
Нейронные цепи в цнс
Нервная система человека и животных может быть представлена как система нейронных цепочек, передающих возбуждающие и тормозные сигналы (нервная сеть). Эти элементарные нейронные цепи служат, например, для усиления слабых сигналов, уменьшения слишком интенсивной активности, выделения контрастов, поддержания ритмов или сохранения рабочего состояния нейронов путем регулировки их входов. Такие нейронные цепи построены из стандартных элементов, которые выполняют наиболее часто повторяющиеся операции и могут быть включены в схемы самых разнообразных нервных структур.
Существуют значительные количественные различия нервных сетей у разных видов позвоночных и беспозвоночных. Так, у человека нервная система включает около 10 10 элементов, у примитивных беспозвоночных - около 10 4 нейронов, Однако в строении и функционировании всех нервных систем имеются общие черты. Практически во всех отделах центральной нервной системы обнаружены дивергенция нервных путей, конвергенция нервных путей и различные варианты тормозных связей между элементами нервных цепочек.
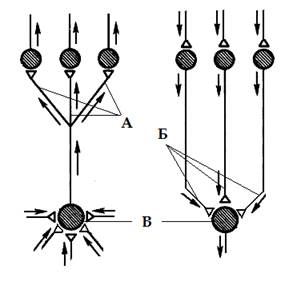
Дивергенция и конвергенция путей. Дивергенция (расхождение) пути (рис 2.А) - возникает в результате контактирования одного нейрона с множеством нейронов более высоких порядков. Так, например, происходит разделение аксона чувствительного нейрона, входящего в спинной мозг, на множество веточек (коллатералей), которые направляются к разным сегментам спинного мозга и в головной мозг, где происходит передача сигнала на вставочные и далее - на моторные нервные клетки. Дивергенция пути сигнала наблюдается так же у вставочных и у эффекторных нейронов.
Дивергенция пути обеспечивает расширение сферы действия сигнала, благодаря ей, информация поступает одновременно к разным участкам ЦНС. Это называют иррадиацией возбуждения (или торможения). Дивергенция настолько обычное явление, что можно говорить о принципе дивергенции в нейронных цепях.
| Конвергенция - это схождение многих нервных путей к одним и тем же нейронам (рис 2.Б). Например, у позвоночных на каждом мотонейроне спинного мозга и ствола головного мозга образуют | Рис 2. Дивергенция (А), конвергенция (Б) и пространственная суммация (В) нервных путей в центральной нервной системе. |
нервные окончания тысячи сенсорных, а также возбуждающих и тормозных вставочных нейронов разных уровней. Мощная конвергенция обнаруживается и на нейронах ретикулярной формации ствола мозга, на многих корковых нейронах у позвоночных и, видимо, на командных нейронах.
Конвергенция многих нервных путей к одному нейрону делает этот нейрон интегратором соответствующих сигналов. Вероятность возбуждения такого нейрона-интегратора зависит не от каждого пришедшего стимула в отдельности, а от суммы и направления стимулов, действующих одновременно, то есть суммы всех синаптических процессов происходящих на его плазматической мембране. Другими словами, вероятность распространения возбуждения через нейрон-интегратор определяется алгебраическим сложением величин возбуждающих и тормозных входов на нем, активных в данный момент. Такое сложение является результатом или пространственной или временной суммации. Пространственная суммация – результат сложения нервных импульсов приходящих одновременно к нейрону через разные синапсы (рис 2.В), временная суммация – сложение приходящих поочередно, через один синапс с небольшими интервалами времени. В обоих случаях нейрон интегратор, называют общим путем для конвергирующих на него нервных сигналов, а если речь идет о мотонейроне, т. е. конечном звене нервного пути к мускулатуре, говорят об общем конечном пути.
Результат суммации заключается в возможности изменения направления распространения возбуждения в ЦНС, (то есть не строго в пределах одной рефлекторной дуги), а значит и в изменении характера ответной реакции организма в ответ на действие раздражителя. Ответ организма, реализуемый в результате, становится более адекватным внешним условиям и состоянию нервной системы. Пример такого выбора ответа можно видеть, если речь идет о конвергенции не на одном нейроне, а на группе нейронов совместно регулирующих общую функцию, что в ЦНС не редкость. Наличие конвергенции множества путей на одной группе мотонейронов лежит в основе феноменов пространственного облегчения и окклюзии.
Пространственное облегчение - это превышение эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в ЦНС над суммой их раздельных эффектов. Т.е. при раздельном действии афферентных сигналов возбуждение возникает в меньшем числе эфферентных нейронов и эффект оказывается слабее. Феномен объясняется суммацией совместно возникающих ВПСП до критического уровня деполяризации в группе мотонейронов, в которых при раздельной активации входов ВПСП оказывались слишком слабыми для генерации ответа.
Окклюзия - это явление, противоположное пространственному облегчению. В этом случае эффект окажется выше если афферентные сигналы действуют порознь, а при их совместном действии возбуждается меньшая группа мотонейронов. Причина окклюзии состоит в том, что здесь афферентные входы в силу конвергенции частично связаны с одними и теми же мотонейронами, и каждый может возбуждать их, как и оба входа вместе.
Таким образом, если эффект нескольких стимулов, поступающих одновременно или в быстрой последовательности будет выше, чем сумма эффектов отдельных стимулов то это явление называется облегчением; если же эффект на сочетание стимулов меньше, чем сумма ответов на отдельные стимулы, то такое явление - окклюзия.
Такое явление следует учитывать, например, при тренировке различных функцинальных показателей скелетных мышц.
Простые тормозные и усиливающие цепи.
Тормозные цепи, виды торможения. Торможение, как и возбуждение, - активный процесс, оно возникает в результате сложных физико-химических изменений в тканях. Благодаря процессу торможения достигается ограничение распространения возбуждения в ЦНС и обеспечивается координация рефлекторных актов, внешне этот процесс проявляется ослаблением функции какого-либо органа.
К настоящему времени анализ тормозных явлений в ЦНС позволил выделить две формы разновидности постсинаптическое и пресинаптическое торможение.
Постсинаптическое торможение развивается не постсинаптических мембранах межнейронных синапсов и связано с гиперполяризацией постсинаптической мембраны под влиянием медиаторов, которые выделяются при возбуждении специальных тормозных нейронов. При этом, локально возникающая на постсинаптической мембране гиперполяризация - тормозной постсинаптический потенциал (ТПСП) – затрудняет электротоническое распространение возбуждающих постсинаптических потенциалов (ВПСП) от других синапсов, к аксонному холмику. В результате в зоне аксонного холмика не происходит выведение мембранного потенциала на критический уровень. Потенциал действия не образуется, нейрон не возбуждается.
Постсинаптическое торможение активно используется в нейронных сетях, и в зависимости от вариантов связывания нейронов друг с другом выделяют несколько его видов: реципрокное (прямое), параллельное, возвратное, латеральное (рис.3)
Реципрокное торможение (рис 3.А) – это взаимное (сопряженное) торможение центров антагонистических рефлексов, обеспечивающее координацию этих рефлексов. Классический пример реципрокного торможения - это торможение мотонейронов мышц-антагонистов у позвоночных. Торможение осуществляется с помощью специальных тормозных вставочных нейронов. При активации путей, возбуждающих, например, мотонейроны мышц-сгибателей, мотонейроны мышц-разгибателей тормозятся импульсами вставочных клеток.
Возвратное торможение (рис. 3.Б) - это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям к тормозным клеткам. Возвратное торможение наблюдается, например, в мотонейронах спинного мозга позвоночных. Эти клетки отдают возвратные коллатерали в мозг к тормозным вставочным клеткам Реншоу, которые имеют синапсы на этих же мотонейронах. Торможение обеспечивает ограничение ритма мотонейронов, позволяющее чередовать сокращение и расслабление скелетной мышцы, что важно для нормальной работы двигательного аппарата.
Такую же роль играет возвратное торможение и в других нервных сетях.
Параллельное торможение (рис. 3.В) – играет сходную с возвратным роль, но в этом случае возбуждение блокирует само себя, посылая тормозной сигнал на нейрон который одновременно и активирует.
Это возможно, если возбуждающий импульс сам не должен вызвать возбуждения на нейроне-мишени, но его роль важна при пространственной суммации, в комбинации с другими сигналами.
Латеральное торможение (рис. 3.Г) – это торможение нервных клеток, расположенных по соседству с активной, которое этой клеткой и инициируется. При этом вокруг возбужденного нейрона возникает зона, в которой развивается очень глубокое торможение.
Латеральное торможение наблюдается, например, в конкурирующих сенсорных каналах связи. Оно наблюдается у соседних элементов сетчатки позвоночных, а также в их зрительных, слуховых и других сенсорных центрах. Во всех случаях латеральное торможение обеспечивает контраст, т. е. выделение существенных сигналов или их границ из фона.
Пресинаптическое торможение развивается в аксо-аксональных синапсах, образованных на пресинаптических окончаниях нейрона.
В основе пресинаптического торможения лежит развитие медленной и длительной деполяризации пресинаптического окончания, что и приводит к развитию торможения. В деполяризованном участке нарушается процесс распространения возбуждения и поступающие к нему импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуде, не обеспечивают выделения достаточного количества медиатора – нейрон не возбуждается.
Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные нейроны, аксоны которых и образуют синапсы на пресинаптических окончаниях аксона-мишени.
Разновидности пресинаптического торможения изучены недостаточно, вероятно они те же, что и для постсинаптического торможения. Точно известно о наличии параллельного и латерального пресинаптического торможения (рис. 4).
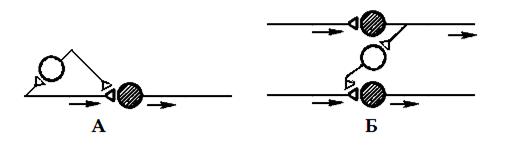
Рис. 4. Разновидности пресинаптического торможения: А – параллельное, Б – латеральное. Темные нейроны – возбуждающие, светлые – тормозные.
В реальной действительности взаимоотношения возбуждающих и тормозных нейронов значительно сложнее, чем представлено на рисунках, тем не менее, все варианты пре- и постсинаптического торможения можно объединить в две группы. Во-первых, когда блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное и возвратное торможение), во-вторых, когда блокируются другие нервные элементы под влиянием импульсов от соседних возбуждающих нейронов с включением тормозных клеток (латеральное и прямое торможение).
Кроме того, тормозные клетки сами могут быть заторможены другими тормозными нейронами, это может облегчить распространение возбуждения.
Роль процесса торможения.
- Оба известных вида торможения со всеми их разновидностями выполняют, прежде всего, охранительную роль. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов, утомлению, истощению и прекращению деятельности ЦНС.
- Торможение играет важную роль в обработке поступающей в ЦНС информации. Особенно ярко выражена эта роль у пресинаптического торможения. Оно более точно регулирует процесс возбуждения, поскольку этим торможением могут быть полностью заблокированы отдельные нервные волокна. К одному возбуждающему нейрону могут приходить сотни и тысячи различных импульсов по разным путям, но число дошедших до нейрона импульсов определяется пресинаптическим торможением.
- Поскольку блокада торможения ведет к широкой иррадиации возбуждения и судорогам, следует признать, что торможение является важным фактором обеспечения координационной деятельности ЦНС.
Усиливающие цепи и механизмы усиления. Нейронные сети имеют не только тормозные механизмы, препятствующие распространению возбуждения, но и системы, усиливающие приходящий к ним сигнал. Рассмотрим некоторые из них.
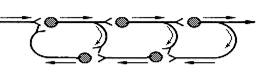
Самовозбуждающиеся нервные цепи (цепи с положительной обратной связью) (рис.5). Некоторые данные свидетельствуют о том, что в мозгу животных и человека существуют замкнутые самовозбуждающиеся цепочки нейронов, в которых нейроны соединены синапсами возбуждающего действия. Возникнув в ответ на внешний сигнал, возбуждение в такой цепочке циркулирует, иначе реверберирует, до тех пор, пока или какой-либо внешний тормоз не выключит одно из звеньев цепи, или в ней не наступит утомление. Выходные пути от такой цепочки (ответвляющиеся по коллатералям аксонов нервных клеток - участников цепи) во время работы передают равномерный поток импульсов, создающий ту или иную настройку в нервных клетках-мишенях. Ее функции могут состоять в том, чтобы обеспечивать длительное поддержание индуцированной однажды активности.
сигнал, который включил в ней циркуляцию (реверберацию) импульсов. Считают, что это возможный механизм (или один из механизмов) краткосрочной памяти, однако этому практически нет экспериментальных доказательств.
Синаптическая потенциация — увеличение амплитуды постсинаптического потенциала, если интервал между последовательным возникновением потенциалов действия в пресинаптической мембране невелик, то есть происходит частая и ритмическая активация синапса. Явление потенциации связывают с накоплением ионов кальция в пресинаптическом окончании, который дополнительно вбрасывается туда при каждом новом стимуле и не успевает полностью удаляться между частыми стимулами. Вследствие этого, каждый новый пресинаптический потенциал вызывает высвобождение большего числа квантов медиатора.
Такую же природу имеет и посттетаническая потенциация. В этом случае увеличение числа квантов медиатора, высвобождаемых нервным импульсом, после предшествующего ритмического раздражения приводит к увеличению синаптической реакции нейрона на одиночное раздражение пресинаптических путей. Посттетаническая потенциация может длиться от нескольких минут до нескольких часов в различных структурах мозга. Предполагают, что постсинаптическая потенциация играет важную роль в пластических перестройках функций синапсов, и лежит в основе механизмов организации условных рефлексов и памяти. Например, особенно длительная посттетаническая потенциация обнаружена в гиппокампе – структуре, которая, играет важную роль в явлениях памяти и научения.
Ритмическая стимуляция может приводить и к снижению активности синапсов. Процесс снижения постсинаптических потенциалов во время или по окончании тетанической стимуляции по сравнению с исходной амплитудой называется синаптической депрессией; по аналогии с потенциацией, различают тетаническую и посттетаническую депрессию. Возможно, синаптическая депрессия имеет место во многих участках нервной системы и является нейронным коррелятом привыкания (габитуации). У беспозвоночных габитуация простых поведенческих реакций прямо соответствует депрессии участвующих синапсов; то же самое относится и к флексорному рефлексу у кошки. Таким образом, синаптическая депрессия, так же как синаптическая потенциация, составляет элементарный процесс научения.
1.Понятие нервных центров
Нервный центр — центральный компонент рефлекторной дуги, где происходит переработка информации, вырабатывается программа действия, формируется эталон результата.
2.Свойства нервных центров
1)Одностороннее проведение возбуждения — возбуждение передается с афферентного на эфферентный нейрон. Причина: клапанное свойство синапса.
2)Задержка проведения возбуждения: скорость проведения возбуждения в нервном центре на много ниже таковой по остальным компонентам рефлекторной дуги. Чем сложнее нервный центр, тем дольше проходит по нему нервный импульс. Причина: синаптическая задержка. Время проведения возбуждения через нервный центр — центральное время рефлекса.
3)Суммация возбуждения — при действии одиночного подпорогового раздражителя ответной реакции нет. При действии нескольких подпороговых раздражителей ответная реакция есть. Рецептивное поле рефлекса — зона расположения рецепторов, возбуждение которых вызывает определенный рефлекторный акт.
Имеется 2 вида суммации: временная и пространственная.
Временная суммация — возникает ответная реакция при действии нескольких следующих друг за другом раздражителей. Механизм: суммируются возбуждающие постсинаптические потенциалы рецептивного поля одного рефлекса. Происходит суммация во времени потенциалов одних и тех же групп синапсов.
Пространственная суммация — возникновение ответной реакции при одновременном действии нескольких подпороговых раздражителей. Механизм: суммация возбуждающего постсинаптического потенциала от разных рецептивных полей. Суммируются потенциалы разных групп синапсов.
4)Центральное облегчение — объясняется особенностями строения нервного центра. Каждое афферентное волокно входя в нервный центр иннервирует определенное количество нервных клеток. Эти нейроны — нейронный пул. В каждом нервном центре много пулов. В каждом нейронном пуле — 2 зоны: центральная (здесь афферентное волокно над каждым нейроном образует достаточное для возбуждения количество синапсов), периферическая или краевая кайма (здесь количество синапсов недостаточно для возбуждения). При раздражении возбуждаются нейроны центральной зоны. Центральное облегчение: при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть больше арифметической суммы раздражения каждого из них, т. к. импульсы от них отходят к одним и тем же нейронам периферической зоны.
5)Окклюзия — при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть меньше арифметической суммы раздражения каждого из них. Механизм: импульсы сходятся к одним и тем же нейронам центральной зоны. Возникновение окклюзии или центрального облегчения зависит от силы и частоты раздражения. При действии оптимального раздражителя, (максимального раздражителя (по силе и частоте) вызывающего максимальную ответную реакцию) — появляется центральное облегчение. При действии пессимального раздражителя (с силой и частотой вызывающих снижение ответной реакции) — возникает явление окклюзии.
5)Посттетаническая потенция — усиление ответной реакции, наблюдается после серии нервных импульсов. Механизм: потенциация возбуждения в синапсах;
6)Рефлекторное последействие — продолжение ответной реакции после прекращения действия раздражителя:
кратковременное последействие — в течение нескольких долей секунды. Причина — следовая деполяризация нейронов;
длительное последействие — в течение нескольких секунд. Причина: после прекращения действия раздражителя возбуждение продолжает циркулировать внутри нервного центра по замкнутым нейронным цепям.
7)Трансформация возбуждения — несоответствие ответной реакции частоте наносимых раздражений. На афферентном нейроне происходит трансформация в сторону уменьшения из-за низкой лабильности синапса. На аксонах эфферентного нейрона, частота импульса больше частоты наносимых раздражений. Причина: внутри нервного центра образуются замкнутые нейронные цепи, в них циркулирует возбуждение и на выход из нервного центра импульсы подаются с большей частотой.
8)Высокая утомляемость нервных центров — связана с высокой утомляемостью синапсов.
Тонус нервного центра — умеренное возбуждение нейронов, которое регистрируется даже в состоянии относительного физиологического покоя. Причины: рефлекторное происхождение тонуса, гуморальное происхождение тонуса (действие метаболитов), влияние вышележащих отделов центральной нервной системы.
Вопрос 2 Изосерологические системы крови человека. Групповая система АВО и система резус – характеристика антигенов и антител и методы определения групповой и резус-принадлежности.
ГРУППОВАЯ СИСТЕМА АВО.
С открытием венским врачом К. Ландштейнером (1901) групп крови стало понятно, почему в одних случаях трансфузии крови проходят успешно, а в других заканчиваются трагически для больного. К. Ландштейнер впервые обнаружил, что плазма одних людей способна склеивать эритроциты других людей. В основе ее лежит наличие в эритроцитах антигенов, названных агглютиногенами и обозначаемых буквами А и В, а в плазме — природных антител, или агглютининов, именуемых α и β. Агглютинация эритроцитов наблюдается лишь в том случае, если встречаются одноименные агглютиноген и агглютинин: А и α, В и β.
Установлено, что агглютинины, являясь природными антителами (AT), имеют два центра связывания, а потому одна молекула агглютинина способна образовать мостик между двумя эритроцитами. При этом каждый из эритроцитов может при участии агглютининов связаться с соседним, благодаря чему возникает конгломерат (агглютинат) эритроцитов.
В крови одного и того же человека не может быть одноименных агглютиногенов и агглютининов, так как в противном случае происходило бы массовое склеивание эритроцитов, что несовместимо с жизнью. Возможны только четыре комбинации, при которых не встречаются одноименные агглютиногены и агглютинины, или четыре группы крови:
Кроме агглютининов, в плазме, или сыворотке, крови содержатся гемолизины: их также два вида и они обозначаются, как и агглютинины, буквами α и β. При встрече одноименных агглютиногена и гемолизина наступает гемолиз эритроцитов. Действие гемолизинов проявляется при температуре 37—40 οС. Вот почему при переливании несовместимой крови у человека уже через 30—40 с. наступает гемолиз эритроцитов.
Для решения вопроса о совместимости групп крови пользуются следующим правилом:
среда реципиента должна быть пригодна для жизни эритроцитов донора. Такой средой является плазма, следовательно, у реципиента должны учитываться агглютинины и гемолизины, находящиеся в плазме, а у донора — агглютиногены, содержащиеся в эритроцитах.
Кровь I группы совместима со всеми другими группами крови, поэтому человек, имеющий I группу крови, называется универсальным донором. С другой стороны, эритроциты IV группы крови не должны давать реакции агглютинации при смешивании с плазмой людей с любой группой крови, поэтому люди с IV группой крови называются универсальными реципиентами.
В повседневной практике для решения вопроса о группе переливаемой крови пользуются иным правилом: переливаться должны одногруппная кровь и только по жизненным показаниям, когда человек потерял много крови. Лишь в случае отсутствия одногруппной крови с большой осторожностью можно перелить небольшое количество иногруппной совместимой крови.
Существуют две основных номенклатуры обозначения антигенов этой системы: по Ландштейнеру и Винеру и по Фишеру Р. и Раису Р. Современная номенклатура — это совмещение двух номенклатур. Антигены Современныйвариант: Rho (D): rh’(C): rhu(E): Нго (d): hr’(c): hr”(e)
Наиболее активным в антигенном отношении является антиген D, в меньшей степени — СиЕ, а тем более d, с, е. Реципиент имеет резус-положительную кровь, если его эритроциты обязательно содержат антиген D. Антиген D выявляется у 86% людей, С — у 70,8%, Е — у 31,0%, d — у 99%, с -*иу 84%; е — у 86%. Учитывая, что антиген D определяет принадлежность людей к группе резус-положительных, таких людей среди европейцев много — 86%, у представителей монгольской расы — 100%. Антиген D является основной причиной сенсибилизации (иммунизации) во время бере-менности и гемолитической болезни новорожденных, он легко проникает через плаценту.
В настоящее время известны и другие факторы резус-системы. Из них особый интерес представляет вариант фактора D, который обозначается D . Он не всегда определяется в эритро-
цигах, но в ответ на его введение у резус-отрицательного человека вырабатывается анги-D. Поэтому у резус-отрицательного человека необходимо определить и отсутствие антигена D".
В эритроците антигены системы резус находятся в виде группы антигенов. Наиболее частые комбинации такие: CDE — 16%, CDe — 53%, cDE — 15%, cde — 12%. У аборигенов Австралии в эритроцитах не выявлен ни один представитель системы резус. Такой вариант называют резус-нуль.
Для определения резус-принадлежности, т.е. выявления антигенов системы резус в эритроцитах используют стандартные сыворотки (реагенты) антирезус, различные по специфичности, т.е. содержащие антитела к разным антигенам этой системы. Для определения антигена D чаще всего применяют сыворотку антирезус с добавлением 10% раствора желатина или используют стандартный реагент антирезус, приготовленный заранее с 33% раствором полиглкжина.
Антигенами называются структурно чужеродные для данного конкретного организма вещества (высокомолекулярные соединения - белки и полисахариды), способные вызвать иммунный ответ (начальные слоги двух слов: "АНТИтела ГЕНерация"). Носителями таких чужеродных веществ будут бактерии, вирусы, грибки, трансплантаты, опухолевые клетки.
Антигены состоят из крупной неспецифической молекулы - переносчика (полисахарида, белка или липида с молекулярным весом более 10 000) и расположенных на поверхности этой молекулы детерминантных групп ( антигенных детерминант ), обусловливающих серологическую специфичность антигена. Детерминантные группы, отделенные от макромолекулы - переносчика, называются гаптенами . Гаптены приобретают иммуногенность лишь после соединения с высокомолекулярным белком-носителем. Гаптены не могут стимулировать выработку антител, но могут связываться с ними. Следует подчеркнуть, что иммуногенность - комплексная характеристика, которая зависит от свойств самого антигена, пути его введения и способа иммунизации. Они реагируют с соответствующими (гомологичными) антителами, но не запускают синтез новых антител.
Термин антиген используется в двух случаях.
Во-первых, так называют молекулы, которые индуцируют иммунный ответ. Эти молекулы еще называют иммуногенами .
Во-вторых, антигенами называют молекулы, которые реагируют с антителами илипримированными T-лимфоцитами . При этом не имеет значения, способны ли эти молекулы сами по себе индуцировать образование таких антител или T-лимфоцитов.
Два значения слова "антиген" не всегда совпадают.
Гаптены имеют по одному эпитопу, тогда как полисахариды и гомополимеры имеют множественное число эпитопов одной и той же специфичности. Белки несут на своей поверхности множество эпитопов, но уже разной специфичности.
Если антиген - линейный пептид или полисахарид, то во взаимодействии с антителом принимают участие около 5-6 аминокислотных остатков или молекул гексозы, соответственно. Если же антиген - глобулярный белок, то с антителом может контактировать до 16 аминокислотных остатков.
Если схематически изобразить поверхность белкового антигена и отметить на ней центры эпитопов, выявлямых с помощью индивидуальных антител, входящих в данную сыворотку, то на этой карте можно будет выявить кластеры доминантных эпитопов. Именно эти кластеры можно определить как антигенные детерминанты . Важно иметь в виду, что на поверхности антигена может находиться несколько антигенных детерминант различной структуры.
Основные классы антигенов - это углеводы ( полисахариды ), липиды , нуклеиновые кислоты и белки. Антитела – специфические белки гамма- глобулиновой природы, образующиеся в организме в ответ на антигенную стимуляцию и способные специфически взаимодействовать с антигеном
Основные биологические характеристики антител.
1. Специфичность – способность взаимодействия с определенным (своим) антигеном (соответствие эпитопа антигена и активного центра антител).
2. Валентность- количество способных реагировать с антигеном активных центров ( это связано с молекулярной организацией- моно- или полимер). Иммуноглобулины могут быть двухвалентными ( IgG ) илиполивалентными (пентамер IgM имеет 10 активных центров). Двух- и более валентные антитела навываютполными антителами. Неполные антитела имеют только один участвующий во взаимодействии с антигеном активный центр ( блокирующий эффект на иммунологические реакции, например, на агглютинационные тесты). Их выявляют в антиглобулиновой пробе Кумбса, реакции угнетения связывания комплемента.
3. Афинность – прочность связи между эпитопом антигена и активным центром антител, зависит от их пространственного соответствия.
4. Авидность – интегральная характеристика силы связи между антигеном и антителами, с учетом взаимодействия всех активных центров антител с эпитопами. Поскольку антигены часто поливалентны, связь между отдельными молекулами антигена осуществляется с помощью нескольких антител.
5. Гетерогенность – обусловлена антигенными свойствами антител, наличием у них трех видов антигенных детерминант:
- изотипические – принадлежность антител к определенному классу иммуноглобулинов;
- аллотипические- обусловлены аллельными различиями иммуноглобулинов, кодируемых соответствующими аллелями Ig гена;
- идиотипические- отражают индивидуальные особенности иммуноглобулина, определяемые характеристиками активных центров молекул антител. Даже тогда, когда антитела к конкретному антигену относятся к одному классу, субклассу и даже аллотипу, они характеризуются специфическими отличиями друг от друга (идиотипом). Это зависит от особенностей строения V- участков H- и L- цепей, множества различных вариантов их аминокислотных последовательностей.
На протяжении многих веков человеческий организм, его жизнеобеспечивающие функции, психика и система восприятия являлись одним из самых трудоёмких объектов для всестороннего и глубокого изучения. С появлением новых направлений и исследований в медицине, генетике, нейробиологии и психологии стало возможным провести множество параллелей о взаимном влиянии наших психоментальных процессов и биологических функций организма.
В данной статье мы обзорно рассмотрим с позиции нейробиологии и ииссиидиологии взаимосвязи некоторых отделов мозга и проходящих в них нейрональных цепочек с психологическими состояниями и привычками человека.
Нейронная сеть (нервная система человека) – сложная сеть структур, которая обеспечивает взаимосвязанное поведение разных систем в организме. Нейрон – это специальная клетка, состоящая из ядра, тела и многочисленных отростков – дендритов (длинные отростки называют аксонами). Зоны контакта между нейронами называются синапсами. Средний человеческий мозг располагает ресурсом в 100 млрд. нейронов. Каждая клетка, в свою очередь, может дать примерно 200 тыс. синаптических ответвлений. В нейронах происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма на внешние и внутренние раздражения [1].
Одна из ключевых задач нейрона - передача электрохимического импульса по нейронной цепи по доступным (резонационным) связям с другими нейронами. При этом каждая из связей характеризуется некоторой величиной, которая называется синаптической силой. Она определяет, что будет происходить с электрохимическим импульсом при его передаче другому нейрону: усилится он, ослабеет или останется неизменным.
Биологическая нейронная сеть имеет высокую степень связности: на один нейрон приходится порою несколько тысяч связей с остальными нейронами. Однако это приблизительное значение, которое в каждом из конкретных случаев разное. Передача импульсов от нейрона к нейрону порождает некоторое возбуждение всей сети нейронов. Несколько упрощая, можно сказать, что каждая нейронная сеть представляет собой мысль, навык, воспоминание, то есть некий блок информации.
Здесь мне бы хотелось подчеркнуть, что конкретика этих убеждений будет зависеть от того, в каком отделе головного мозга будут чаще всего задействованы нейронные цепочки.
На сегодняшний день считается, что отличительной особенностью человеческого мозга от мозга животных является заметно увеличенные области лобных долей, которые представляют собой один из отделов неокортекса (от лат. neo – новая, cortex- кора). Этот отдел полушарий головного мозга в процессе эволюции был сформирован довольно поздно. И если у хищников он едва намечен, то у современного человека лобные доли занимают около 25% общей площади больших полушарий мозга.
В тоже время каждый из нас может себя узнать и в таких проявлениях как резкие перепады настроения, окрашенные пессимистичным или негативным взглядом на жизнь, снижение мотивации, устремления, самооценки, усиление чувства вины или беспомощности и многие другие подобные состояния.
Одной из основных причин является ещё достаточно устойчивая сосредоточенность личности на собственных интересах. По этой причине задействуются отделы мозга, связанные с эмоциями и получением различных видов удовольствий: тактильных, вкусовых, обонятельных, эстетических и других; в корне многих мотиваций лежит получение выгоды и желание комфорта, побуждающее личность идти путём наименьшего сопротивления к данным состояниям.
Все образующиеся в нашем мозге нейросети не обособлены, а тесно и сложно взаимоувязаны между собой, и именно эти взаимосвязи образуют сложные идеи, глубокие переживания, воспоминания из жизни, образы давно испытанных эмоций. Человеческий мозг взаимодействует с миллионами бит информации каждую секунду, но сознательно мы способны воспринимать только небольшую часть из них.
Как уже говорилось выше, если часто реагировать одним образом, активизируется определенная нейронная сеть и соответствующее поведение превращается в устойчивую привычку. Чем чаще используется сеть, тем прочнее она становится и тем проще получить к ней доступ.
Также возможен и обратный процесс: устойчивые связи между нейронами, не используемыми вместе, ослабевают. Всякий раз, когда мы прекращаем или предотвращаем действие, ментальный процесс, оформленный в нейросеть, соединенные между собой нервные клетки и группы клеток ослабляют свою связь. При этом тончайшие дендриты, отходящие от каждого нейрона и связывающие его с другими нейронами, освобождаются для связи с другими. Этот механизм обусловлен нейропластичностью (нейрогенезом) - в зависимости от стимуляции нейронов, одни связи становятся прочнее и эффективнее, а другие ослабевают, высвобождая потенциал для формирования новых.
Когда мозг привыкает не думать, не решать сложных задач и не преодолевать сложности, не трансформировать внутренние и внешние конфликты, то происходит торможение развития и постепенная деградация личности, потому что электрические импульсы задействуют уже знакомые им пути, не прокладывая новых.
В цикле книг по ииссиидиологии для развития нейронных цепочек в сторону высокочувственного и высокоинтеллектуального потенциала личности предлагается модель интеллектуально-альтруистичного развития и образа жизни. Это позволяет нам в большей степени задействовать Человеческие отделы мозга и гармонизировать активность эгоистично-импульсивных выборов и программ. Ииссиидиология и Центры Интеллекта и Альтруизма, создаваемые для её практического применения, направлены на радикальное позитивное человеческое переформатирование и трансформацию нейронных сетей, соответствующих модели нашего восприятия себя и мира вокруг нас [8].
С точки зрения физиологии, привычки есть не что иное, как образование в мозговых структурах устойчивых нервных взаимосвязей, отличающихся повышенной готовностью к функционированию по сравнению с другими цепочками нейронного реагирования. Чем больше мы повторяем какие-либо действия, мысли, слова, тем более активными и автоматическими становятся соответствующие нейронные пути.
Разные отделы полушарий мозга играют в этом процессе свою необходимую роль. Когда их деятельность синхронизирована между собой, то передача информации между нейронами становится оптимальной.
Известно, что мозг состоит из эмбриональной ткани, поэтому он всегда открыт для развития, обучения и перемен. Учёные убеждены, что наш мозг способен простой мыслью, воображением, визуализацией, изменять структуру и функцию серого вещества, и это может происходить даже без специальных, внешних воздействий, а под влиянием лишь тех мыслей, которыми он наполнен. Всё вышесказанное подводит к пониманию того, что каждому, кто задумывается о качестве своего мышления и привычек, требуется фундаментальное изменение накопленных убеждений – преодоление и смену инстинктивной генетической программы и прежнего воспитания на истинно человеческие представления, основанные на высокоинтеллектуальных и высокочувственных представлениях любого аспекта жизни.
Читайте также:
