
Нервная система у беспозвоночных животных в процессе эволюции
3. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ФИЛОГЕНЕЗЕ
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. У одного и того же типа животных нервные клетки могут одновременно и независимо происходить из трех разных зародышевых листков. Полигенез нервных клеток беспозвоночных является основой разнообразия медиаторных механизмов их нервной системы.
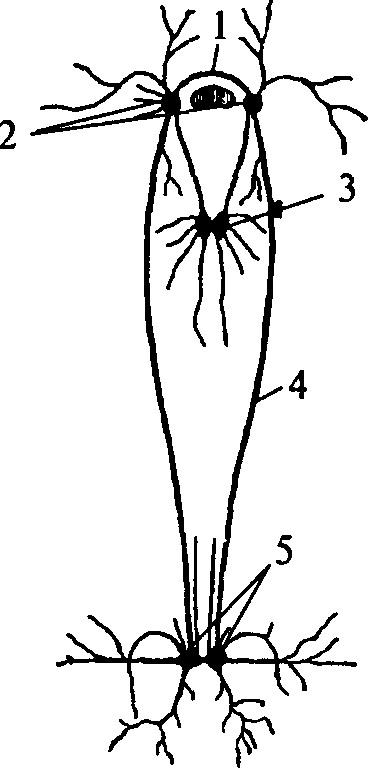
Нервная система впервые появляется у кишечнополостных животных. Кишечнополостные — это двухслойные животные. Их тело представляет собой полый мешок, внутренняя полость которого является пищеварительной полостью. Нервная система кишечнополостных принадлежит к диффузному типу. Каждая нервная клетка в ней длинными отростками соединена с несколькими соседними, образуя нервную сеть. Нервные клетки кишечнополостных не имеют специализированных поляризованных отростков. Их отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками диффузной нервной системы бывают нескольких типов. Существуют плазматические контакты, обеспечивающие непрерывность сети (анастомозы). Появляются и щелевидные контакты между отростками нервных клеток, подобные синапсам. Причем среди них существуют контакты, в которых синаптические пузырьки располагаются по обе стороны контакта — так называемые симметричные синапсы, а есть и несимметричные синапсы: в них везикулы располагаются только с одной стороны щели.
Нервные клетки типичного кишечнополостного животного гидры равномерно распределены по поверхности тела, образуя некоторые скопления в районе ротового отверстия и подошвы (рис. 8). Диффузная нервная сеть проводит возбуждение во всех направлениях. При этом волну распространяющегося возбуждения сопровождает волна мышечного сокращения.
Рис. 8. Схема строения диффузной нервной системы кишечнополостного животного:
1 — ротовое отверстие; 2 — щупальце; 3 — подошва
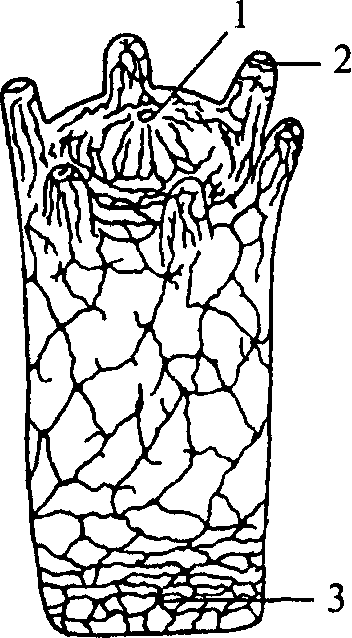
Рис. 9. Схема строения диффузностволовой нервной системы турбеллярии:
1 — нервный узел; 2 — глотка; 3 — брюшной продольный ствол; 4 — боковой нервный ствол
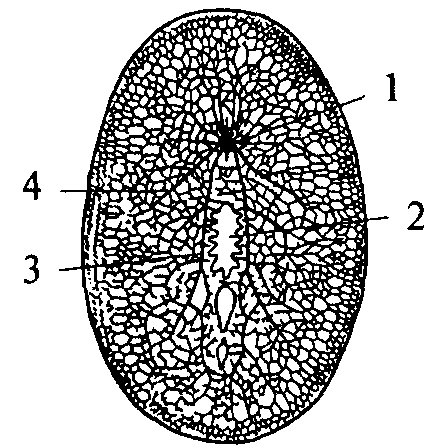
Следующим этапом развития беспозвоночных является появление трехслойных животных — плоских червей. Подобно кишечнополостным они имеют кишечную полость, сообщающуюся с внешней средой ротовым отверстием. Однако у них появляется третий зародышевый слой — мезодерма и двусторонний тип симметрии. Нервная система низших плоских червей принадлежит диффузному типу. Однако из диффузной сети уже обособляются несколько нервных стволов (рис. 9, 3, 4).
У свободно живущих плоских червей нервный аппарат приобретает черты централизации. Нервные элементы собираются в несколько продольных стволов (рис. 10, 4, 5) (для самых высокоорганизованных животных характерно наличие двух стволов), которые соединяются между собой поперечными волокнами (комиссурами) (рис. 10, 6). Упорядоченная таким образом нервная система называется ортогоном. Стволы ортогона представляют собой совокупность нервных клеток и их отростков (рис. 10).
1 — щупальцевидный вырост; 2 — нерв, иннервирующий вырост; 3 — мозговой ганглий; 4 — боковой продольный нервный ствол; 5 — брюшной продольный нервный ствол; 6 — комиссура
ганглия появляются длинные отростки, идущие в продольные стволы ортогона (рис. 10, 4, 5).
Таким образом, ортогон представляет собой первый шаг к централизации нервного аппарата и его цефализации (появлению мозга). Централизация и цефализация являются результатом развития сенсорных (чувствительных) структур.
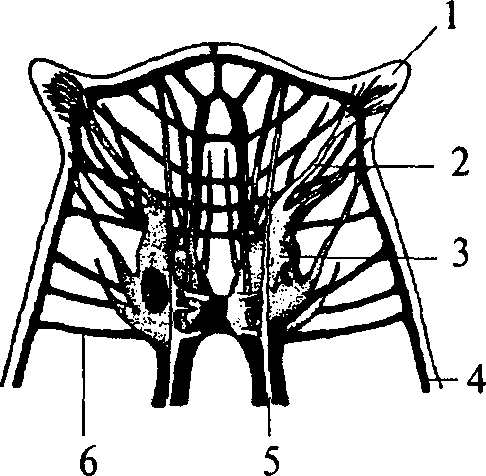
Следующим этапом развития беспозвоночных животных является появление сегментированных животных — кольчатых червей. Их тело метамерно, т.е. состоит из сегментов. Структурной основой нервной системы кольчатых червей является ганглий — парное скопление нервных клеток, расположенных по одному в каждом сегменте. Нервные клетки в ганглии размещаются по периферии. Центральную его часть занимает нейропиль — переплетение отростков нервных клеток и глиальные клетки. Ганглий расположен на брюшной стороне сегмента под кишечной трубкой. Он посылает свои чувствительные и двигательные волокна в свой сегмент и в два соседних. Таким образом, каждый ганглий имеет три пары боковых нервов, каждый из которых является смешанным и иннервирует свой сегмент. Приходящие с периферии чувствительные волокна попадают в ганглий через вентральные корешки нервов. Двигательные волокна выходят из ганглия по дорсальным корешкам нервов. Соответственно этому чувствительные нейроны расположены в вентральной части ганглия, а двигательные — в дорсальной. Кроме того, в ганглии есть мелкие клетки, иннервирующие внутренние органы (вегетативные элементы), они расположены латерально — между чувствительными и двигательными нейронами. Среди нейронов чувствительной, двигательной или ассоциативной зон ганглиев кольчатых червей не обнаружено группирования элементов, нейроны распределены диффузно, т.е. не образуют центров.
Ганглии кольчатых червей соединены между собой в цепочку. Каждый последующий ганглий связан с предыдущим при помощи
1 — надглоточный нервный ганглий;
2 — подглоточный нервный ганглий;
3 — сложный слившийся ганглий грудного сегмента; 4 — брюшной ганглий; 5 — периферический нерв; 6 — коннектива
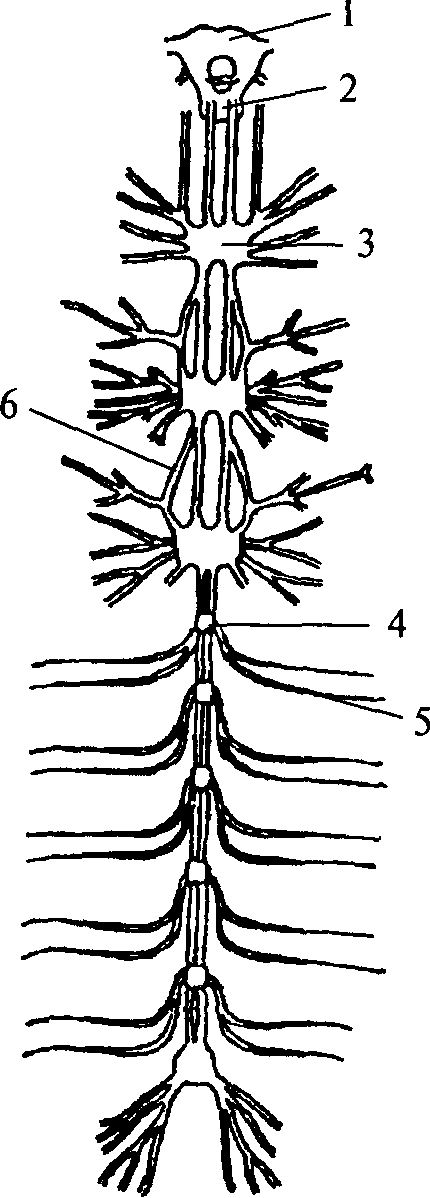
нервных стволов, которые называются коннективами. На переднем конце тела кольчатых червей два слившихся ганглия образуют крупный подглоточный нервный узел. Коннективы от подглоточного нервного узла, огибая глотку, вливаются в надглоточный нервный узел, который является самой ростральной (передней) частью нервной системы. В состав надглоточного нервного ганглия входят только чувствительные и ассоциативные нейроны. Двигательных элементов там не обнаружено. Таким образом, надглоточный ганглий кольчатых червей является высшим ассоциативным центром, он осуществляет контроль над подглоточным ганглием. Подглоточный ганглий контролирует нижележащие узлы, он имеет связи с двумя-тремя последующими ганглиями, тогда как остальные ганглии брюшной нервной цепочки не образуют связей длинней, чем до соседнего ганглия.
В филогенетическом ряду кольчатых червей есть группы с хорошо развитыми органами чувств (многощетинковые черви). У этих животных в надглоточном ганглии обособляются три отдела. Передний отдел иннервирует щупальца, средняя часть иннервирует глаза и антенны. И наконец, задняя часть развивается в связи с совершенствованием химических органов чувств.
Сходную структуру имеет нервная система членистоногих, т.е. построена по типу брюшной нервной цепочки, однако может достигать высокого уровня развития (рис. 11). Она включает в себя значительно развитый надглоточный ганглий, выполняющий функ-
1 — грибовидное тело; 2 — протоцеребрум; 3 — зрительная лопасть; 4 — дейтоцеребрум; 5 — тритоцеребрум
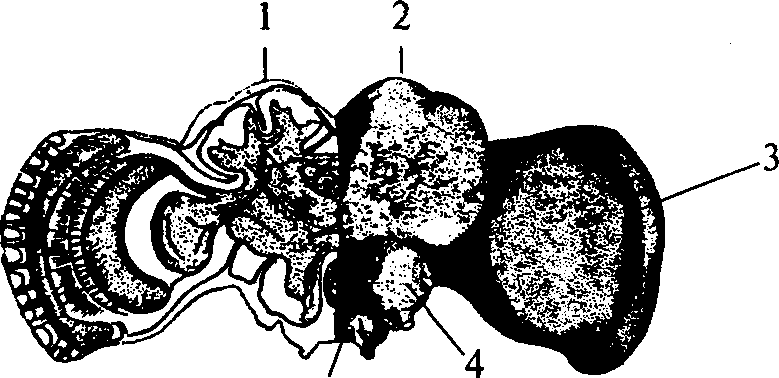
цию мозга, подглоточный ганглий, управляющий органами ротового аппарата, и сегментарные ганглии брюшной нервной цепочки. Ганглии брюшной нервной цепочки могут сливаться между собой, образуя сложные ганглиозные массы.
Головной мозг членистоногих состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Сложным строением отличается мозг насекомых. Особенно важными ассоциативными центрами насекомых являются грибовидные тела, располагающиеся на поверхности протоцеребрума, причем чем более сложным поведением характеризуется вид, тем более развиты у него грибовидные тела. Поэтому наибольшего развития грибовидные тела достигают у общественных насекомых (рис. 12).
Практически во всех отделах нервной системы членистоногих существуют нейросекреторные клетки. Нейросекреты играют важную регулирующую роль в гормональных процессах членистоногих.
В процессе эволюции первоначально диффузно расположенные биполярные нейросекреторные клетки воспринимали сигналы либо отростками, либо всей поверхностью клетки, затем сформировались нейросекреторные центры, нейросекреторные тракты и нейросекреторные контактные области. В последующем произошла специализация нервных центров, увеличилась степень надежности во взаимоотношениях двух основных регуляторных систем (нервной и гуморальной) и сформировался принципиально новый этап регуляции — подчинение нейросекреторным центрам периферических эндокринных желез.
1 — церебральная комиссура; 2 — церебральные ганглии; 3 — педальные ганглии; 4 — коннектива; 5 — висцеральные ганглии
Нервная система моллюсков также имеет ганглионарное строение (рис. 13). У простейших представителей типа она состоит из нескольких пар ганглиев. Каждая пара ганглиев управляет определенной группой органов: ногой, висцеральными органами, легкими и т.д. — и расположена рядом с иннервируемыми органами или внутри их. Одноименные ганглии попарно соединены между собой комиссурами. Кроме того, каждый ганглий связан длинными коннективами с церебральным комплексом ганглиев.
У более высокоорганизованных моллюсков (головоногие) нервная система преобразуется (рис. 14). Ганглии ее сливаются и образуют общую окологлоточную массу — головной мозг. От заднего отдела головного мозга отходят два крупных мантийных нерва и образуют два больших звездчатых ганглия. Таким образом, у головоногих наблюдается высокая степень цефализации.
Среди беспозвоночных наиболее примитивный тип нервной системы в виде диффузной нервной сети встречается у типа кишечнополостных. Их нервная сеть представляет собой скопление нейронов, отростки которых могут перекрещиваться, прилегать друг к другу и лишены функциональной дифференциации на аксоны и дендриты. Диффузная нервная сеть не разделена на центральный и периферический отделы и может быть локализована в эктодерме и энтодерме.
Эпидермальные нервные сплетения, напоминающие нервные сети кишечнополостных, могут быть обнаружены и у более высоко оревизованных беспозвоночных (плоские и кольчатые черви), однако здесь они занимают соподчиненное положение по отношению к Центральной нервной системе (ЦНС), которая выделяется как самостоятельный отдел.
В качестве примера такой централизации и концентрации нервных элементов можно привести ортогональную нервную систему плоских червей. Ортогон высших турбеллярий представляет собой упорядоченную структуру, которая состоит из ассоциативных и двигательных клеток, формирующих вместе несколько пар продольных тяжей, или стволов, соединенных большим числом поперечных и кольцевых комиссуральных стволов. Концентрация нервных элементов сопровождается их погружением в глубь тела.
Плоские черви являются билатерально симметричными животными с четко выраженной продольной осью тела. Движение у свободноживущих форм осуществляется преимущественно в сторону головного конца, где концентрируются рецепторы, сигнализирующие о приближении источника раздражения. К числу таких рецепторов турбеллярий относятся пигментные глазки, обонятельные ямки, ста-тоцист, чувствительные клетки покровов, наличие которых способствует концентрации нервной ткани на переднем конце тела. Этот процесс приводит к формированию головного ганглия, который, по меткому выражению Ч. Шеррингтона, можно рассматривать как ганглиозную надстройку над системами рецепции на расстоянии.
Ганглионизсщия нервных элементов получает дальнейшее развитие у высших беспозвоночных, кольчатых червей, моллюсков и членистоногих. У большинства кольчатых червей брюшные стволы ганглионизированы таким образом, что в каждом сегменте тела формируется по одной паре ганглиев, соединенных коннективами с другой парой, расположенной в соседнем сегменте.
Ганглии одного сегмента у примитивных аннелид соединены между собой поперечными комиссурами, и это приводит к образованию лестничной нервной системы. В более продвинутых отрядах кольчатых червей наблюдается тенденция к сближению брюшных стволов вплоть до полного слияния ганглиев правой и левой сторон и перехода от лестничной к цепочечной нервной системе. Идентичный, цепочечный тип строения нервной системы существует и у членистоногих с различной выраженностью концентрации нервных элементов, которая может осуществляться не только за счет слияния соседних ганглиев одного сегмента, но и при слиянии последовательных ганглиев различных сегментов.
Процесс ганглионизации у беспозвоночных может привести к формированию нервной системы разбросанно-узлового типа, которая встречается у моллюсков. В пределах этого многочисленного типа имеются филогенетически примитивные формы с нервной системой, сопоставимой с ортогоном плоских червей (боконервные-моллюски), и продвинутые классы (головоногие моллюски), у которых слившиеся ганглии формируют дифференцированный на отделы мозг.
Прогрессивное развитие мозга у головоногих моллюсков и насекомых создает предпосылку для возникновения своеобразной иерархии командных систем управления поведением. Низший уровень интеграции в сегментарных ганглиях насекомых и в подглоточной массе мозга моллюсков служит основой для автономной деятельности и координации элементарных двигательных актов. В то же время мозг представляет собой следующий, более высокий уровень интеграции, где могут осуществляться межанализаторный синтез и оценка биологической значимости информации.
В целом, говоря об эволюции нервной системы беспозвоночных, было бы упрощением представлять ее как линейный процесс. Факты, полученные в нейроонтогенетических исследованиях беспозвоночных, позволяют допустить множественное (полигенетическое) происхождение нервной ткани беспозвоночных. Следовательно, эволюция нервной системы беспозвоночных могла идти широким фронтом от нескольких источников с изначальным многообразием.
Впервые в эволюции НС появляется у типа кишечнополостные. Это диффузная НС. Кишечнополостные обладают радиальной (лучевой) симметрией и имеют две жизненные формы — прикрепленные полипы и свободноплавающие медузы. Тело у них состоит из двух слоев клеток — эктодермы и энтодермы. Основу каждого слоя составляют эпителиальномускульные клетки. Таким образом, моторика кишечнополостных бесске- летная, носящая перистальтический характер. Нейроны расположены как в экто-, так и в эктодермальном слоях. Сенсорные функции выполняют специализированные эктодермальные нервные клетки, чувствительные волоски которых выходят на поверхность тела и воспринимают раздражения (см. рис. 16.2, б). Отдельные части тела животных с диффузной НС способны к автономным движениям за счет того, что в каждом участке есть чувствительная клетка, двигательная клетка и нервное волокно. Но даже у самых примитивных кишечнополостных можно найти элементы концентрации — нейроны около рта и на подошве располагаются гуще, чем на остальном теле.
Нервные сети кишечнополостных могут иметь синцитиальное строение, т.е. отростки нервных клеток могут сливаться между собой, проводя возбуждение в разные стороны. Но в большинстве случаев возбуждение передается с помощью типичных синапсов, большинство которых поляризовано, т.е. проводят возбуждение только в одну сторону.
У медуз, которые способны к более активному передвижению, НС несколько усложняется: но краю зонтика у них наблюдаются скопления нервных клеток, которые вместе со своими отростками образуют сплошное нервное кольцо. Таким образом, концентрация нервных элементов у медуз больше, чем у полипов. Более того, у медуз появляются примитивные органы чувств двух типов — статоцисты и глазки — поверхностные скопления светочувствительных клеток. Статоцисты — это органы равновесия, которые можно встретить почти у всех беспозвоночных животных. В большинстве случаев они представляют собой замкнутые пузырьки, на внутренних стенках которых лежат чувствительные клетки. Полость пузырька заполнена жидкостью, в которой находится сгатолит — кристаллик углекислой извести. При изменении положения тела в пространстве чувствительные клетки испытывают давление и возбуждаются.
Животные, следующие в эволюционном развитии за кишечнополостными, — это разные типы червей. Все животные, расположенные на эволюционном древе выше кишечнополостных, — животные трехслойные, проходящие в процессе онтогенеза стадию трех зародышевых листков, которые в своем дальнейшем развитии дают ткани. Таким образом, даже у самых примитивных червей — плоских — уже есть гладкая мышечная ткань, но движения сохраняют перистальтический характер. Концентрация нейронов, только намечавшаяся у медуз, приобретает у червей вполне выраженный характер — появляются нервные ганглии. Но в отличие от кишечнополостных, у которых сокращение мускульных волоконец охватывает все тело, черви способны последовательно сокращать отдельные участки тела, что возможно благодаря иннервации разных участков нейронами, расположенными в разных ганглиях.
Принципиальное отличие червей, как и всех остальных беспозвоночных (за исключением гребневиков и иглокожих) от кишечнополостных, — это наличие билатеральной (двусторонней) симметрии. В связи с этим у них появляется передний конец тела, в сторону которого обычно осуществляется движение. Вследствие этого на нем концентрируются важнейшие органы чувств и, соответственно, передние ганглии укрупняются.
В типе плоские черви, самом примитивном типе червей (например, пла- нария, бычий цепень), централизация нервных клеток еще очень невелика. Их НС состоит из парного головного ганглия и нескольких продольных стволов, соединенных кольцевыми перемычками. Начиная с круглых червей (например, аскарида) у беспозвоночных появляется окологлоточное нервное кольцо, окружающее переднюю часть пищеварительной трубки. Оно обычно состоит из парных надглоточных ганглиев, которые часто сливаются, образуя так называемый головной мозг, и непарного подглоточного ганглия, соединенных нервными волокнами.
В наиболее выраженном виде узловая НС наблюдается в типах кольчатые черви (самые высокоорганизованные черви, например, дождевой червь или нереида), членистоногие и моллюски.
Тело кольчатых червей и членистоногих построено но метамерному принципу и состоит из хорошо выраженных сегментов. От подглоточного узла начинается брюшная нервная цепочка. Исходно (у кольчатых червей) она состоит из парных ганглиев, соединенных комиссурами. Части нервных стволов, соединяющие ганглии каждой стороны соседних сегментов, называются коннективами (см. рис. 16.3, а). Вся цепь имеет характерный вид лестницы (лестничная НС). У разных видов кольчецов наблюдается постепенное сближение и слияние парных ганглиев, при этом коннективы остаются парными. В дальнейшем объединяются и коннективы, и в своем классическом виде брюшная нервная цепочка выглядит как на рис. 16.3, б.
Расположение ганглиев на брюшной стороне связано, по-видимому, с расположением основных рецептивных областей. Черви передвигаются путем ползания и получают многие сенсорные импульсы от вентральной поверхности. В передней же части тела наибольшее значение имеет рецепция, получаемая от рострального конца, в связи с чем головной ганглий поднимается вверх.
НС членистоногих (ракообразные, паукообразные, насекомые) устроена значительно совершеннее, чем у кольчатых червей. Это связано как с более сложным поведением, так и с особенностями организации. Для этих животных характерна гетерономная сегментация, т.е. сегменты тела не одинаковы, как у кольчатых червей, а отличаются друг от друга как по строению, так и по функциям. Конечности состоят из нескольких члеников, подвижно соединенных друг с другом и с телом с помощью суставов. Следовательно, принципиально меняется характер движения — он становится рычаговым. Помимо этого, большая часть гладкой мускулатуры заменяется поперечнополосатой (впервые в эволюции), что способствует гораздо более быстрым движениям. Что касается НС, то концентрация и централизация нейронов (если говорить о беспозвоночных) достигает своего максимума. Нередко наблюдается слияние нервных узлов брюшной нервной цепочки с образованием более крупных ганглиев, т.е. наблюдается и продольная концентрация элементов НС.
Но особенно сильно НС членистоногих отличается от НС червей в строении ее головного конца. У большинства этих животных ГМ (надглоточный ганглий) состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Каждый отдел головного мозга иннервирует разные органы чувств и состоит из нескольких ганглиев, отделенных друг от друга белым веществом. В протоцере- бруме расположены особенно важные ассоциативные центры — грибовидные тела (рис. 16.5).
Наибольшее развитие головного мозга наблюдается у общественных насекомых (муравьев, пчел, термитов), т.е. у животных с особенно сложным поведением. Причем даже в пределах одного вида величина мозга может значительно варьироваться в зависимости от сложности поведения. Так, например, у рабочих муравьев грибовидные тела развиты значительно сильнее, чем у цариц и самцов.
Что касается моллюсков, то это резко отграниченный тип беспозвоночных, ведущий начало от древних кольчатых червей, но имеющий несегмен- тированное тело. От окологлоточного кольца у моллюсков отходят четыре продольных нервных ствола. У высших форм моллюсков на стволах формируются парные ганглии, соединенные коннективами, некоторые ганглии соединяются и с помощью комиссур (рис. 16.6). Нервные клетки имеются не только в ганглиях, но и рассеяны в периферических нервах. Такой тип нервной системы называют разбросанно-узловым.

Рис. 16.5. Нервная система насекомого

Рис. 16.6. Нервная система брюхоногого моллюска
У высших моллюсков крупные ганглии расположены, как правило, рядом с иннервируемым органом или непосредственно в его стенках. Надглоточные ганглии иннервируют глаза и статоцисты, глотку и щупальца; ганглии, иннервирующие мускулатуру ноги, расположены в самой ноге; рядом с ними лежат ганглии, иннервирующие мантию; следующая пара ганглиев связана с жабрами и органами химического чувства; еще одна пара управляет внутренними органами.
Особенно высокой сложности строения НС достигает у головоногих моллюсков (рис. 16.7).

Рис. 16.7. Головной мозг головоногого моллюска
Большинство этих животных — свободноплавающие и очень подвижные хищники. Для них характерно настолько сложное поведение, что их иногда называют приматами моря. Ганглии у них очень велики и образуют сплошную окологлоточную нервную массу (ГМ). В этой общей массе выделяют отдельные ганглии, выполняющие различные функции. В мозге головоногих даже найдены участки (в зрительной доле), имеющие слоистое строение. Для защиты такой НС головоногие имеют подкожную хрящевую капсулу, окружающую мозг аналогично черепу позвоночных.
Нервная система у всех животных имеет эктодермальное происхождение. Она выполняет следующие функции:
- связь организма с окружающей средой (восприятие, передача раздражения и ответная реакция на раздражение);
- связь всех органов и систем органов в единое целое;
- нервная система лежит в основе формирования высшей нервной деятельности.
Впервые нервная система появилась у кишечнополостных и имела диффузный или сетчатый тип нервной системы, т.е. нервная система представляет собой сеть нервных клеток, распределенных по всему телу и связанных между собой тонкими отростками. Типичное строение она имеет у гидры, но уже у медуз и полипов появляются скопления нервных клеток в определенных местах (около рта, по краям зонтика), эти скопления нервных клеток являются предшественниками органов чувств.
Дальше эволюция нервной системы идет по пути концентрации нервных клеток в определенных местах тела, т.е. по пути образования нервных узлов (ганглиев). Эти узлы в первую очередь возникают там, где находятся клетки воспринимающие раздражение из окружающей среды. Так при радиальной симметрии возникает радиальный тип нервной системы, а при билатеральной симметрии концентрация нервных узлов происходит на переднем конце тела. От головных узлов отходят парные нервные стволы, идущие вдоль тела. Такой тип нервной системы называется ганглиозно-стволовым.
Типичное строение этот тип нервной системы имеет у плоских червей, т.е. в переднем конце тела имеются парные ганглии, от которых отходят вперед нервные волокна и органы чувств, и нервные стволы, идущие вдоль тела.
У круглых червей головные ганглии сливаются в окологлоточное нервное кольцо, от которого также идут нервные стволы вдоль тела.
У кольчатых червей образуется нервная цепочка, т.е. в каждом членике формируются самостоятельные парные нервные узлы. Все они соединяются как продольными, так и поперечными тяжами. В результате нервная система приобретает строение, напоминающие лестницу. Часто обе цепочки сближаются, соединяясь по средней части тела в непарную брюшную нервную цепочку.
У членистоногих такой же тип нервной систем, но количество нервных узлов уменьшается, а размер их увеличивается, особенно в головном или в головогрудном отделе, т.е. идет процесс цефализации.
У моллюсков нервная система представлена узлами в разных отделах тела, соединенных между собой тяжами и отходящими от узлов нервами. У брюхоногих моллюсков имеются педальный, церебральный и плеврально-висцеральные узлы; у двустворчатых – педальный и плеврально-висцеральный; у головоногих – плеврально-висцеральный и церебральный нервные узлы. Вокруг глотки у головоногих моллюсков наблюдается скопление нервной ткани.
Нервная система у хордовых представлена нервной трубкой , которая дифференцируется на головной и спинной мозг.
У низших хордовых нервная трубка имеет вид полой трубки (невроцель) с отходящими от трубки нервами. У ланцетника в головном отделе образуется небольшое расширение – зачаток головного мозга. Это расширение получило название желудочка.
У высших хордовых на переднем конце нервной трубки образуется три вздутия: передний, средний и задний пузыри. Из первого мозгового пузыря образуется в дальнейшем передний и промежуточный мозг, из среднего – средний, из заднего – мозжечок и продолговатый мозг, переходящий в спинной.
У всех классов позвоночных животных мозг состоит из 5 отделов (передний, промежуточный, средний, задний и продолговатый), но степень их развития неодинакова у животных разных классов.
Так у круглоротых все отделы головного мозга расположены друг за другом в горизонтальной плоскости. Продолговатый мозг непосредственно переходит в спинной с центральным каналом в нутрии.
У рыб головной мозг более дифференцирован по сравнению с круглоротыми. Объем переднего мозга увеличен, особенно у двоякодышащих рыб, но передний мозг еще не разделен на полушария и функционально служит высшим обонятельным центром. Крыша переднего мозга тонкая, она состоит только из эпителиальных клеток и не содержит нервной ткани. В промежуточном мозге, с которым связан эпифиз и гипофиз, расположен гипоталамус, являющийся центром эндокринной системы. Наиболее развитым у рыб является средний мозг. Хорошо выражены в нем зрительные доли. В области среднего мозга имеется изгиб, характерный для всех вышестоящих позвоночных. Кроме того средний мозг является анализирующим центром. Мозжечок, входящий в состав заднего мозга, развит хорошо в связи со сложностью движения у рыб. Он представляет собой центр координации движения, его размер варьирует в зависимости от активности движения разных видов рыб. Продолговатый мозг обеспечивает связь высших отделов головной мозга со спинным и содержит центры дыхания и кровообращения.
Из головного мозга рыб выходит 10 пар черепно-мозговых нервов.
Такой тип мозга, в котором высшим центром интеграции является средний мозг, называется ихтиопсидным.
У амфибий нервная система по своему строению близка к нервной системе двоякодышащих рыб, но отличается значительным развитием и полным разделением парных вытянутых полушарий, а также слабым развитием мозжечка, что обусловлено малой подвижностью амфибий и однообразием их движений. Но у амфибий появилась крыша переднего мозга, называемая первичным мозговым сводом – архипаллиумом. Число черепно-мозговых нервов, как и у рыб, десять. И тип мозга тот же, т.е. ихтиопсидный.
Таким образом у всех анамний (круглоротых, рыб и амфибий) ихтиопсидный тип головного мозга.
В строении головного мозга рептилий, относящихся к высшим позвоночным, т.е. к амниотам, отчетливо выражены черты прогрессивной организации. Значительное преобладание над другими отделами мозга получают полушария переднего мозга. У их основания расположены крупные скопления нервных клеток – полосатые тела. На латеральной и медиальной сторонах каждого полушария появляются островки старой коры – архикортекс. Размеры среднего мозга сокращаются, и он теряет значение ведущего центра. Анализирующим центром становится дно переднего мозга, т.е. полосатые тела. Такой тип мозга называется зауропсидный или стриарным . Мозжечок увеличен в размерах в связи с многообразием движений пресмыкающихся. Продолговатый мозг образует резкий изгиб, характерный для всех амниот. Из головного мозга выходит 12 пар черепно-мозговых нервов.
Такой же тип мозга характерен и для птиц, но с некоторыми особенностями. Полушария переднего мозга относительно большие. обонятельные доли у птиц развиты слабо, что указывает на роль обоняния в жизни птиц. В противоположность этому средний мозг представлен крупными зрительными долями. Хорошо развит мозжечок, из головного мозга выходит 12 пар нервов.
Головной мозг у млекопитающих достигает максимального развития. Полушария настолько велики, что покрывают средний мозг и мозжечок. Особо развита кора больших полушарий, площадь ее увеличена за счет извилин и борозд. Кора имеет очень сложное строение и называется новой корой – неокортекс. Появляется вторичный мозговой свод – неопаллиум. Спереди от полушарий расположены крупные обонятельные доли. Промежуточный мозг, как и у других классов, включает эпифиз, гипофиз и гипоталамус. Средний мозг относительно мал, он состоит из четырех бугров – четыреххолмия. Передняя кора связана со зрительным анализатором, задняя – со слуховым. Наряду с передним мозгом сильно прогрессирует мозжечок. Из мозга выходит 12 пар черепно-мозговых нервов. Анализирующим центром является кора больших полушарий. Такой тип мозга называется маммальным .
1. Ацефалия - отсутствие головного мозга, свода, черепа и лицевого скелета; это нарушение связано с недоразвитием переднего отдела нервной трубки и сочетается с дефектами спинного мозга, костей и внутренних органов.
2. Анэнцефалия - отсутствие больших полушарий и крыши черепа при недоразвитии ствола мозга и сочетается с другими пороками развития. Эта патология обусловлена незакрытием (дизрафия) головной части нервной трубки. При этом не развиваются кости крыши черепа, а кости основания черепа обнаруживают различные аномалии. Анэнцефалия не совместима с жизнью, средняя частота 1/1500, при чем чаще у женских плодов.
3. Ателэнцефалия – остановка развития (гетерохрония) передней части нервной трубки на стадии трех пузырей. В результате большие полушария и подкорковые ядра не формируются.
4. Прозэнцефалия – конечный мозг делится продольной бороздой, но в глубине оба полушария остаются связанными друг с другом.
5. Голопрозэнцефалия – конечный мозг не делится на полушария и имеет вид полусферы с единой полостью (желудочком).
6. Алобарная прозэнцефалия – разделение конечного мозга только в задней части, а лобные доли остаются неразделенными.
7. Аплазия или гипоплазия мозолистого тела – полное или частичное отсутствие сложной комиссуры мозга, т.е. мозолистого тела.
8. Гидроэнцефалия - атрофия больших полушарий в сочетании с гидроцефалией.
9. Агирия - полное отсутствие борозд и извилин (гладкий мозг) больших полушарий.
10. Микрогирия - уменьшение числа и объема борозд.
11. Врожденная гидроцефалия - преграждение части желудочковой системы мозга и ее выходов, она вызвана первичным нарушением развития нервной системы.
12. Spina bifida - дефект замыкания и обособления от кожной эктодермы спинального отдела нервной трубки. Иногда эта аномалия сопровождается дипломиелией, при которой спинной мозг расщеплен на известном протяжении на две части, каждая со своим центральным карманом.
13. Иниэнцефалия - редкая аномалия, несовместимая с жизнью, встречается чаще у плодов женского пола. Это грубая аномалия затылка и головного мозга. Головы повернута так, что лицо обращено кверху. Дорсально скальп продолжается в кожу люмбодорсальной или сакральной области.
Читайте также:
