
Нервная регуляция тонуса мышц
В большинстве движений участвует множество мышц, причем сокращение и расслабление различных групп мышц происходит в определенном порядке и с определенной силой. Такая согласованность движений называется координацией движений. Она осуществляется нервной системой. Скелетные мышцы иннервируются соматическим отделом нервной системы. К каждой мышце подходит один или несколько нервов, проникающих в ее толщу и разветвляющихся на множество мелких отростков, которые достигают мышечных волокон. Посредством нервов осуществляется связь мышц с ЦНС, которая регулирует любые двигательные акты (ходьба, бег, пищевые движения и т. д.) и длительное напряжение мышц — тонус, поддерживающий определенное положение тела в пространстве. Деятельность мышц носит рефлекторный характер. Мышечный рефлекс может запускаться с раздражения рецепторов, находящихся в самой мышце или в сухожилиях, либо с раздражения зрительных, слуховых, обонятельных, осязательных рецепторов.
В регуляции безусловно-рефлекторных движений принимает участие мозжечок. Он осуществляет координацию движения, регуляцию мышечного тонуса, способствует поддержанию равновесия и позы тела. При поражении мозжечка его регуляторные двигательные функции нарушаются.
Сокращаясь, мышца действует на кость как на рычаг и производит механическую работу. На осуществление работы мышцы затрачивается энергия, которая образуется в результате распада и окисления органических веществ, поступивших в мышечную клетку. Основным источником энергии является АТФ. Кровь доставляет мышцам питательные вещества и кислород и уносит образующиеся продукты диссимиляции (углекислый газ и др.). При длительной работе наступает утомление и снижение работоспособности мышцы, возникающее из-за несоответствия между ее кровоснабжением и возросшими потребностями в питательных веществах и кислороде. Систематическая мышечная работа усиливает кровоснабжение мыши и костей, к которым они прикрепляются. Это приводит к увеличению мышечной массы и усиленному росту костей. Сильные мышцы легко справляются с поддержанием туловища в нужном положении, противостоят развитию сутулости, искривлению позвоночника.
ЗАКЛЮЧЕНИЕ
Значение скелета очень велико.Костная система выполняет ряд функций, имеющих или преимущественно механическое, или преимущественно биологическое значение. Рассмотрим функции, имеющие преимущественно механическое значение. Для всех позвоночных характерен внутренний скелет, хотя среди них встречаются виды, которые, наряду с внутренним скелетом, имеют еще и более или менее развитый наружный скелет, возникающий в коже (костная чешуя в коже рыб). В начале своего появления твердый скелет служил для защиты организма от вредных внешних влияний (наружный скелет беспозвоночных). С развитием внутреннего скелета у позвоночных он сначала стал опорой и каркасом для мягких тканей. Отдельные части скелета превратились в рычаги, приводимые в движение мышцами, вследствие чего скелет приобрел локомоторную функцию. В итоге механические функции скелета проявляются в его способности осуществлять защиту, опору и движение.
Опора достигается прикреплением мягких тканей и органов к различным частям скелета. Движение возможно благодаря тому, что кости являются длинными и короткими рычагами, соединенными подвижными сочленениями и приводимыми в движение мышцами, управляемыми нервной системой.
Наконец, защита осуществляется путем образования из отдельных
костей костного канала — позвоночного, защищающего спинной мозг, костной коробки — черепа, защищающего головной мозг; костной клетки — грудной, защищающей жизненно важные органы грудной полости (сердце, легкие, печень, желудок, селезенку, частично почки и др., то есть важнейшие органы разных систем); костного вместилища — таза, защищающего важные для продолжения вида органы размножения, выделения.
Биологическая функция костной системы связана с участием скелета в обмене веществ, особенно в минеральном обмене (скелет является депо минеральных солей — фосфора, кальция, железа и др.). Это важно учитывать для понимания болезней обмена (рахит и др.) и для диагностики с помощью лучи-стой энергии (рентгеновские лучи, радионуклиды). Кроме того, скелет выполняет еще кроветворную функцию. При этом кость является не просто защитным футляром для костного мозга, а последний составляет органическую часть ее. Определенное развитие и деятельность костного мозга отражаются на строении костного вещества, и, наоборот, механические факторы сказываются на функции кроветворения: усиленное движение способствует кроветворению, поэтому при разработке физических упражнений необходимо учитывать единство всех функций скелета.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Тонус исследуют после поперечной перерезки спинного мозга. Сразу после перерезки в эксперименте или после травматического повреждения его у человека наблюдаются мышечная атония и отсутствие рефлексов (спинальный шок). Главная причина спинального шока - выключение влияния на спинной мозг вышележащих отделов ЦНС (повторная перерезка спинного мозга ниже первой после исчезновения шока повторно шок не вызывает). Спинальный шок у лягушек длится несколько минут, у собак - несколько дней, у человека - около 2 мес. После исчезновения спинального шока тонус мышц, иннервируемых посредством сегментов спинного мозга, которые находятся ниже перерезки (повреждения), резко повышается. Сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются. Механизм повышения тонуса мышц нижних конечностей в случае повреждения спинного мозга (после окончания спинального шока) отражает рис. 5.8.
Гипертонус имеет рефлекторную природу - он развивается вследствие афферентной импульсации от мышечных рецепторов. Деафферентация, например, нижней конечности у лягушки ведет к исчезновению тонуса у нее (опыт Бронжеста).
Афферентную импульсацшо от мышечных рецепторов по задним корешкам спинного мозга, которая ведет к возбуждению а-мо-тонейронов спинного мозга и сокращению скелетных мышц, обеспечивают три фактора:
• растяжение и возбуждение мышечных рецепторов под влиянием сил гравитации конечности;
• спонтанная активность мышечных рецепторов;
• спонтанная активность у-мотонейронов, которая реализуется следующим образом. Возбуждение у-мотонейронов вызывает возбуждение и сокращение иннервируемых ими ин-трафузальных мышечных волокон, в результате чего увеличи-
вается натяжение мышечного рецептора, поскольку концы его зафиксированы на скелетной мышце. Вследствие натяжения мышечного веретена раздражаются и возбуждаются его афферентные окончания(проприорецепто-ры), импульсы от которых поступают к а-мотонейро-нам, возбуждая их. В свою очередь а-мотонейроны посылают импульсы к скелетной мышце и вызывают ее постоянное (тоническое) сокращение.
Мы рассмотрели рефлекторную и проводниковую функции изолированного спинного мозга. В нормальных условиях деятельность спинного мозга контролируется вышележащими отделами ЦНС посредством импульсации ко всем его нервным элементам.
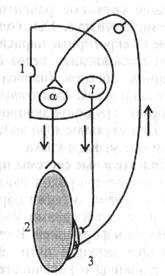 | Рис. 5.8. Схема механизма гипертонуса мышц нижних конечностей в случае повреждения спинного мозга. 1 — полусегмент спинного мозга; 2 - скелетная мышца; 3 – мышечное веретено; Т - афферентный путь; I - эфферентные пути; а - мотонейрон, иннервирующий скелетную мышцу; у - мотонейрон, иннервирующий интрафузальныс мышечные волокна (проприорецептора) 5.3. ДВИГАТЕЛЬНЫЕ СИСТЕМЫ СТВОЛА МОЗГА В этот отдел мозга физиологи включают продолговатый мозг, мост и средний мозг. А. В стволе мозга находятся ядра Ш-Х11 пар черепных нервов. Ядра глазодвигательного нерва (n/ oculomotorius, III); ядра блокового нерва (пn/ trochlearis, IV); ядра отводящего нерва (n/ abductns, VI) управляют движением глаз в различных направлениях. Ядра тройничного нерва (n/ trigeminus, V) иннервируют жевательную мускулатуру и вызывают движение нижней челюсти. Двигательное ядро лицевого нерва (n/ facialis, |
VII) ызывает сокращение мимической мускулатуры. К ядрам преддверно-улиткового нерва (n. vestibulocochlearis, VIII) относятся: вестибулярные ядра, которые получают импульсы от вестибулорецепторов и участвуют в регуляции позы и равновесия тела; улитковые ядра, иннервирующие слуховые рецепторы. Ядра языкоглоточного нерва (n. glossopharyngeus, IX), содержащего двигательные, чувствительные и секреторные парасимпатические волокна.
Ядра блуждающего нерва (n. vagus, X) регулируют функции внутренних органов (парасимпатический нерв). Вожбуждение двигательного ядра добавочного нерва (п. accessories, XI) вызывает сокращение грудино-ключично-сосцевидной и трапециевидной мышц. Двигательное ядро подъязычного нерва (n. hypoglossus, XII) иннервирует мышцы языка.
Б. Двигательные системы продолговатого мозга и мостя. Из перечисленных ядер продолговатый мозг включает ядра 1Х-ХII пар черепных нервов, а также образования ретикулярной формации. Мост представлен ядрами V-VIII пар черепных нервов, а также ретикулярной формацией. В стволе мозга проходят восходящие и нисходящие нервные пути. Важную роль в регуляции тонуса мышц туловища и конечностей играют вестибулярные ядра, основным из которых является ядро Дейтерса.
1. Рефлексы, дуги которых замыкаются в продолговатом мозге и мосту, можно объединить в три основные группы:
• жизненно важные вегетативные: дыхания, сердечно-сосудистой системы, пищеварительной системы, глотания;
• защитные рефлексы: чиханья, кашля, мигания, слезоотделения, рвотный;
• соматические рефлексы, участвующие в регуляции тонуса и двигательной активности мышц туловища, конечностей, шеи, лица.
Рефлексы любого отдела ЦНС изучают после отделения вышележащих отделов мозга. У бульварного животного, у которого произведена перерезка ствола мозга между мостом и средним мозгом, сохраняются все рефлексы, в возникновении которых играют роль спинной мозг, а также указанные черепные нервы и их ядра. Особо важное значение в регуляции тонуса мышц туловища и конечностей имеет ядро Дейтерса. С его помощью осуществляется, в частности, лабиринтный рефлекс позы - повышение тонуса мышц-разгибателей при положении животного вверх ногами. Рефлекс в натуральных условиях направлен на сохранение позы. Он возникает с рецепторов вестибулярного аппарата (при его разрушении рефлекс исчезает). Проприорецепторы мышц шеи при изучении этого рефлекса должны быть выключены, что достигается путем гипсования шеи (исключаются сгибание шеи и раздражение проприорецепторов мышц шеи).
2. Общее состояние бульварного животного характеризуется тем, что произвольные движения с помощью мышц, иннервируемых отделами ЦНС, расположенными ниже перерезки, невозможны, тонус
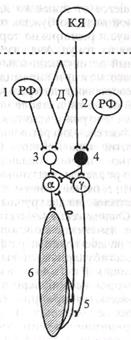 | Рис. 5.9. Механизмы регуляции тонуса мышц и возникновения децеребрационной ригидности при перерезке ствола мозга между средним мозгом и мостом (пунктирная линия). КЯ - красное ядро; РФ - ретикулярная формация моста (1) и продолговатого мозга (2); Д - ядро Дейтерса. Нейроны спинного мозга: 3 — возбуждающий, 4 - тормозной, а — мотопейрон, у - мотонейрон; 5 - проприорецептор (мышечное веретено); 6 - мышца-разгибатель Волокна ядра Дейтерса оказывают возбуждающее влияние на спинальные а- и у-мотонейроны мышц - разгибателей конечностей, туловища, шеи, а также реципрокное тормозное влияние на мото - нейроны сгибателей. Аналогично на мышечный тонус влияет ретикулярная формация моста - медиальный ретикулоспинальный тракт. Вместе они составляют медиальную нисходящую разгибательную систему. В. Двигательные системы среднего мозга включают красные ядра, передние и задние холмики четверохолмия: передние - первичные зрительные центры вместе с латеральными коленчатыми телами; задние — первичные слуховые центры вместе с медиальными коленчатыми телами, ядра III и IV пары черепных нервов, черную субстанцию, ретикулярную формацию и голубое пятно. Ниже приведена характеристика рефлексов. |
1. Характеристика рефлексов, осуществляемых средним мозгом (мезенцефальное животное). Они более совершенны, их набор значительно больше, тонус мышц близок к нормальному, поскольку сохраняется тормозящее влияние красного ядра на а- и у-мотонейроны спинного мозга, наблюдается незначительная ригидность (см. рис. 5.9). Руброспиналънъш тракт (тракт Монакова), которыйначинается от красного ядра покрышки среднего мозга и идет в спинной мозг, возбуждая а- и у - мотонейроны сгибателей, повышает их тонус и реципрокно тормозит мотонейроны мышц - разгибателей, снижая их тонус. Аналогично на мышечный тонус действуют латеральный ретикулоспинальный (от ретикулярной формации продолговатого мозга) и пирамидный кортикоспинальный тракты. Вместе они составляют латеральную нисходящую сгибательную систему. Таким образом, в стволе мозга имеются четыре главных парных моторных центров и трактов, регулирующих тонус мышц туловища и конечностей, - два ретикулоспинальных пути, руброспинальный путь и волокна ядра Дейтерса (см. рис, 5.9). Рефлексы мезенцефального животного изучил голландский физиолог Р.Магнус (1924). Соматические рефлексы, связанные с изменением положения тела (точнее - головы) и перемещением его в пространстве, Р. Магнус разделил соответственно на две группы - статические и статокинетические.
2. Статические рефлексы (перераспределение мышечного тонуса при изменении положения головы в покое) в свою очередь делятся на две группы: рефлексы позы (шейные и лабиринтные вестибулярные) и выпрямительные (выпрямление головы и туловища). Рефлекс выпрямления головы возникаете рецепторов вестибулярного аппарата, кожи и сетчатки. При этом возбуждаются проприорецепторы мышц шеи, и начинается выпрямление туловища. В осуществлении выпрямительных рефлексов принимают участие также и рецепторы кожи — дублирование рефлексогенных зон. При выключении рецепторов кожи или вестибулярного аппарата выпрямительные рефлексы не нарушаются. При выключении обеих названных рефлексогенных зон выпрямительные рефлексы не возникают. Для осуществления выпрямительных рефлексов, т.е. восстановления нарушенной позы, необходимо обязательное участие красных ядер, повреждение которых, как показано в эксперименте на животных, предотвращает возникновение выпрямительных рефлексов. Однако у человека и обезьян выпрямительные рефлексы осуществляются при обязательном участии коры большого мозга; при ее выключении выпрямительные рефлексы не возникают. При изменении положения головы появляются также рефлексы компенсаторного положения глаз, обеспечивающие правильное видение при различных положениях головы относительно тела и в пространстве. Например, если повернуть голову вправо, глазное яблоко поворачивается влево, как бы стремясь удержать первоначальное поле зрения.
3. Статокинетические рефлексы возникают при ускорениях прямолинейного и вращательного движений организма. Перераспределение тонуса направлено на преодоление действующих на человека ускорений и сохранение нормальной позы. Для их осуществления необходимо обязательное участие структур среднего мозга.
Примером рефлекса прямолинейного ускорения является лифтный рефлекс. В начале быстрого подъема лифта (или при остановке движущегося вниз лифта) в нижних конечностях повышается тонус мышц-разгибателей. В начале опускания лифта (или при остановке движущегося вверх лифта) повышается тонус мышц-сгибателей. О рефлекторном характере изменения тонуса мышц конечностей при лифтном рефлексе свидетельствует его отсутствие у животных при выключении вестибулярных рецепторов разрушением лабиринта.
Статокинетические рефлексы вращения включают мускулатуру тела и глазные мышцы. Движение глазных яблок (глазной нистагм) при этом способствует сохранению зрительной ориентации. Нистагм имеет две фазы. При ускорении вращения сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению. После этого они быстро отклоняются обратно (по ходу вращения). Медленное отклонение вызывается с рецепторов полукружных каналов, быстрый компонент связан с влиянием корковых центров (при наркозе он исчезает). При замедлении вращения, напротив, медленное движение глаз направлено в сторону вращения, а быстрое - против направления вращения. Движение глаз при нистагме (вправо-влево, вверх-вниз, круговые движения) зависит от возбуждения рецепторов соответственно горизонтального, сагиттального и фронтального полукружных каналов.
Наряду с регуляцией мышечного тонуса, позы и равновесия структуры ствола головного мозга участвуют в контроле спинального автоматизма шагания и, следовательно, в контроле локомоции (например, ходьбы). Важное значение в осуществлении этой функции придают клинообразному ядру в задненижнем отделе среднего мозга, который обозначают как локомоторную область среднего мозга, имеющую эфферентный выход к ядрам моста.
Произвольная локомоция без участия корковых нейронов невозможна.
В условиях эксперимента при сохранении среднего мозга у животного возникает сторожевой рефлексов - повороты головы, глаз, ушей, туловища в сторону звука или света (настора-живание), при этом повышается тонус мышц-сгибателей (животное как бы готовится к прыжку). В случае действия светового раздражителя наблюдаются зрачковый рефлекс (сужение зрачка), аккомодация, дивергенция и конвергенция зрительных осей, чтоосуществляется с помощью передних холмиков четверохолмия (первичный зрительный центр) и III и IV пар черепных нервов. Задние холмики четверохолмия - первичные слуховые центры.
4. Функции голубого пятна. Оно расположено в среднем мозге, оказывает моделирующее влияние на двигательные механизмы спинного мозга. Аксоны норадренергических нейронов голубого пятна идут в переднем и боковом канатиках к а-мотонейронам передних рогов спинного мозга. Полагают, что норадреналин является тормозным медиатором для н-мотонейронов. Импульсная активность нейронов голубого пятна повышена в фазе быстрого сна, что выключает мышечный тонус и фазические движения мускулатуры туловища, конечностей. При одностороннем разрушении голубого пятна возникают вращательные движения в сторону, противоположную разрушению, что, как предполагают, связано с нарушением функциональных связей голубого пятна и базальных ганглиев. В электрофизиологических экспериментах было показано, что норадренергические нейроны голубого пятна оказывают ингибирующее действие почти на все иннервируемые ими образования ЦНС. Поскольку эти нейроны возбуждаются во время различных стрессовых воздействий, считают, что их ингибирующий эффект играет двоякую роль: 1) под их влиянием при стрессе нервная система предохраняется от перевозбуждения: 2) возбудимость клеток нервной системы поддерживается на постоянном уровне с целью оптимальной передачи сигналов.
5. Функции черной субстанции. Она участвует в регуляции мышечного тонуса, координации жевания и глотания, мелких движений конечностей (например, печатание на машинке, письмо), в осуществлении эмоций. В случае повреждения черной субстанции наблюдается бедность движений.
Во всех двигательных реакциях и регуляции мышечного тонуса весьма важную роль играет ретикулярная формация, основная масса нейронов которой находится в стволе мозга.
Мышечная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы тела. Эти процессы регулируются различными отделами ЦНС.
Роль спинного мозга
Мышечный тонус является по своей природе рефлекторным актом. Для его возникновения достаточна рефлекторная деятельность спинного мозга. При длительном растяжении мышц в поле силы тяжести возникает постоянное раздражение их проприорецепторов, потоки импульсов от которых проходят по толстым афферентным волокнам в спинной мозг, где передаются непосредственно (безучастия вставочных нейронов) на альфа-мотонейроны передних рогов и вызывают тоническое напряжение мышц. Такие двухнейронные (или моносинаптические) рефлекторные дуги лежат в основе тонических сухожильных (с рецепторов сухожилий) и миотатических рефлексов на растяжение (с рецепторов мышечных веретен). Это рефлексы активного противодействия мышцы ее растяжению. В произвольной двигательной деятельности человека иногда требуется подавление этих рефлексов, например, при выполнении шпагата.
Степеньтонического напряжения мышцы зависит от частоты импульсов, посылаемых к ней альфа-мотонейронами. Однако, потоки этих импульсов могут регулироваться вышележащими этажами нервной системы, в частности, неспецифическими отделами ствола мозга с помощью так называемой гамма — регуляции. Разряды гамма-мотонейронов спинного мозга под влиянием ретикулярной формации повышают чувствительность рецепторов мышечных веретен. В результате при той же длине мышцы увеличивается поток импульсов от рецепторов к альфа-мотонейронам и далее к мышце, повышая ее тонус.
Роль коры головного мозга, мозжечка и ствола мозга
В регуляции тонуса участвует также медленная часть пирамидной системы и различные структуры экстрапирамидной системы (подкорковые ядра, красные ядра и черная субстанция среднего мозга, мозжечок, ретикулярная формация ствола мозга, вестибулярные ядра продолговатого мозга).
Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел ретикулярной формации среднего мозга, а угнетение — тормозящий отдел продолговатого мозга. В отличие от нее специфическая система (моторные центры коры больших полушарий и ствола мозга) действует избирательно, т. е. на отдельные группы мышц-сгибателей или разгибателей. Усиление тонуса мышц-сгибателей вызывают корковые влияния, передающиеся непосредственно к нейронам спинного мозга но корково-спинномозговой (пирамидной) системе, а также через красные ядра (по корково-красноядерно-спинномоз-говой системе) и частично через ретикулярную формацию (по ретикуло-спинномозговой системе). В противоположность им, влияния, передающиеся через вестибулярные ядра продолговатого мозга к вставочным и двигательным нейронам передних рогов спинного мозга (по вестибуло-спинномозговым путям), вызывают тоническое повышение возбудимости мотонейронов мышц-разгибатслей. что обеспечивает повышение тонуса этих мышц.
Мозжечок формирует правильное распределение тонуса скелетных мышц: через красные ядра среднего мозга он повышает тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга — усиливает тонус мышц-разгибателей. В поддержании позы и равновесия тела, регуляции тонуса мышц основное значение имеет медиальная продольная зона мозжечка — кора червя. При мозжечковых расстройствах падает тоническое напряжение мышц (атония) и вследствие ненормального распределения тонуса мышц конечностей возникает нарушение походки (атаксия).
Бледное ядро угнетает тонус мышц, а полосатое тело снижаем его угнетающее действие.
Высший контроль тонической активности мышц осуществляет кора больших полушарий, в частности ее моторные, премоторные и лобные области. С ее участием происходит выбор наиболее целесообразной для данного момента позы тела, обеспечивается ее соответствие двигательной задаче. Непосредственное отношение к регуляции тонуса мышц имеют медленные пирамидные нейроны положения. Корковые влияния на тонические реакции мышц передаются через медленную часть пирамидного тракта и через экстрапирамидную систему.
Рефлексы поддержания позы (установочные)
Специальная группа рефлексов способствует сохранению позы — это так называемыеустановочные рефлексы. К ним относятся статические и стато-кинетические рефлексы, в осуществлении которых большое значение имеют продолговатый и средний мозг.
Статические рефлексы возникают при изменении положения тела или его частей в пространстве: 1) при изменениях положения головы в пространстве — лабиринтные рефлексы, возникающие при раздражении рецепторов вестибулярного аппарата. 2) шейные рефлексы — возникающие с проприорецепторов мышц шеи при изменении положения головы по отношению к туловищу, и 3) выпрямительные рефлексы — с рецепторов кожи, вестибулярного аппарата и сетчатки глаза. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед — тонус мышц-сгибателей (лабиринтный рефлекс). С помощью выпрямительного рефлекса происходят последовательные сокращения мышц шеи и туловища, а затем и конечностей. Этот рефлекс обеспечивает вертикальное положение тела теменем кверху. У человека он проявляется, например, при нырянии.
Стато-кинетические рефлексыкомпенсируют отклонения тела при ускорении ши замедлении прямолинейного движения (лифтный рефлекс), а также при вращениях(отклонения головы, тела и глаз в сторону, противоположную движению). Перемещение глаз со скоростью вращения тела, но в противоположную сторону, и быстрое их возвращение в исходное положение — нистагм глаз — обеспечивает сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
Роль различных отделов ЦНС в регуляции движений
Спинной мозг обеспечивает протекание многих элементарных двигательных рефлексов, включение которых в сложные двигательные акты и регуляция по мощности, пространственной ориентации и моменту включения осуществляется вышележащими отделами головного мозга под контролем коры больших полушарий.
Роль спинного мозга и подкорковых отделов ЦНС в регуляции движений
Спинной мозг осуществляет ряд элементарных двигательных рефлексов:рефлексы на растяжение (миотатические и сухожильные рефлексы, например, коленный рефлекс), кожные сгибательные рефлексы (например, защитный рефлекс отдергивания конечности при уколах, ожогах), разгибательные рефлексы (рефлекс отталкивания от опоры, лежащий в основе стояния, ходьбы, бега), перекрестные рефлексы и др.
Элементарные двигательные рефлексы включаются в более сложные двигательные акты — регуляцию деятельности мышц-антагонистов, ритмических и шагательных рефлексов, лежащих в основе локомоций и других движений.
Для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, по и одновременное расслабление мышц-разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей — торможение. При разгибании сустава, наоборот, тормозятся центры сгибателей и возбуждаются центры разгибателей. Такие координационные взаимоотношения между спинальными моторными центрам и названы реципрокной (взаимосочетанной) иннервацией мышц-антагонистов. Однако реципторные отношения между центрами мышц-антагонистов в необходимых ситуациях (например, при фиксации суставов, при точностных движениях) могут сменяться одновременным их возбуждением.
Нейроны промежуточной продольной зоны коры мозжечка согласуют позные реакции с движениями. Они выполняют также точные расчеты по ходу движений, необходимые для коррекции ошибок и адаптации моторных программ к текущей ситуации. Программирование каждого последующего шага осуществляется ими на основе анализа предыдущего. Кроме того производится согласование движений рук и ног, и особенно — регуляция активности мышц-разгибателей, обеспечивающих опорную фазу движения. Значение мозжечка в четкомподдержании темпа ритмических движений объясняют геометрически правильным чередованием рядов эфферентных клеток Пуркинье и походящих к ним афферентных волокон.
К управлению ритмическими движениями непосредственное отношение имеют активирующие и угнетающие отделы ретикулярной формации, влияющие на силу и темп сокращения мышц, а также подкорковые ядра, которые организуют автоматическое их протекание и содружественные движения конечностей. Включение древних форм ритмических движений (циклоидных) в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи. То же самое происходит при освоении акта ходьбы — с переходом от отдельных шагов к ритмической походке. Плавность ритмических движений, четкое чередование реципрокных сокращений мышц обеспечивают премоторные отделы коры.
Роль различных отделов коры больших полушарий
В высшей регуляции произвольных движений важнейшая роль принадлежитпередне-лобным областям (передним третичным полям). Здесь помимо обычных вертикальных колонок нейронов существует принципиально новый тип функциональной единицы — в форме замкнутого нейронного кольца. Циркуляция импульсов в этой замкнутой системе обеспечивает кратковременную память. Она сохраняет в коре возбуждение между временем прихода сенсорных сигналов и формированием ответной эфферентной команды. Такой механизм служит основой сенсомоторной интеграции при программировании движений, при осуществлении зрительно-двигательных реакций.
Функцией передне-лобной (третичной) области коры является сознательная оценка текущей ситуации и предвидение возможного будущего, выработка цели и задачи поведения, программирование произвольных движений, их контроль и коррекция. Соответствие выполняемых действий поставленным задачам придает движениям человека определенную целесообразность и осмысленность. При поражении лобных долей движения человека становятся бессмысленными.
Речевая регуляция движений
Спецификой регуляции движений у человека является то, что они подчиненыречевым воздействиям, т. е. могут программироваться лобными долями в ответ на поступающие извне словесные сигналы, а также благодаря участию внешней или внутренней речи (мышления) самого человека. В этой функции принимают участие расположенные в левом полушарии человека сенсорный центр речи Вернике и моторный центр речи — центр Брока. Считают, что афферентная импульсация от речевой мускулатуры является важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц, а формирующиеся на речевой основе избирательные связи в коре облегчают составление моторных программ.
Эта управляющая система еще не развита у ребенка 2-3 лет. Она появляется лишь к 3-4 годам. Внешняя речь, сменяясь постепенно шепотом и переходя затем во внутреннюю речь, становится важным регулятором моторных действий взрослого человека.
Нисходящие моторные системы
Высшие отделы головного мозга осуществляют свои влияния па деятельность нижележащих отделов, в том числе спинного мозга, через нисходящие пути, которые группируют обычно в две основные нисходящие системы — пирамидную и экстрапирамидную.
В настоящее время предлагают подразделять основные нисходящие пути, исходя из расположения нервных окончаний в спинном мозге и функциональных различий, на следующие 2 системы: более молодую латеральную, волокна которой оканчиваются в боковых (латеральных) частях спинного мозга и связанную преимущественно с мускулатурой дистальных звеньев конечностей (сюда относят корково-спинномозговую и красноядерно-спинномозговую системы), и древнююмедиальную, волокна которой оканчиваются во внутренних (медиальных) частях белого вещества, связанную главным образом с мускулатурой туловища и проксимальных звеньев конечностей, состоящую из вестибуло-спинномозговой и ретикуло-спинномозговой систем.
Пирамидная системавыполняет 3 основные функции:
посылает мотонейропам спинного мозга импульсы — команды к движениям (пусковые влияния);
изменяет проведение нервных импульсов во вставочных спинальных нейронах, облегчая протекание нужных в данный момент спинномозговых рефлексов;
осуществляет контроль за потоками афферентных сигналов в нервные центры, выключая постороннюю информацию и обеспечивая обратные связи от работающих мышц.
Волокна пирамидной системы вызывают преимущественно возбуждение мотонейронов мышц-сгибателей, особенно влияя на отдельные мышцы и даже части мыши верхних конечностей, в частности на мышцы пальцев рук.

Рис. 1. Схема основных нисходящих путей регуляции двигательной деятельности
1 — быстрая подсистема и 2 — медленная подсистема корково-спиномозго-вого пути (пирамидного тракта); 3 — корково-красноядерно-спиномозговой путь. Латеральная система — I, 2, 3. Медиальная система — 4, 5. М — мотонейрон спинного мозга, получающий фазные (Фазн.) и тонические (Тонич.) возбуждающие (+) и тормозящие (-) влияния.
Читайте также:
