предплечий, шеи), каждое мышечное волокно иннервируется одним мотонейроном. В мышцах, обеспечивающих преимущественно поддержание позы, десятки и даже сотни мышечных волокон получают двигательную иннервацию от одного мотонейрона, посредством разветвления его аксона.
Двигательное нервное волокно, подойдя к мышечному волокну, проникает под эндомизий и базальную пластинку и распадается на терминали, которые вместе с прилежащим специфическим участком миосимпласта образуют аксо-мышечный синапс или моторную бляшку. Под влиянием нервного импульса волна деполяризации с нервного окончания передается на плазмолемму миосимпласта, распространяется далее по Т-канальцам и в области триад передается на терминальные цистерны саркоплазматической сети, обуславливая выход ионов кальция и начало процесса сокращения мышечного волокна.
Нейромоторная единица - совокупность нейрона и группы мышечных волокон, иннервируемых аксоном этого нейрона.
В состав нейромоторной единицы входят:
нервная клетка - в основном мотонейроны, тела которых лежат в передних рогах спинного мозга;
аксон мотонейрона - миелиновые волокна;
группа мышечных волокон - в зависимости от вида деятельности количества волокон различно. Если тонкая работа 2-4, если грубая - до нескольких тысяч.
Виды нейромоторных единиц
По характеру возбуждения, возникающего в мышечных волокнах все нейромоторные единицы делятся на 2 группы.
Фазные нейромоторные единицы- образуются альфа-мотонейронами. Это самые крупные мотонейроны (диаметр 10-20 мкм). Скорость проведения возбуждения по отростку этого аксона - 120 м/с. Аксон обеспечивает одиночную иннервацию, образуя на каждом волокне 1-2 синапса. В каждом синапсе при возбуждении возникает достаточный потенциал концевой пластинки, который обеспечивает возникновение потенциала действия в мышце (импульсное возбуждение). Лучше работают под действием одиночного возбуждения (импульса), при котором возникает распространяющееся возбуждение.
Среди фазных нейромоторных единиц выделяют быстрые и медленные.
Быстрые - продолжительность потенциала действия в 2 раза меньше, чем в медленных. Волна сокращения в 5 раз меньше, чем в медленных. Скорость распространения возбуждения в быстрых нейромоторных единицах в 2 раза больше, чем в медленных. Таким образом, быстрые фазные нейромоторные единицы обеспечивают динамическую работу, когда быстрое сокращение сменяется быстрым расслаблением. Для этого вида нейромоторных единиц характерно анаэробное образование энергии. Эти мышечные волокна практически не содержат миоглобина - светлые, белые мышцы;
Медленные- обеспечивают в основном статическую работу, медленное, длительное сокращение мышц. Основной поставщик энергии окислительно-восстановительные процессы. Содержит миоглобин, который депонирует кислород. По цвету темные, красные мышцы.
Тонические нейромоторные единицы- образуются гамма-мотонейронами передних рогов спинного мозга (самые мелкие, диаметр 4-6 мкм). Скорость проведения импульса по отросткам этих нейронов - 30 м/с, в синапсах возникает местный потенциал концевой пластинки (местное возбуждение). Аксоны тонических нейронов обеспечивают множественную иннервацию, т. е. на каждом мышечном волокне образуются несколько десятков синапсов и за счет суммации местных потенциалов и возникает потенциал действия. Возбуждение тонической нейромоторной единицы происходит под влиянием серии импульсов (частота около 10 Гц). Тонические нейромоторные единицы обеспечивают медленное сокращение мышц, участвуют в возникновении тонуса.
Скелетные мышцы образуются различными мышечными волокнами, которые входят в состав фазных и тонических нейромоторных единиц. Попеременное включение нейромоторных единиц обеспечивает изменение функционального состояния мышцы. В состоянии покоя работают тонические нейромоторные единицы, в состоянии активности - фазные нейромоторные единицы.
Кроме двигательной иннервации присутствует вегетативная. Все скелетные мышцы получают импульсы из симпатической нервной системы, которая регулирует обменные процессы.
Мышечные веретена состоят из интрафузальных мышечных волокон, количество которых не превышает 12. (В данном контексте экстрафузалъные мышечные волокна считают обычными). Крупные интрафузальные волокна начинаются от полюсов (концов) мышечных веретен и фиксируются к соединительной ткани — перимизию. Некрупные интрафузальные волокна фиксируются к коллагеновой капсуле веретена. В средней части (экваторе) веретена саркомеры практически полностью замещены ядрами, расположенными в форме сумки—сумчато-ядерные мышечные волокна (характерно для широких волокон) или цепочки — цепочечно-ядерные мышечные волокна (характерно для тонких волокон).
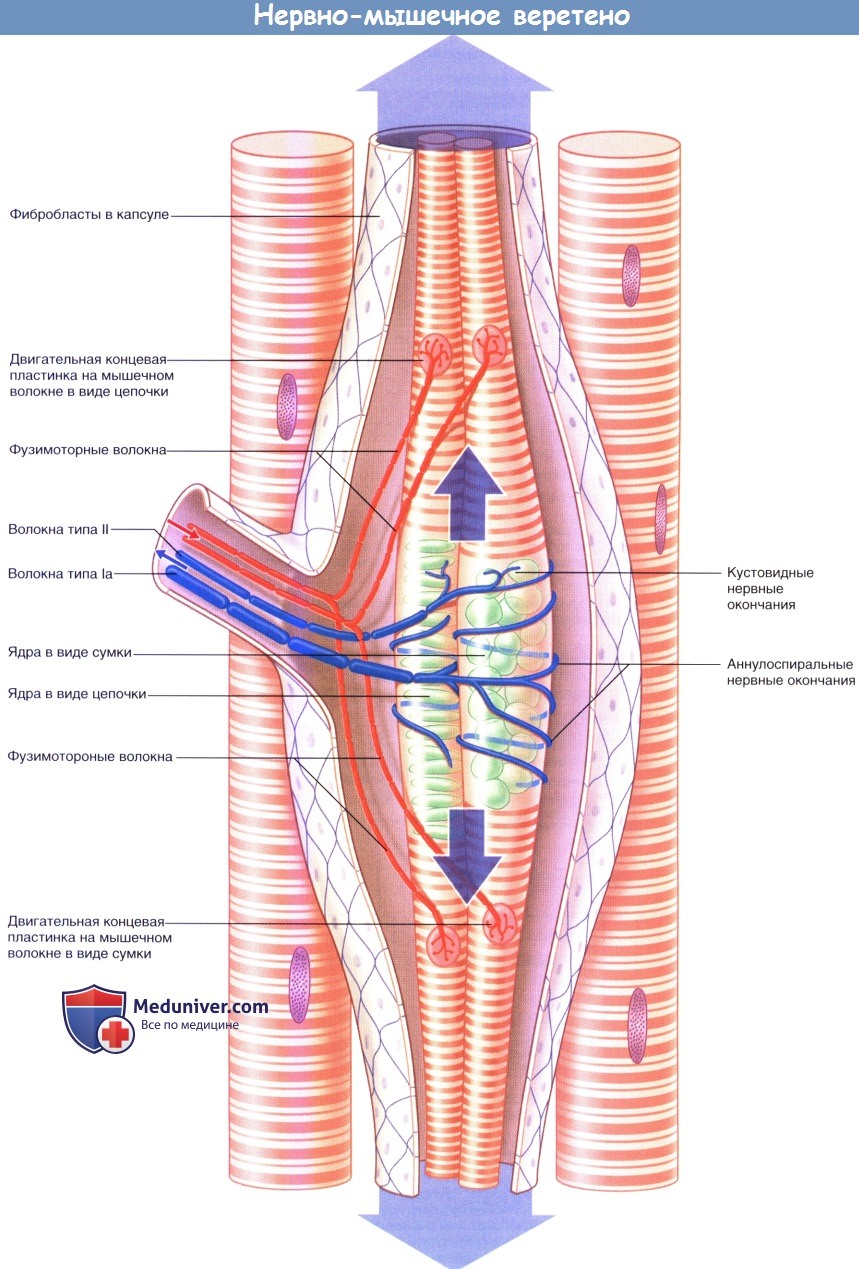
Упрощенное строение нервно-мышечного веретена. Большими стрелками обозначено пассивное растяжение аннулоспиральных нервных окончаний при удлинении мышцы в состоянии покоя.
Стрелками среднего размера обозначено активное растяжение аннулоспиральных нервных окончаний фузимоторными нервными волокнами.
Активное растяжение в достаточной степени компенсирует эффект разгрузки одновременного сокращения экстрафузальных мышечных волокон.
Стрелками маленького размера показаны направления проведения импульсов к мышечному веретену и от него при сокращении мышцы.
1. Иннервация. Иннервацию мышечных веретен осуществляют двигательные и чувствительные нервные волокна. Двигательные нервные волокна — фузимоторные—в соответствии с толщиной относят к типу Аγ, а нервные волокна экстрафузальных мышц — к типу Аα. Фузимоторные аксоны разветвляются и иннервируют поперечно-исчерченные сегменты концевых участков интрафузальных мышц. Единичные первичные чувствительные нервные волокна типа 1а оборачиваются спирально-кольцевым образом вокруг сумчато-ядерных или цепочечно-ядерных мышечных волокон. Вторичные кустовидные чувствительные нервные окончания образованы нервными волокнами II типа и располагаются с одной или с двух сторон от первичного волокна.
2. Активация. Мышечные веретена представляют собой рецепторы растяжения. При растяжении мышечного веретена ионные каналы поверхностной мембраны чувствительных нервных окончаний открываются, за счет чего создаются положительные электроволны. В результате их суммирования в области терминального участка аксона чувствительного нервного волокна происходит образование рецепторного потенциала, который при достижении пороговых значений генерирует нервные импульсы.
Растяжение мышечных веретен может происходить пассивным и активным путями.
3. Пассивное растяжение. Пассивное растяжение мышечного веретена происходит при пассивном удлинении всего мышечного брюшка. Например, при воспроизведении сухожильных рефлексов, в частности коленного, во время удара по сухожилию происходит пассивное растяжение мышечных веретен брюшка четырехглавой мышцы бедра. Волокна типов Iа и II направляются к спинному мозгу и образуют синапсы с дендритами α-мотонейронов. (Принадлежность мотонейронов к типу а определяется Аα-диаметром их аксонов.)
Сухожильные рефлексы относят к моносинаптическим. Для них характерен латентный период (промежуток времени до ответной реакции на стимул), составляющий 15-25 мс.
Помимо возбуждающего действия на гомонимные мотонейроны (т. е. двигательные нейроны, иннервирующие одни и те же мышцы), афферентные волокна мышечных веретен опосредованно через вставочные тормозные мотонейроны оказывают тормозное действие на а-мотонейроны мышц-антагонистов. Этот процесс получил название реципрокного торможения. Задействованные тормозные нейроны называют вставочными нейронами типа Iа.
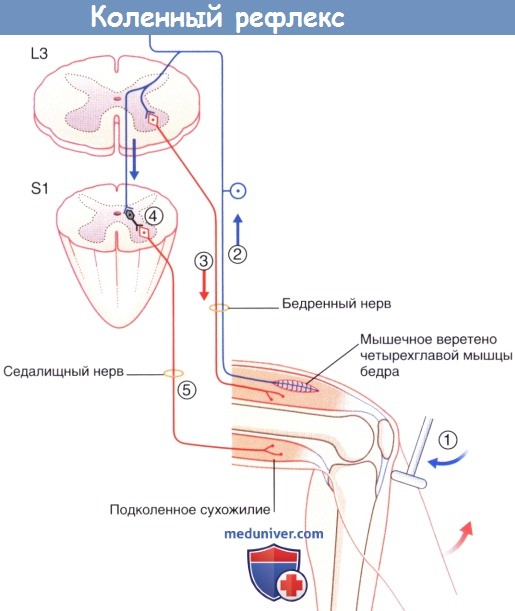
Коленный рефлекс, в котором задействовано реципрокное торможение.
Стрелочками указано проведение нервных импульсов.
(1) Удар по коленной связке вызывает растяжение мышечных веретен в четырёхглавой мышце спины.
(2) Мышечные веретена посылают возбуждающий потенциал в спинной мозг.
(3) α-Мотонейроны вызывают подергивание четырехглавой мышцы и разгибание колена.
(4 и 5) Тормозные вставочные нейроны типа Ia полностью подавляют активность подколенного сухожилия.
4. Кодирование информации. Первичные афференты мышечных веретен проявляют наибольшую активность во время растяжения. Они определяют скорость растяжения мышцы: чем быстрее происходит растяжение, тем больше импульсов они генерируют.
Вторичные афференты мышечных веретен по сравнению с первичными активнее задействованы в процессе поддержания определенной позиции и определяют степень растяжения мышцы: чем выше степень статического растяжения, тем больше формируется импульсов.
б) Нервные окончания сухожилий. Сухожильные органы Гольджи располагаются в местах соединения сухожилий с мышцами. Единичное нервное волокно типа Ib распадается на сложноорганизованные мельчайшие разветвления, оплетающие пучки сухожильных волокон, связанные с соединительно-тканной капсулой.
Десятки мышечных волокон погружаются в сухожильные интракапсулярные волокна, которые последовательно связываются с другими мышечными волокнами в пределах определенной мышцы. Под действием натяжения, возникающего в процессе сокращения мышцы, происходит активация луковицеобразных нервных окончаний. В связи с тем, что скорость возникновения импульсов в исходном нервном волокне зависит от силы натяжения, сухожильные нервные окончания определяют силу сокращения мышцы.
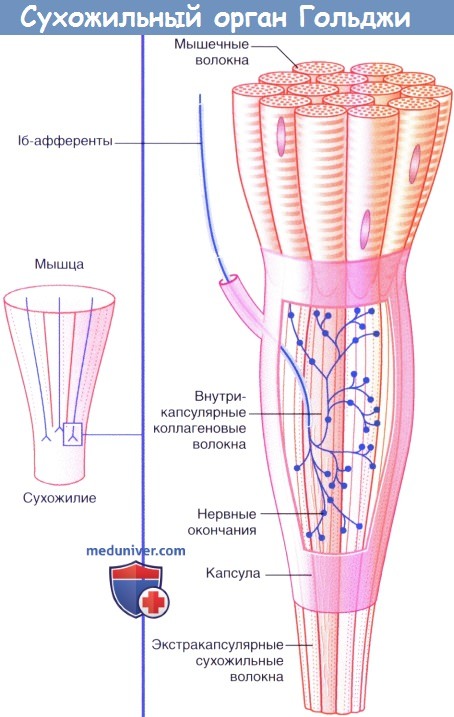
Сухожильный орган Гольджи. 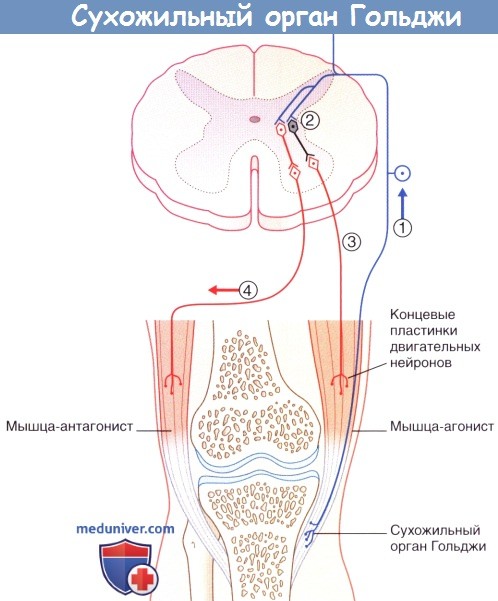
Рефлекторные эффекты при стимуляции сухожильного органа Гольджи.
(1) Сокращение мышцы-агониста вызывает возбуждение афферентов сухожильного органа Гольджи,
что приводит к (2) возбуждению тормозных вставочных нейронов, образующих синапсы с гомонимными мотонейронами,
а также (3) возбуждению тормозных вставочных нейронов, образующих синапсы с (4) мотонейронами мышцы-антагониста.
в) Свободные нервные окончания. В мышцах имеется множество свободных нервных окончаний, большинство которых расположено во внутримышечной соединительной ткани и покрывающих мышцы фасциях. Эти нервные окончания отвечают за болевую чувствительность, возникающую при непосредственном повреждающем воздействии или при накоплении продуктов распада, к которым относят молочную кислоту.
г) Иннервация суставов. Немиелинизированные нервные волокна со свободными нервными окончаниями в большом количестве присутствуют в связках и капсулах суставов, а также во внешних частях внутрисуставных менисков. Эти нервные волокна обеспечивают болевую чувствительность при деформациях суставов, а также участвуют в формировании защитного рефлекса для капсулы сустава. Так, например, переднюю капсулу запястья иннервируют срединный и локтевой нервы; внезапное растяжение при насильственном разгибании вызывает рефлекторную активацию двигательных волокон, что приводит к сгибанию верхней конечности в лучезапястном суставе.
В экспериментах на животных показано, что при воспалении сустава в возбужденном состоянии находится большее количество нервных волокон, чем при растяжении капсулы здорового сустава. Вероятно, существуют нервные окончания, единственным стимулом которых служит воспалительный процесс.
Инкапсулированные нервные окончания, расположенные внутри суставных капсул и вокруг них, представлены тельцами Руффини, реагирующими на натяжение, ламеллярными окончаниями, воспринимающими давление, а также тельцами Пачини, ответственными за ощущение вибрации.
д) Миофасциальный болевой синдром. Миофасциальный болевой синдром—распространенное заболевание, проявляющееся региональной мышечной болезненностью, связанной с чрезмерной чувствительностью пучков напряженных мышечных волокон. (Сходные синдромы наблюдают при другом заболевании—фибромиалгии. Однако при фибромиалгии происходит центральное нарушение болевой чувствительности—дисфункция системы модуляции боли.)
Прикосновение к пучкам мышечных волокон с повышенной чувствительностью вызывает боль; клинически эту область называют миофасциальной триггерной точкой. Для боли не характерно распространение в области дерматома какого-то определенного нерва; в некоторых случаях боль может выходить за пределы триггерной точки—иррадиировать. Кроме того, боль могут сопровождать вегетативные проявления, такие как покраснение и пилоэрекция. Возникновение триггерных точек может быть связано с травмой мышц, чрезмерными нагрузками во время профессиональной деятельности или спортивных занятий при нарушении процесса нормального восстановления.
Спонтанно активирующиеся очаги получили название активных миофасциальных триггерных точек (МТТ), а находящиеся в данный момент времени в неактивном состоянии—латентных миофасциальных триггерных точек. Происходящие процессы недостаточно ясны с точки зрения патофизиологии, однако установлено, что тканевая жидкость, окружающая активные МТТ, содержит большее количество ассоциированных с воспалением веществ (например, брадикинины, простагландины, протоны Н + ).
Со временем боль может захватывать новые области или усиливаться в результате раздражения нейронов задних рогов. Высвобождение другими ветвями раздраженных нейронов субстанции Р может привести к возникновению новых МТТ в области той же или прилежащей мышцы.
Стойкое сокращение мышечных волокон, прилежащих к узелковым утолщениям, объясняют инактивацией ацетилхолинэстеразы в базальной мембране их концевых двигательных пластинок. Принципы лечения этого заболевания—длительное пассивное растяжение пораженных мышц, длительное давление в горизонтальном положении больного (например, путем подкладывания теннисного мячика под пораженную область), а также механическое повреждение прокалыванием иглой или введением местных анестетиков и/или стероидов.
е) Резюме. Мышцы. Двигательная единица состоит из двигательного нейрона и иннервируемой им группы мышечных волокон. Каждая двигательная единица содержит определенный гистохимический вид мышечных волокон. Концевое утолщение нервно-мышечного синапса, содержащее пузырьки с АХ, отделено от синаптических складок сарколеммы базальной мембраной, в которой присутствует ацетилхолинэстераза.
В состав мышечных веретен входят интрафузальные мышечные волокна, активация которых происходит с обоих концов за счет γ-фузимоторных нейронов. Чувствительные волокна типа Iа образуют первичные аннулоспиральные нервные окончания в области средней части (экватора), а волокна типа II — вторичные нервные окончания. Оба типа волокон являются рецепторами растяжения. Растяжение может происходить пассивным (например, при сухожильном рефлексе) или активным путем во время фузимоторной активности. Гомонимные мотонейроны—моносинаптические; мышцы-антагонисты реципрокно ингибируются посредством вставочных нейронов Iа. Первичные афференты мышечных веретен определяют скорость мышечного сокращения, а вторичные — степень.
В процессе произвольных движений происходит одновременная активация А α- и Аγ-мотонейронов.
Сухожильные органы Гольджи определяют силу мышечных сокращений. В состав сухожильных органов входит инкапсулированное сухожилие, иннервацию которого осуществляют афференты типа Iб, вызывающие двухсинаптическое торможение гомонимных мотонейронов и реципрокное возбуждение мышц-анта-гонистов.
Свободные внутримышечные нервные окончания обеспечивают болевую чувствительность.
Суставы. Свободные нервные окончания в большом количестве присутствуют в связках и капсулах суставов, а также во внешних частях внутрисуставных менисков. Они обеспечивают болевую чувствительность и формирование суставного защитного рефлекса. Инкапсулированные нервные окончания принимают импульсы от движений суставов.
- Вернуться в оглавление раздела "Неврология."
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Рассмотрена иннервация скелетной мышцы. Описан состав и строение нейрона, а также функции нейрона. Рассмотрены функции двигательных, чувствительных и вегетативных нейронов. Описан состав периферического нерва. Введено понятие двигательной единицы и концевой пластинки (синапса).
Иннервация скелетной мышцы
Иннервация (от in — в, внутри и nervus — нервы) — снабжение органов и тканей нервами, что обеспечивает их связь с центральной нервной системой (ЦНС).
Иннервация скелетной мышцы — это наличие нервных волокон, которые передают импульсы из ЦНС к мышце и от мышцы в ЦНС.
Денервация скелетной мышцы — нарушение передачи импульсов из ЦНС к мышце или от мышцы в ЦНС. Денервация мышечных волокон возможна из-за перерезки или повреждения нерва. Денервация мышечных волокон происходит также по мере старения из-за уменьшения количества нервных волокон, иннервирующих скелетную мышцу.
Теперь рассмотрим этот вопрос подробнее. Давайте сначала разберемся, что представляет собой нейрон.
Нейрон – это структурная единица нашей нервной системы, главная функция которого – передача информации от одного участка тела другому. Чтобы передать информацию нейрон возбуждается, затем вырабатывает нервный импульс. Нейроны также участвуют в обработке и хранении информации. Но этого вопроса мы касаться не будем.
Нейрон – это высокоспециализированная клетка. Он состоит из тела и длинного отростка – аксона (рис. 1.). 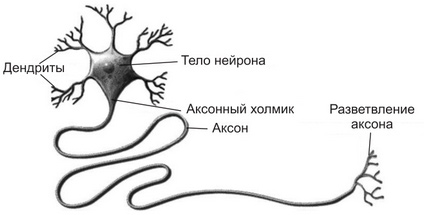
Длина аксона может достигать одного метра и более. На теле есть много мелких отростков – дендритов. Через эти отростки нейрон получает информацию от других нейронов и передает ее через аксон или другому нейрону, или органу, к которому он подходит (в нашем случае – мышце).
Скелетные мышцы иннервируются одним или несколькими периферическими нервами. За несколькими исключениями (лицевой и подъязычный нерв) все периферические нервы являются смешанными. Смешанный периферический нерв в своем составе содержит:
- двигательные волокна (нейроны);
- чувствительные волокна (нейроны);
- вегетативные волокна (нейроны).
Когда мы рассматривали строение скелетной мышцы, то указывалось, что периферические нервы имеют каналы в перимизии. Через эти каналы периферические нервы подходят к мышечным волокнам.
Двигательные нейроны (мотонейроны) несут информацию от ЦНС к мышце. Тела мотонейронов расположены в передних рогах спинного мозга. Аксоны мотонейронов идут к мышцам в составе периферического нерва.
Двигательные нейроны делятся на два типа: α-мотонейроны (альфа-мотонейроны) и γ-мотонейроны (гамма-мотонейроны).
Один α-мотонейрон иннервирует (то есть пускает веточки) достаточно много мышечных волокон. Иногда на один аксон приходится более 2000 мышечных волокон. Такая система, состоящая из одного нейрона и мышечных волокон, которые он иннервирует, называется двигательной единицей (ДЕ). Это понятие ввел в физиологию нобелевский лауреат Чарльз Скотт Шеррингтон в начале XX века. Особенности состава и функционирования ДЕ мы рассмотрим позднее.
Место, где аксон α-мотонейрона соединяется с мышечным волокном называется концевой пластинкой (синапсом). Через синапс к мышечному волокну из ЦНС (центральной нервной системы) поступают сигналы, которые вызывают его возбуждение.
Чувствительные волокна несут в ЦНС информацию о различных показателях активности мышцы (длине мышцы, скорости ее сокращения, степени напряжения). Если бы ЦНС не могла получать эту информацию, управление напряжением мышцы было бы невозможно. Точно так же было бы невозможно управление нашими движениями. Тела чувствительных нейронов расположены в задних рогах спинного мозга.
Чувствительные нейроны либо заканчиваются свободными нервными окончаниями, либо иннервируют рецепторы скелетных мышц (мышечные веретена и рецепторы сухожилий).
- Свободные нервные окончания называются ноцицепторами. Они расположены между мышечными волокнами и несут в ЦНС информацию о боли.
- От мышечных веретен отходят чувствительные нейроны (Ia-афференты), несущие в ЦНС информацию о длине и скорости сокращения скелетной мышцы.
- От сухожильных рецепторов отходят чувствительные нейроны (Ib-афференты), несущие в ЦНС информацию о напряжении скелетной мышцы.
Вегетативные нейроны иннервируют гладкие мышцы стенок кровеносных сосудов скелетных мышц.
Иннервация скелетных мышц пожилых людей ухудшается. Ученые находят, что с возрастом уменьшается количество α-мотонейронов, иннервирующих скелетную мышцу. Это является одним из факторов, обусловливающих возрастное уменьшение массы скелетных мышц и их силы — саркопению.
- Ванек, Ю. Спортивная анатомия.– М.: Академия, 2008.- 304 с.
- Мак-Комас, А.Дж. Скелетные мышцы.– Киев: Олимпийская литература, 2001.– 407 с.
- Мышцы в спорте. Анатомия. Физиология. Тренировка. Реабилитация. — М.: Практическая медицина, 2016.- 408 с. (отличная книга, рекомендую!).
- Самсонова, А.В. Гипертрофия скелетных мышц человека. – СПб: Кинетика, 2018. – 159 с.
Двигательная иннервация мышечных волокон скелетных мышц
- Двигательная иннервация мышечных волокон скелетных мышц
- Нервно-мышечный синапс
- Потенциал действия
- Все страницы
Двигательная и чувствительная соматическая иннервация мышечных волокон скелетных мышц осуществляется соответственно альфа- и гамма?мотонейронами передних рогов спинного мозга и двигательных ядер черепных нервов и псевдоуниполярными чувствительными нейронами спинномозговых узлов и чувствительных ядер черепных нервов.
Вегетативная иннервация мышечных волокон в скелетных мышцах не обнаружена, но ГМК стенки кровеносных сосудов мышц имеют симпатическую адренергическую иннервацию.
Двигательная иннервация
Каждое экстрафузальное мышечное волокно [1] имеет прямую двигательную иннервацию — нервно-мышечные синапсы, образованные терминальными ветвлениями аксонов альфа?мотонейронов и специализированными участками плазмолеммы мышечного волокна (концевая пластинка, постсинаптическая мембрана).
Экстрафузальные мышечные волокна входят в состав нейромоторных (двигательных) единиц и обеспечивают сократительную функцию мышц.
Интрафузальные мышечные волокна [2] образуют нервно-мышечные синапсы с эфферентными волокнами гамма?мотонейронов.
Рис. 7–6. Двигательная единица
Двигательная единица (рис. 7–6) включает один мотонейрон и группу иннервируемых им экстрафузальных мышечных волокон. Количество и размеры двигательных единиц в различных мышцах значительно варьируют.
Каждая двигательная единица образована только быстросокращающимися или только медленносокращающимися мышечными волокнами (см. ниже).
Формирование двигательных единиц происходит в постнатальном периоде, а до рождения каждое мышечное волокно иннервируется несколькими мотонейронами. Аналогичная ситуация возникает при денервации мышцы (например, при повреждении нерва) с последующей реиннервацией мышечных волокон. Понятно, что в этих ситуациях страдает эффективность сократительной функции мышцы.
Физиология нервно-мышечных синапсов рассмотрена в главах 4 (см. рис. 4–8) и 6 (см. рис. 6–2 в статье Синапсы и 6–3 в статье Организация и функция синапса).
Как и любой синапс, нервно-мышечное соединение состоит из трех частей: пресинаптической области, постсинаптической области и синаптической щели.
Двигательная нервная терминаль нервно-мышечного синапса снаружи покрыта шванновской клеткой [3], имеет диаметр 1–1,5 мкм и образует пресинаптическую область нервно-мышечного синапса. В пресинаптической области в большом количестве присутствуют синаптические пузырьки, заполненные ацетилхолином (5–15 тыс. молекул в одном пузырьке) и имеющие диаметр порядка 50 нм.
На постсинаптической мембране — специализированной части плазмолеммы мышечного волокна — имеются многочисленные инвагинации, от которых на глубину 0,5–1,0 мкм отходят постсинаптические складки, чем существенно увеличивается площадь мембраны. В постсинаптическую мембрану встроены н?холинорецепторы, их концентрация достигает 20–30 тысяч на 1 мкм 2 .
Постсинаптические н?холинорецепторы (рис. 7–7) Диаметр открытого канала в составе рецептора равен 0,65 нм, что вполне достаточно для свободного прохождения всех необходимых катионов: Na+, K+, Ca2+. Отрицательные ионы, такие как Cl–, не проходят через канал вследствие сильного отрицательного заряда в устье канала.
Рис. 7–7. Никотиновый холинорецептор постсинаптической мембраны. А — рецептор не активирован, ионный канал закрыт. Б — после связывания рецептора с ацетилхолином канал кратковременно открывается. Реально через канал проходят преимущественно ионы Na+ в силу следующих обстоятельств: — в среде, окружающей рецептор ацетилхолина, в достаточно больших концентрациях имеется лишь два положительно заряженных иона: во внеклеточной жидкости Na+ и во внутриклеточной жидкости K+; — сильный отрицательный заряд внутренней поверхности мышечной мембраны (от –80 до –90 мВ) притягивает положительно заряженные ионы натрия внутрь МВ, одновременно предотвращая попытки ионов калия двигаться наружу.
Внесинаптические холинорецепторы. Холинорецепторы присутствуют также в мембране мышечного волокна вне синапса, но здесь их концентрация на порядок величины меньше, чем в постсинаптической мембране.
Через синаптическую щель проходит синаптическая базальная мембрана. Она удерживает в области синапса терминаль аксона, контролирует расположение холинорецепторов в виде скоплений в постсинаптической мембране. В синаптической щели также находится фермент ацетилхолинэстераза, расщепляющий ацетилхолин на холин и уксусную кислоту.
Нервно-мышечная передача возбуждения состоит из нескольких этапов.
- ПД по аксону достигает области двигательного нервного окончания.
- Деполяризация мембраны нервного окончания приводит к открытию потенциалозависимых Са2+?каналов и входу Са2+ в двигательное нервное окончание.
- Повышение концентрации Са2+ приводит к запуску экзоцитоза квантов ацетилхолина из синаптических пузырьков.
- Ацетилхолин попадает в синаптическую щель, где путём диффузии достигает рецепторов на постсинаптической мембране. В нервно-мышечном синапсе в ответ на один ПД выделяется около 100–150 квантов ацетилхолина.
- Активация н?холинорецепторов постсинаптической мембраны. При открытии каналов н?холинорецепторов возникает входящий Na–ток, что приводит к деполяризации постсинаптической мембраны. Появляется потенциал концевой пластинки, который при достижении критического уровня деполяризации вызывает ПД в мышечном волокне.
- Ацетилхолинэстераза расщепляет ацетилхолин и действие выделившийся порции нейромедиатора на постсинаптическую мембрану прекращается.
В физиологических условиях каждый нервный импульс, поступающий в нервно-мышечное соединение, вызывает возникновение потенциала концевой пластинки, амплитуда которого в три раза больше необходимой для возникновения ПД. Появление такого потенциала связано с избыточностью выделения медиатора. Под избыточностью подразумевается выделение в синаптическую щель значительно большего количества ацетилхолина, чем требуется для запуска ПД на постсинаптической мембране. Этим гарантируется, что каждый ПД мотонейрона вызовет реакцию в иннервируемом им МВ.
Холиномиметики. Метахолин, карбахол и никотин оказывают на мышцу тот же эффект, что и ацетилхолин. Различие заключается в том, что эти вещества не разрушаются ацетилхолинэстеразой или разрушаются более медленно, в течение многих минут и даже часов.
Антихолинэстеразные соединения. Неостигмин, физостигмин и диизопропилфлуорофосфат инактивируют фермент таким образом, что имеющаяся в синапсе ацетилхолинэстераза теряет способность гидролизовать ацетилхолин, выделяющийся в концевой двигательной пластинке. В результате происходит накопление ацетилхолина, что в ряде случаев может вызывать мышечный спазм. Это может приводить к смертельным исходам при спазме гортани у курильщиков. Неостигмин и физостигмин инактивируют ацетилхолинэстеразу в течение нескольких часов, после чего их действие проходит, и синаптическая ацетилхолинэстераза восстанавливает свою активность. Диизопропилфлуорофосфат, являющийся нервно-паралитическим газом, блокирует ацетилхолинэстеразу на недели, что делает это вещество смертельно опасным.
- Миорелаксанты периферического действия (кураре и курареподобные препараты) широко применяются в анестезиологии. Тубокурарин препятствует деполяризующему действию ацетилхолина. Дитилин приводит к миопаралитическому эффекту, вызывая стойкую деполяризацию постсинаптической мембраны.
- Ботулотоксин и столбнячный токсин блокируют секрецию медиатора из нервных терминалей.
- бетта- и гамма-Бунгаротоксины блокируют холинорецепторы.
- Миастения тяжёлая псевдопаралитическая (myasthenia gravis) — аутоиммунное заболевание, при котором образуются АТ к н?холинорецепторам. Циркулирующие в крови АТ связываются с н?холинорецепторами постсинаптической мембраны МВ, препятствуют взаимодействию холинорецепторов с ацетилхолином и угнетают их функцию, что приводит к нарушению синаптической передачи и развитию мышечной слабости. Ряд форм миастений вызывает появление АТ к кальциевым каналам нервных окончаний в нервно-мышечном соединении.
- Денервация мышцы. При двигательной денервации происходит значительное увеличение чувствительности мышечных волокон к эффектам ацетилхолина вследствие увеличенного синтеза рецепторов ацетилхолина и их встраивания в плазмолемму по всей поверхности мышечного волокна.
Потенциал действия мышечного волокна
Природа и механизм возникновения потенциала действия рассмотрены в главе 5. ПД МВ длится 1–5 мс, скорость его проведения по сарколемме, включая T-трубочки, составляет 3–5 м/с.
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология : Учебник. - М.: ГЭОТАР-Медиа, 2009. – 688 с. Глава 7. Физиология сократительных элементов. - Скелетная мышца. - Иннервация. - Двигательная иннервация. - С. 101-102.
[1] Экстрафузальное мышечное волокно (истор.; лат. extra вне, снаружи + гист. fusus веретено) - общее название поперечнополосатых мышечных волокон, составляющих основную массу скелетных мышц и обладающих способностью к сокращению на всем протяжении. (Словарь медицинских терминов)/
[ 2] Интрафузальное мышечное волокно (син. волокно мышечное внутриверетенное - myofibra intrafusalis, LNH) мышечное волокно, входящее в состав нервно-мышечного веретена; периферические части интрафузального мышечного волокна обладают поперечной исчерченностью и способны к сокращению, центральная часть не сокращается.
[3] Шванн Теодор (Schwann T.), немецкий гистолог и физиолог (1810–1882); вместе с М. Шульце создал клеточную теорию (1839 г.); в 1836 г. открыл пепсин, в 1838 г. опубликовал первую работу по строению миелиновой оболочки.
Читайте также:
