
Надпочечники как симпатический ганглий
Подобно задней доле гипофиза, мозговой слой надпочечников представляет собой производное нервной ткани. Его можно рассматривать как специализированный симпатический ганглий. Скопления хромаффинной ткани обнаруживают в симпатической нервной системе (параганглиях). Цепочка хромаффинных телец расположена кпереди от брюшной аорты, в области бифуркации аорты; каротидные тельца также составляют часть хромаффинной системы организма.
Хромаффинные клетки надпочечников секретируют в основном адреналин и в меньшей степени норадреналин, тогда как постганглионарные клетки симпатической нервной системы — преимущественно норадреналин.
Сходство продуктов и способов реагирования симпатической нервной системы и мозгового слоя надпочечников стало основанием для объединения этих структур в единую симпатоадреналовую систему с выделением нервного и гормонального её звена.
Хромаффинные клетки и клетки симпатических ганглиев образуются в эмбриогенезе из клеток зародышевого нервного гребешка, называемых симпатогониями. Эти клетки служат предшественниками симпатобластов (из которых развиваются клетки симпатических ганглиев) и феохромоблас-тов (дающих начало хромаффинным клеткам). Из хромаффинных клеток может развиться феохромоцитома. Из других клеток нервного гребешка возникают другие типы катехоламин-продуцирующих опухолей:
■ из симпатобластов — симпатобластома;
■ из феохромобластов — феохромобластома;
■ из клеток симпатического ганглия — ганглионейрома.
Опухоли первых двух видов называют нейробластомами, третьего — ган-глионейромой (ганглионейробластома). Эти типы опухолей наблюдают у новорождённых и детей раннего возраста, и очень редко — у взрослых. Нейробластомы чаще обнаруживают в возрасте 1-3 лет, это очень злокачественные опухоли. Не менее 50% этих опухолей локализуется в брюшной полости (до 35% — в надпочечниках). Ганглионейробластома образуется путём трансформации клеток нейробластомы, содержит как нейробласты, так и ганглиозные клетки на разных стадиях дифференцировки. Ганглио-неврома — доброкачественная опухоль, состоящая из зрелых ганглиозных клеток.
У взрослых наиболее часто наблюдают феохромоцитому, образующуюся из хромаффинных клеток. В 90% случаев продуцирующая катехоламины опухоль хромаффинной ткани локализуется в мозговом веществе надпочечников, а в 10% — вне этих желёз. Злокачественными бывают менее 10% феохромоцитом.
При хромаффинных опухолях надпочечниковой и вненадпочечниковой локализации в кровоток поступает большое количество адреналина и но-радреналина. Это обусловливает возникновение гипертонических кризов на фоне нормального артериального давления (пароксизмальная форма заболевания), стойко повышенного артериального давления и периодически повторяющегося на этом фоне ещё большего подъёма давления (смешанная форма); стойкой артериальной гипертензии без кризов (постоянная форма).
Для оценки функционального состояния симпатоадреналовой системы используют целый спектр показателей. Однако в клинической практике применяют только часть из этих маркёров, главным образом для диагностики феохромоцитомы. Приблизительно у 1 из 200 пациентов с повышенным артериальным давлением выявляют феохромоцитому.
Катехоламины. Все три катехоламина естественного происхождения — норадреналин, адреналин и дофамин — функционируют как нейромедиаторы в центральной нервной системе. Норадреналин — нейромедиатор постганглионарных симпатических нервных окончаний, действует локально, в непосредственной близости от мест его высвобождения. Адреналин — циркулирующий в крови гормон мозгового вещества надпочечников — влияет на процессы, происходящие по всему организму. Существует также периферическая дофаминергическая система, но она не охарактеризована детально.
Рис. 66-1. Биосинтез, высвобождение и метаболизм катехоламинов. В верхней части рисунка схематически изображено периферическое симптоматическое нервное окончание; области луковицеобразных телец на терминальном волокне представлены расширениями, которые выявляют методом гистохимической флюоресценции как области высокой концентрации нейромедиаторов. В нижней части рисунка показаны хромаффинные клетки мозгового вещества надпочечников. Процессы биосинтеза, высвобождения, модуляции и обратного захвата показаны в последовательном порядке только в целях демонстрации: в организме все эти процессы протекают одновременно. ТГ — тирозингидроксилаза; ААД — декарбоксилаза ароматической L-аминокислоты;
ДА — дофамин; ДБГ — дофамин-р-гидроксилаза; НА — норадреналин; ФММТ — фенилэтаноламин-М-метилтрансфераза; А — адреналин; КОМТ — катехол-О-метилтрансфераза; НМН — норметанефрин; МАО— моноаминоксидаза; ДГМК — 3,4-дигидроксиманделиковая кислота; МГМК—З-метокси-4-гидроксиманделиковая кислота; АХ—ацетилхолин; Тир — тирозин; МН — метанефрин.
Биосинтез катехоламинов (рис. 66-1). Катехоламины синтезируются из аминокислоты тирозина, который последовательным гидрооксилированием преобразуется в диоксифенилаланин (дофа), затем декарбоксилируется, превращаясь в дофамин и гидроксилируется в b-положении боковой цепи, образуя норадреналин. Начальный этап этого процесса —- гидроксилирование тирозина — представляет собой этап, ограничивающий скорость всего процесса; он регулируется так, чтобы синтез диоксифенилаланина был связан с высвобождением норадреналина. Такая регуляция достигается изменениями как активности, так и количества тирозингидроксилазы. В мозговом веществе надпочечников и в тех центральных нейронах, в которых адреналин используется в качестве нейромедиатора, норадреналин N-метилируется в адреналин ферментом фенилэтаноламин-N-метилтрансферазой (ФNMT). Основная часть крови, снабжающей мозговое вещество надпочечников, обогащена кортикостероидами из коры надпочечников, и, поскольку надпочечниковая ФNМТ индуцируется глюкокортикоидами, способность мозгового вещества надпочечников образовывать адреналин может быть связана с ее стратегическим расположением внутри коры надпочечников.
Метаболизм катехоламинов (см. рис. 66-1). Основные метаболические трансформации катехоламинов включают в себя O-метилирование в метагидроксильной группе и окислительное дезаминирование. Процесс O-метилирования катализируется ферментом катехол-O-метилтрансферазой (КОМТ), а окислительному дезаминированию способствует моноаминоксидаза (МАО). КОМТ в печени и почках играет важную роль в метаболизме циркулирующих в крови катехоламинов. МАО, представляющая собой митохондриальный фермент, присутствующий в большинстве тканей, включая нервные окончания, менее значима в метаболизме циркулирующих в крови катехоламинов, но является важным фактором в регуляции их запасов в окончаниях периферических симпатических нервов. Метанефрины и 3-метокси-4-гидроксиманделиковая кислота (МГМК) являются главными конечными продуктами метаболизма и адреналина и норадреналина. Гомованилиновая кислота (ГВК) является конечным продуктом метаболизма дофамина.
Запасы катехоламинов и их высвобождение. Как в мозговом веществе надпочечников, так и в симпатических нервных окончаниях катехоламины сохраняются в субцеллюлярных гранулах и высвобождаются путем экзоцитоза. Большие запасы катехоламинов в этих тканях обеспечивают важный физиологический резерв, который поддерживает адекватное снабжение катехоламинами во время интенсивной стимуляции.
Мозговое вещество надпочечников. Хромаффинная ткань мозгового вещества обоих надпочечников здорового человека имеет массу около 1 г и содержит приблизительно 6 мг катехоламинов, 85% из которых приходится на долю адреналина. Высокая концентрация катехоламинов в хромаффинных гранулах поддерживается активным процессов их поступления, в котором участвуют мембраны гранул, и внутригранулярным комплексом хранения, в функционировании которого, по-видимому, участвуют АТФ, кальций и специфический белок гранул (хромогранин А). Секреция катехоламинов, стимулируемая ацетилхолином, высвобождающимся из преганглионарных симпатических нервов, возникает после попадания внутрь гранулы кальция, стимулирующего растворение мембраны хромаффинной гранулы и мембраны клетки; облитерация клеточной мембраны в месте ее растворения и вытеснение всего растворенного содержимого гранулы во внеклеточное пространство завершают процесс экзоцитоза (см. рис. 66-1). Ежесуточно происходит обмен приблизительно 2—10% общего запаса катехоламинов, содержащихся в мозговом веществе надпочечников.
Периферические симпатические нервные окончания. Периферические симпатические нервные окончания образуют сеть или сплетение, которое обеспечивает тесный контакт терминальных волокон с клетками, эффекторного органа. Весь норадреналин периферических тканей находится в симпатических нервных окончаниях, и густо иннервированные ткани содержат по 1— 2 мкг его на 1 г ткани. Норадреналин в нервных окончаниях находится в дискретных субцеллюлярных частицах, подобных хромаффинным гранулам мозгового вещества надпочечников. МАО, содержащаяся в миотохондриях нервных окончаний, играет важную роль в регуляции локальной концентрации норадреналина. Амины, находящиеся в луковицеобразных тельцах, защищены от окислительного дезаминирования; однако амины, локализованные внутри цитоплазмы, дезаминируются и превращаются в неактивные метаболиты. Высвобождение медиаторов из нервных окончаний происходит в ответ на раздражение и возникающие постсинаптические потенциалы передаются в виде импульсов по терминальным симпатическим волокнам (см. рис. 66-1).
Периферический адренергический нейроэффекторный синапс. Поглощение нейронами. Периферические симпатические нервные окончания участвуют в системе транспорта аминов, активно поглощая их из внеклеточной жидкости. Субстратами для этого процесса служит ряд синтетических аминов и аминов естественного происхождения. Поглощением нейронами, или обратным захватом, высвобожденного локально норадреналина заканчивается действие этого нейромедиатора и поддерживается постоянство его запасов (см. рис. 66-1).
Пресинаптическая модуляция. Характер взаимосвязи между прохождением импульсов по нервному волокну и высвобождением норадреналина находится под влиянием целого ряда факторов. Так, пониженная температура и ацидоз снижают количество норадреналина, высвобождаемого в ответ на импульсы симпатического нерва. Некоторые химические медиаторы воздействуют на периферическое симпатическое нервное окончание (определяемое как место пресинаптического действия), модифицируя характер передачи импульсов по симпатическим нервным волокнам путем влияния на количество норадреналина, выделяемого в ответ на нервные импульсы. Пресинаптическая модуляция может быть как подавляющей, так и усиливающей. Определенные модуляторы, такие как катехоламины и ацетилхолин, могут или угнетать, или усиливать высвобождение норадреналина, вызывая антагонистические эффекты, опосредуемые соответственно через различные адренергические или холинергические рецепторы. К веществам, оказывающим угнетающее действие на высвобождение норадреналина в пресинаптическом нервном .окончании, относятся: катехоламины (a2-адренорецептор), ацетилхолин (мускариновый рецептор), дофамин (D2-рецептор), гистамин (Н2-рецептор), серотонин, аденозин, энкефалины и простагландины. К числу усиливающих пресинаптических модуляторов относятся: катехоламины (бета2-адренорецептор), ацетилхолин (никотиновый рецептор) и ангиотензин II. Общее значение пресинаптической модуляции, так же как и относительную степень важности различных медиаторов, еще предстоит выяснить.
Пресинаптические адренорецепторы. Катехоламины уменьшают высвобождение норадреналина, действуя через Пресинаптические a-адренорецепторы по классической системе негативной обратной связи. Регуляция посредством обратной связи осложняется тем, что активация b-адренорецепторов усиливает высвобождение норадреналина. Были выдвинуты две гипотезы физиологического интегрирования антагонистических воздействий a- и b-адренорецепторов на высвобождение норадреналина. Одна из них заключается в том, что b-адренорецепторы опосредуют реакции, развивающиеся при более низких концентрациях агониста, чем реакция, опосредуемые через a-адренорецепторы, в то время как a-адреноопоссредуемые реакции доминируют при более высоких концентрациях агониста. Поэтому при низких уровнях симпатической стимуляции, когда концентрации норадреналина в синаптической щели низки, b-адреноопосредуемая положительная обратная связь может доминировать, в результате чего его высвобождение усилится. И наоборот, при высоких уровнях симпатической стимуляции; когда концентрация норадреналина в симпатической щели высока, доминирует a-адреноопосредуемая обратная связь и происходит угнетение высвобождения норадреналина. Другая гипотеза состоит в том, что Пресинаптические b-адренорецепторы более чувствительны к адреналину, чем к норадреналину; поэтому уровни содержания циркулирующего в крови адреналина могут стимулировать Пресинаптические b-адренорецепторы, тем самым увеличивая высвобождение норадреналина и усиливая симпатическую нейропередачу.
Пресинаптические холинергические рецепторы. Несмотря на то что было описано как угнетающее, так и усиливающее действие ацетилхолина на высвобождение норадреналина, угнетающее действие первого, опосредуемое через мускариновый холинергический рецептор, происходит при более низких его концентрациях и, вероятно, имеет большое физиологическое значение. Это периферическое угнетающее действие ацетилхолина на адренергическую нейропередачу может усиливать реципрокные изменения центральных парасимпатических и симпатических эфферентных импульсов, возникающих при регуляции различных физиологических реакций.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
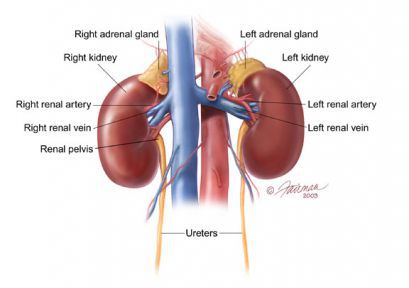
Мозговое вещество надпочечников и параганглии являются частью симпатического отдела вегетативной, или автономной, нервной системы.
Исторический очерк
В 1948 г. Алквист (Alquist), сравнив величину эффектов адреналина, норадреналина и изопротере-нола, предположил существование двух групп мембранных белков — адренорецепторов, которые он назвал а- и Р-адренорецепторами. В дальнейшем среди них были выделены α1A-, α1В-, α1С-, α2A-, α2В-, α2С-, β1-, β2-, β3-, β4-подтипы. Эти рецепторы по-разному распределены в ЦНС и периферических тканях.
В 1950 г. фон Эйлер и Энгель (Engel) обнаружили повышенную экскрецию адреналина и норадреналина с мочой у больных с феохромоцитомами, а после того, как Армстронг показал, что метаболитом катехоламинов является ванилилминдальная кислота (ВМК), именно ее экскрецию начали использовать в качестве диагностического показателя феохромоцитомы. В 1958 г. Лабросс (LaBrosse) выявил присутствие в моче норметанефрина. В 1970 г. фон Эйлер был удостоен Нобелевской премии за пионерские исследования в области запасания, секреции и инактивации катехоламинов.
Анатомия
Эмбриология
Симпатическая нервная система развивается у плода из примитивных клеток нервного валика (симпатогоний). Примерно на пятой неделе внутриутробной жизни эти клетки мигрируют из первичных спинальных ганглиев в грудную область, образуя симпатическую цепочку позади аорты. Затем они перемещаются вперед, формируя остальные симпатические ганглии.
На 6-й неделе внутриутробной жизни группы этих первичных клеток мигрируют вдоль центральной вены и проникают в кору надпочечников плода, образуя их мозговое вещество, которое удается различить на 8-й неделе. Мозговое вещество надпочечников в это время состоит из симпатогоний и феохромобластов, созревающих в дальнейшем в феохромоциты. Они формируют розеткообразные структуры с примитивными клетками в центре. На 12-й неделе при электронной микроскопии в этих клетках обнаруживаются плотные гранулы. У новорожденных мозговое вещество занимает очень небольшое место в ткани надпочечников и выглядит аморфным, но к 6 месяцам постнатальной жизни приобретает зрелый вид.
Феохромобласты и феохромоциты скапливаются также по обеим сторонам аорты, образуя параганглии. Основные скопления этих клеток обнаруживаются на уровне нижней брыжеечной артерии и бифуркации аорты. Сливаясь, они образуют орган Цуккеркандля, отчетливо видимый у плода. Считается, что именно этот орган служит основным источником катехоламинов в первый год жизни; затем он атрофируется. Феохромоциты (хромаффинные клетки) присутствуют также в абдоминальном симпатическом сплетении и других отделах симпатической нервной системы.
Макроскопическое строение
Анатомические связи между корковым и мозговым веществами надпочечников неодинаковы у разных видов. У акул эти органы полностью разделены. У амфибий они уже тесно контактируют друг с другом, а у птиц их клетки составляют единый орган. У человека мозговое вещество надпочечников занимает центральное положение в самой широкой части железы, и лишь немногие клетки проникают в более узкие части надпочечников. Масса мозгового вещества обоих надпочечников у взрослых составляет примерно 1000 мг (около 15% общей массы двух надпочечников), хотя ее доля у разных людей может быть разной. Четкая граница между корковым и мозговым веществом отсутствует. Клетки коркового вещества обычно окружают центральную вену в мозговом веществе и образуют в его толще небольшие островки.
Микроскопическое строение
Хромаффинные клетки, или феохромоциты, мозгового вещества надпочечников имеют форму крупных удлиненных овалов. Они располагаются в виде сети, альвеол или тяжей вокруг многочисленных капилляров и венозных синусов, куда поступает кровь из коркового вещества. Феохромоциты содержат крупные ядра и развитый аппарат Гольджи. В их цитоплазме присутствует большое число пузырьков, или гранул диаметром 100-300 нм, сходных с нейросекреторными гранулами периферических симпатических нервов. Катехоламины (адреналин и/или норадреналин) составляют примерно 20% массы нейросекреторных пузырьков. Пузырьки, содержащие норадреналин, выглядят темнее содержащих адреналин. В этих пузырьках присутствуют также белки, липиды, АТФ, а также хромогранины, нейропептид Y, энкефалины и проопиомеланокортин (вместе с АКТГ и β-эндорфином).
Иннервация
Клетки мозгового вещества надпочечников иннервируются преганглионарными симпатическими волокнами, которые выделяют ацетилхолин и энкефалины. Большинство этих волокон начинается в подкапсульном сплетении на задней поверхности надпочечников и, не прерываясь, проникает в них в виде пучков (по 30-50 волокон). Пучки преганглионарных волокон тянутся вдоль сосудов и достигают мозгового вещества, не разветвляясь в корковом. Некоторые волокна оканчиваются в стенке центральной вены. Однако большинство этих волокон оканчивается на феохромоцитах.
Кровоснабжение
Надпочечники человека получают кровь из верхней, средней и нижней надпочечниковых ветвей нижней диафрагмальной артерии, непосредственно из аорты и из почечных артерий. Под капсулой надпочечников сосуды образуют сплетение, питающее корковое вещество. Некоторые сосуды проникают прямо в мозговое вещество. Однако основное кровоснабжение клеток мозгового вещества осуществляется, по-видимому, сосудами портальной системы, берущими начало в капиллярах коркового вещества. Вокруг центральной вены надпочечников располагается также сплетение лимфатических капилляров.
У млекопитающих кортизол индуцирует синтез фермента, катализирующего превращение норадреналина в адреналин (фенилэтаноламин-N-метилтрасферазы, ФЭМТ). Хромаффинные клетки, содержащие адреналин, питаются в основном кровью сосудов, дренирующих корковое вещество, тогда как клетки, содержащие норадреналин, снабжаются артериями, проникающими непосредственно в мозговое вещество.
Короткая центральная вена правого надпочечника впадает прямо в нижнюю полую вену. Центральная вена левого надпочечника (несколько более длинная) впадает в левую почечную вену.
Важнейший регулятор (стимулятор) биосинтеза и секреции катехоламинов в надпочечниковой железе — симпатический большой чревный нерв (Чебоксаров, 1910) (рис. 32). Большой чревный нерв состоит в основном из преганглионарных, миелиновых волокон, берущих начало от симпатических центров, расположенных в грудной области спинного мозга. Ветви нерва, подходящие к надпочечникам, не прерываются в ганглиях и, следовательно, медиатором их возбуждения в надпочечниковой железе является ацетилхолин.
Последний выделяется в синаптическую щель из везикул пресинаптической мембраны в ответ на возбуждение, приходящее по нерву, взаимодействует с никотиновыми холинорецепторами и вызывает усиление синтеза и секреции адреналина и норадреналина железой (Теней и др., 1973). Хромаффинная ткань — гомолог симпатических ганглиев и, по существу, функционально эквивалентна постганглионарным (безмиелиновым) симпатическим волокнам и их синапсам.
Однако если постганглионарные нервные волокна выделяют в интрамуральных (внутриорганных) окончаниях в качестве медиатора преимущественно норадреналин, то мозговой слой надпочечников выделяет в кровь смесь обоих катехоламинов обычно с преобладанием адреналина. В связи с этим хромаффинную ткань можно рассматривать как модифицированное периферическое звено симпатической нервной системы, осуществляющее свое воздействие гуморально. Мозговой слой надпочечников й симпатическая нервная система функционируют в тесной взаимосвязи и представляют собой единую систему регуляции — симпато-адреналовую (Кеннон,1928; Орбели, 1928).
Таким образом, один из главных способов регуляции продукции катехоламинов в надпочечниках — нервно-рефлекторный механизм, эффекторное звено которого представлено большим чревным нервом. Центральное звено данной рефлекторной дуги находится в симпатических центрах спинного мозга и, кроме того, в вегетативных центрах гипоталамуса. Именно этим путем, начинающимся с экстеро- и интерорецепторов, осуществляется быстрый рефлекторный выброс катехоламинов надпочечниками в ответ на различные воздействия — эмоциональное возбуждение, болевые сигналы, мышечную нагрузку, охлаждение, гипоксию, гипотонию, гипогликемию и т.д. Наряду с секрецией симпатические влияния значительно ускоряют и синтез катехоламинов.
Нервно-рефлекторная регуляция — важнейший, но не единственный механизм контроля работы мозгового слоя надпочечников. Изменение уровня секреторных процессов в железе может происходить и в условиях ее денервации под влиянием прямых гуморальных воздействий. К числу физиологических факторов, гуморально стимулирующих выброс катехоламинов из надпочечников, относятся гипогликемия, гиполипоацидемия, действие ацетилхолина, инсулина, кортикостероидов и др. Действие ацетилхолина усиливается низкими концентрациями адреналина. Вместе с тем сами катехоламины ингибируют активность тирозингидроксилазы и тем самым тормозят собственный синтез.
Большинство гуморальных факторов воздействует главным образом на освобождение гормоноидов из секреторных гранул. Синтез же катехоламинов в условиях денервации железы после воздействия стимулирующих секрецию гуморальных агентов резко замедлен.
Центральная нервная система человека осуществляет контроль над деятельностью его организма и разделяется на несколько отделов. Головной мозг посылает и получает сигналы из организма и после их обработки имеет информацию о процессах. Нервная система разделяется на вегетативную и соматическую нервную системы.
Отличия вегетативной и соматической нервной системы
Соматическая нервная система регулируется сознанием человека и может управлять деятельностью скелетной мускулатуры. Все компоненты реакции человека на внешние факторы находятся под контролем полушарий мозга. Она обеспечивает сенсорные и моторные реакции человека, контролируя их возбуждение и торможение.
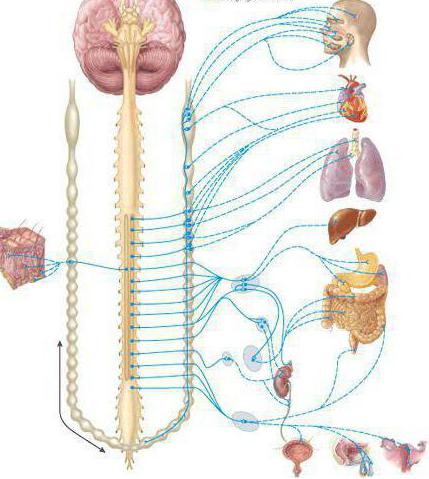
Вегетативная нервная система контролирует периферическую деятельность организма и не контролируется сознанием. Для нее характерны автономность и генерализованность воздействия на организм при полном отсутствии сознания. Эфферентная иннервация внутренних органов позволяет ей контролировать обменные процессы в организме и осуществлять обеспечение трофических процессов скелетной мускулатуры, рецепторов, кожи и внутренних органов.
Строение вегетативной системы
Работа вегетативной нервной системы контролируется гипоталамусом, который находится в центральной нервной системе. Вегетативная нервная система имеет метасегментарное строение. Ее центры находятся в головном, спинном мозге и коре головного мозга. Периферические отделы образованы стволами, ганглиями, сплетениями.
В вегетативной нервной системе различают:
- Симпатическую. Ее центр расположен в грудопоясничном отделе спинного мозга. Для нее характерны паравертебральные и предвертебральные ганглии ВНС.
- Парасимпатическую. Ее центры сосредоточены в среднем и продолговатом мозге, крестцовом отделе спинного мозга. Нервные узлы в основном интрамуральные.
- Метасимпатическую. Иннервирует желудочно-кишечный тракт, сосуды, внутренние органы организма.
В состав ее входит:
- Ядра нервных центров, расположенных в головном и спинном мозге.
- Вегетативные ганглии, которые расположены по периферии.
- Нервные волокна.
Рефлекторная дуга автономной нервной системы
Рефлекторная дуга вегетативной нервной системы состоит из трех звеньев:
- чувствительное или афферентное;
- вставочное или ассоциативное;
- эффекторное.

Чувствительное звено
Чувствительное звено расположено в спинномозговом ганглии. Этот ганглий имеет нервные клетки, сформированные группами, и их контроль осуществляется ядрами центрального головного мозга, большими полушариями и их структурами.
Чувствительное звено представлено частично униполярными клетками, которые имеют один приносящий или уносящий аксон, и они принадлежат спинальным или черепно-мозговым узлам. А также узлами блуждающих нервов, имеющих строение, похожее на спинальные клетки. В это звено входят клетки Догеля II типа, которые являются компонентами вегетативных ганглиев.
Вставочное звено
Вставочное звено в вегетативной нервной системе служит для передачи через низшие нервные центры, которыми являются вегетативные ганглии, и осуществляется это через синапсы. Расположено оно в боковых рогах спинного мозга. Нет прямой связи от афферентного звена на преганглионарные нейроны для их связи, существует кратчайший путь от афферентного нейрона до ассоциативного и от него до преганглионарного нейрона. Передача сигналов и нервных импульсов от афферентных нейронов в различных центрах осуществляется с различным количеством вставочных нейронов.
Например, в дуге спинального автономного рефлекса между чувствительным и эффекторным звеном существует три синапса, два из которых расположены в спинном мозге, а один в вегетативном узле, в котором расположен эфферентный нейрон.
Эфферентное звено
Эфферентное звено представлено эффекторными нейронами, которые расположены в вегетативных узлах. Их аксоны образуют безмиелиновые волокна, которые в составе со смешанными нервными волокнами иннервируют внутренние органы.
Вегетативные рефлекторные дуги расположены в боковых рогах.
Строение нервного узла
Ганглий – это скопление нервных клеток, которые имеют вид узелковых расширений толщиной около 10 мм. По своему строению вегетативный ганглий сверху покрыт соединительнотканной капсулой, которая образует строму из рыхлой соединительной ткани внутри органов. Мультиполярные нейроны, которые строятся из округлого ядра и крупных ядрышек, состоят из одного эфферентного нейрона и нескольких расходящихся афферентных нейронов. Эти клетки относятся по своему типу к клеткам головного мозга и являются двигательными. Их окружает неплотная оболочка – мантийная глия, которая создает постоянную среду для нервной ткани и обеспечивает полноценное функционирование нервных клеток.
Вегетативный ганглий имеет диффузное расположение нервных клеток и множество отростков, дендритов и аксонов.
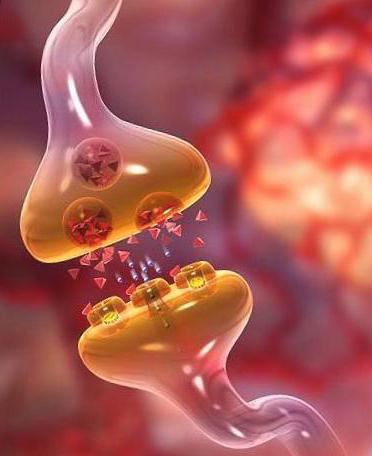
Спинномозговой ганглий имеет нервные клетки, которые расположены группами, и их расположение имеет порядок обусловленный.
Вегетативные нервные ганглии разделяются на:
- Сенсорные нейроны, которые расположены близко к спинному или центральному отделу мозга. Униполярные нейроны, из которых состоит этот ганглий, представляют собой приносящий или уносящий отросток. Они служат для афферентной передачи импульсов, и их нейроны образуют бифуркацию при ветвлении отростков. Эти отростки передают информацию от периферии к центральному афферентному нейрону – это периферический отросток, центральный – от тела нейрона в мозговой центр.
- Моторные, двигательные состоят из эфферентных нейронов, и в зависимости от их положения их называют паравертебральные, предвертебральные.
Симпатические ганглии
Паравертебральные цепочки ганглиев расположены вдоль позвоночного столба в симпатических стволах, которые идут длинной вереницей от основания черепа до копчика.
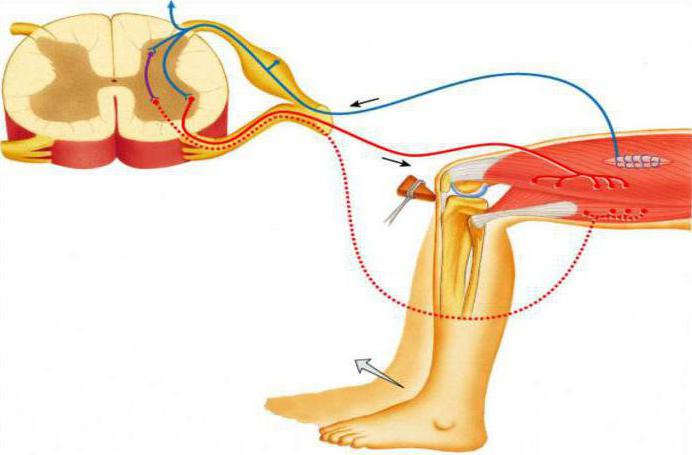
Предвертебральные нервные сплетения находятся ближе к внутренним органам, и их локализация сосредоточена перед аортой. Они образуют брюшное сплетение, которое состоит из солнечного, нижне- и верхнебрыжеечного сплетений. Они представлены двигательными адренергическими и тормозящими действие холинергическими нейронами. Также связь между нейронами осуществляется преганглионарными и постганглионарными нейронами, которые используют медиаторы ацетилхолин и норадреналин.
Интрамуральные нервные узлы имеют нейроны трех типов. Их описание было сделано русским ученым Догелем А.С., который, исследуя гистологию нейронов вегетативной нервной системы, выделил такие нейроны, как длинноаксонные эфферентные клетки первого типа, равноотросчатые афферентные клетки второго типа и ассоциативные клетки третьего типа.
Рецепторы ганглиев
Афферентные нейроны выполняют узкоспециализированую функцию, и их роль заключается в восприятии раздражителей. Такими рецепторами являются механорецепторы (реакция на растяжение или давление), фоторецепторы, терморецепторы, хеморецепторы (отвечают за реакции в организме, химические связи), ноцицепторы (реакция организма на болевые раздражители – это повреждение кожи и другие).
В симпатических стволах эти рецепторы по рефлекторной дуге передают в центральную нервную систему информацию, которая служит сигналом о повреждениях или нарушениях в организме, а также нормальной его работе.
Функции ганглиев
Каждый ганглий имеет свое местоположение, кровоснабжение, и его функции определяются этими параметрами. Спинномозговой ганглий, имеющий иннервацию из ядер головного мозга, обеспечивает непосредственную связь процессов в организме через рефлекторную дугу. От этих структурных компонентов спинного мозга иннервируются железы, гладкая мускулатура мышц внутренних органов. Сигналы, поступающие по рефлекторной дуге, идут медленнее, чем в ЦНС, и они полностью регулируются вегетативной системой, также она обладает трофической, сосудодвигательной функцией.
Читайте также:
