
Нервно мышечная физиология животных
ФИЗИОЛОГИЯ МЫШЦ
Мышцы выполняют в организме животных двигательные функции. Поперечнополосатые скелетные мышцы осуществляют перемещение тела или отдельных его частей в пространстве, с их помощью происходит акт вдоха и выдоха. Поперечнополосатая мускулатура сердца обеспечивает ритмическое перекачивание в артерии крови, притекающей к нему из вен. Гладкая мускулатура внутренних органов, кровеносных сосудов поддерживает длительные тонические сокращения сфинктеров внутренних органов, а также тонус стенок кровеносных сосудов. Ритмические сокращения гладких мышц стенок полых органов (желудка, кишок, протоков пищеварительных желез и др.) обеспечивают передвижение и выделение содержимого этих полых органов.
Физиологические свойства мышц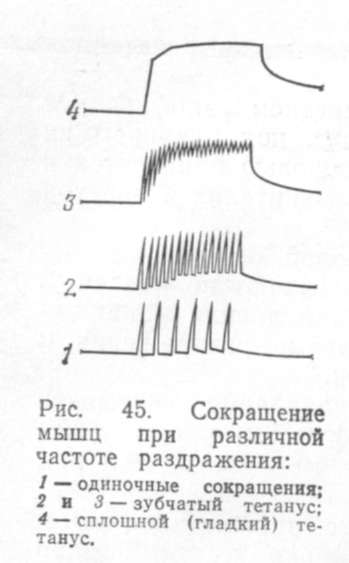
. Основными свойствами мышц являются возбудимость, проводимость и сократимость.
Возникшее в скелетной мышце возбуждение проводится изолированно, т. е. не переходит с одного мышечного волокна на другое. Скорость распространения возбуждения зависит от строения мышечных волокон. В белых волокнах с большим количеством миофибрилл она составляет 12-15, а в красных - 3-4 м/с.
Специфической деятельностью мышечной ткани является ее сокращение при возбуждении. Различают изотонические и изометрические сокращения мышц. При изотоническом сокращении, наблюдаемом, например, при поднятии мышцей груза, волокна ее укорачиваются, но напряжение остается постоянным. Изометрическим называется такое сокращение, при котором мышца не укорачивается, например, если оба конца ее неподвижно закреплены, но зато напряжение мышечных волокон возрастает.
В зависимости от частоты поступающие к мышце раздражений может наступит одиночное или тетаническое ее сокращением В условиях опыта на однократное кратковременное раздражение мышца отвечает одиночным сокращением. На кривой сокращения мышцы (рис. 45) отмечают три пе-Я риода: скрытый, или латентный, - от момента раздражения до начала сокращения, период укорочения и период расслабления. Общая продолжительность одиночного сокращения скелетных мышц у млекопитающих колеблется от 0,04 до 0,1 с. В естественных условиях одиночные сокращения не наблюдаются. Центральная нервная система посылает к мышце не одиночные импульсы, а целый ряд возбуждающих импульсов, в результате этого наступает длительное и сильное ее сокращение.
Длительное сокращение мышцы получило название тетанического, или тетануса. При редких раздражениях, не более 10 в 1 с, возникают одиночные сокращения, при частоте раздражения от 10 до 25 в 1 с - зубчатый тетанус. Если частота раздражения превышает 25 в 1 с, то наступает полный, или гладкий, тетанус (см. рис. 45).
Источником энергии при работе мышц являются химические процессы, которые совершаются в две фазы: анаэробную и аэробную. В анаэробную фазу выделение энергии происходит при распаде аденозинтрифосфорной кислоты (АТФ). Во время аэробной фазы происходит окисление молочной кислоты до углекислого газа и воды также с выделением энергии. При сокращении мышц большая часть энергии превращается в тепловую и только 25- 30% ее преобразуется в механическую.
В настоящее время механизм мышечного сокращения объясняют скольжением протофибрилл. В состоянии покоя мышцы прото-фибриллы расположены в миофибрилле таким образом, что концы тонких нитей актина лежат частично в промежутке между миозиновыми нитями и соединены друг с другом поперечными мостиками 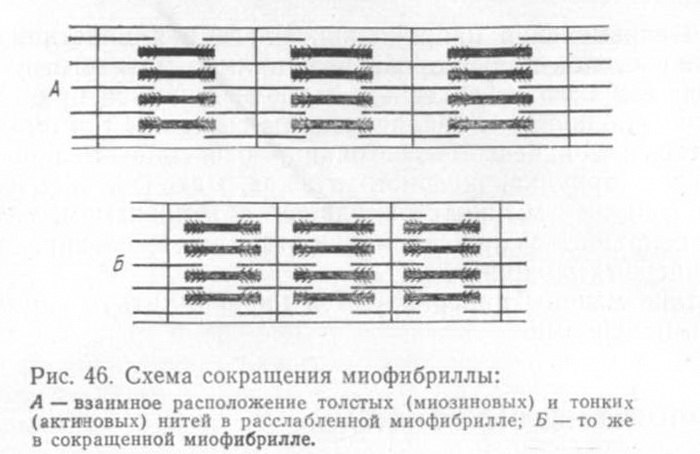
(рис. 46). При сокращении мышцы тонкие нити актина сдвигаются в промежутки между толстыми нитями миозина, что сопровождается укорочением миофибрилл и мышцы (рис. 46,Б).
Сила и работа мышц. Силу мышц определяют по максимальному напряжению, которое она может развить в условиях изометрического сокращения или при поднятии максимального груза. Для измерения силы мышцы определяют тот максимальный груз, который она в состоянии поднять.
Сила мышц при прочих равных условиях зависит не от длины, а от ее поперечного сечения. Чтобы иметь возможность сравнивать силу разных мышц, максимальный груз, который мышца в состоянии поднять, делят на число квадратных сантиметров ее поперечного сечения. Абсолютная сила мышц выражается в кг на 1 см 2 .
Поднимая груз, мышца выполняет механическую работу, которая измеряется произведением массы груза на высоту его подъема и выражается в килограммометрах. Мышца выполняет наибольшую работу при средних нагрузках.
Временное понижение работоспособности мышцы, наступающее в результате работы и исчезающее после отдыха, называется утомлением. Последнее представляет собой сложный физиологический процесс, связанный, прежде всего, с утомлением нервных центров. Определенную роль в развитии утомления играет накопление в работающей мышце продуктов обмена (молочная кислота и др.) и постепенное истощение энергетических запасов.
В покое, вне работы, мышцы полностью не расслаблены, а сохраняют некоторое напряжение, называемое тонусом. Внешним выражением тонуса является определенная степень упругости мышц. Тонус мышц обусловлен непрерывно поступающими нервными импульсами из мотонейронов спинного мозга. Тонус скелетных мышц играет важную роль для поддержания определенного положения тела в пространстве, сохранения равновесия и упругости мышц.
Особенности гладкой мускулатуры. Гладкая мускулатура находится во внутренних органах, в сосудах и коже. В отличие от поперечнополосатых они сокращаются медленно. Скрытый период их сокращения в 300 раз превышает таковой скелетной мускулатуры. Возбуждение в гладкой мышце проводится очень медленно (от 1 см/с в кишечнике до 18 см/с в мочеточнике) и передается от одного гладкого волокна к другому.
Важным свойством гладкой мышцы является ее большая пластичность, т. е. способность сохранять приданную растяжением длину без изменения напряжения. Благодаря пластичности гладких мышц стенок полых органов, например мочевого пузыря, давление внутри него относительно мало изменяется при разной сты пени его наполнения. Гладкие мышцы способны длительное время находиться в тоническом состоянии, особенно это проявляется в сфинктерах желудка, желчного пузыря, матки и других органов. Многие гладкие мышцы обладают автоматизмом, т. е. способностью сокращаться под влиянием импульсов, возникающих в самих мышечных волокнах.
Гладкие мышцы иннервируются парасимпатическими и симпатическими нервами.
Физиологические особенности мышечной ткани.
Свойства нервного волокна.
Механизм передачи возбуждения в нервно-мышечном синапсе.
1. Физиологические особенности мышечной ткани
У позвоночных животных есть 2 вида мышечной ткани: гладкая и поперечно-полосатая. Главная роль в осуществлении движений принадлежит скелетной мускулатуре. Кроме поперечно-полосатой скелетной мускулатуры существует поперечно-полосатая мускулатура сердца, которая отличается от скелетной наличием анастомозов между мышечными волокнами. Поперечно-полосатые мышцы формируют двигательные аппараты скелета, глазодвигательные, жевательные и другие двигательные системы у животных. Поперечно-полосатые мышцы, за исключением сердечной мышцы, полностью контролируются центральной нервной системы. Они лишены автоматизма, то есть не способны работать без сигналов, поступающих из центральной нервной системы. Гладкая мышечная ткань входит в состав мышечных оболочек внутренних органов и кровеносных сосудов. Они в значительно меньшей мере контролируются центральной нервной системой, обладают автоматизмом и собственной нервной сетью, расположенной в мышечной стенке (интрамуральной сетью) и в значительной степени обеспечивающей их самоуправление. Структурно-функциональной единицей мышц является многоядерное мышечное волокно. Объединённые в пучки эти волокна образуют мышцу. Каждое волокно покрыто оболочкой – сарколеммой, и содержит тонкие нити – миофибриллы, а также митохондрии и саркоплазматическую сеть (система поперечных трубочек – Т-система). Каждая миофибрилла сократительное вещество состоит из множества параллельно лежащих толстых и тонких нитей – миофиламентов. Толстые нити или филаменты состоят из белка – миозина, тонкие из актина. Тонкие нити представлены двойной нитью закрученной в двойную спираль. В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина. К молекуле тропомиозина прикреплена молекула тропонина. Расположение толстых миозиновых нитей и тонких актиновых нитей строго упорядочено. Актиновые нити частично входят своими концами в промежутки между миозиновыми нитями. Толстые нити в световом микроскопе, - тёмная полоса, а актиновые - светлая полоса. Благодаря периодическому чередованию светлых и тёмных полос миофибриллы скелетных мышц выглядят поперечнополосатыми.
Механизм сокращения мышечного волокна
Свойства мышечной ткани
Возбудимость, проводимость, сократимость, растяжимость, эластичность, пластичность, автоматизм.
Возбудимость – способность возбудимой ткани отвечать на раздражения. Способность мышцы реагировать на раздражение его двигательного нерва обозначается как непрямая возбудимость мышцы. Способность мышечных волокон реагировать на раздражение, действующее непосредственно на них – прямой.
Проводимость – проявление возбуждения в мышцах происходит изолированно, то есть не переходит с одного мышечного волокна на другое.
Сократимость – способность в ответ на раздражение характерным образом изменять свою форму и развивать механическое напряжение на своих концах.
Скелетная мышца является упругим телом. Если к ней подвесить груз, то она растягивается. Свойство мышцы удлиняться под влиянием нагрузки называется растяжимостью.
Эластичностью называется свойство деформированного тела возвращаться к первоначальному своему состоянию после удаления нагрузки (груза), вызвавшей деформацию.
Пластичностью называется свойство тела сохранять приданную ему длину или форму после прекращения действия внешней деформирующей силы (присуще мёртвой ткани).
Сокращение мышц
При раздражении мышцы одиночным импульсом тока пороговой или надпороговой силы возникает одиночное мышечное сокращение. В нём различают латентный (скрытый) период, период сокращения и период расслабления. Многократная стимуляция с интервалами достаточной длительности будет вызывать ряд отдельных сокращений. Но если интервал укорачивать – сокращения будут сливаться и в конце концов возникает одно длительное сокращение – тетанус. К тетаническим сокращениям способны только скелетные мышцы. Величина тетануса зависит от того, при какой возбудимости застаёт мышцу каждый очередной импульс. Если каждый очередной импульс, действующий на мышцу застаёт её в фазу укорочения от предыдущего импульса, то сокращение мышцы записывается в виде сплошной линии – гладкий тетанус. Если каждый очередной импульс застаёт мышцу в фазу расслабления, то сокращение мышцы записывается в виде зубчатой линии – зубчатый тетанус.
Деятельность мышцы связана с осуществлением работы. Работа мышцы, при которой происходит перемещение груза, называется динамической – изотоническая. Если мышца производит работу, развивая напряжение без укорочения мышечных волокон – статистическая или изометрическая работа. Механическая работа может быть измерена произведением массы груза на высоту подъёма. Внешняя механическая работа мышцы по мере возрастания груза вначале увеличивается, а затем уменьшается. Наибольший груз, который мышца ещё способна поднять показывает абсолютную силу мышц.
Зависимость работы от величины груза выражается законом средних нагрузок: работа мышцы будет наибольшей при средних нагрузках.
Утомление мышц – при длительном сокращении мышца утомляется. Утомлением называется временное понижение работоспособности мышцы наступающей при длительном сокращении и исчезающей после отдыха. Утомление характеризуется: 1) уменьшением силы сокращения; 2) уменьшением степени расслабления.
Причины: 1) накопление продуктов обмена веществ; 2) уменьшение запаса кислорода; 3) истощение энергетических запасов; 4) израсходование запасов медиатора.
Тонус мышц – скелетные мышцы в покое расслабляются не полностью, а находятся в некотором напряжении, в тонусе. Тонус – это способность скелетных мышц длительно находиться на том или ином уровне напряжения под влиянием редких раздражений. Он играет важную роль в поддержании определённого положения тела в пространстве и в деятельности двигательного аппарата.
Физиологические особенности гладких мышц
Гладкие мышцы не имеют выраженной поперечной исчерченности. Они состоят из удлиненных клеток веретеновидной формы.
Характеризуются медленными движениями и более длительными тоническими сокращениями (в гладких мышцах очень велик скрытый период возбуждения – 0,2 – 1 с).
Обладают высокой пластичностью и упругостью.
Они менее возбудимы.
У них более длительный рефрактерный период.
Малая скорость проведения возбуждения. Скорость проведения возбуждения в кишечнике около 1см/с, в матке и мочеточнике – около 18 см/с, в гладких мышцах мигательной перепонки кошки – 80 см/с.
Им характерно свойство автоматизма, то есть способность к спонтанному сокращению независимо от внешних условий.
Высокая чувствительность к некоторым химическим веществам – ацетилхолин, адреналин, норадреналин.
При редких импульсах раздражений впадают в тетаническое сокращение.
Адекватным раздражителем является их растяжение.
Согласно современным представлениям, нервная и мышечная ткани могут находиться в трех основных состояниях — физиологическом покое, возбуждении и торможении.
Физиологический покой — это такое состояние, когда ткань или орган не проявляет признаков присущей им деятельности.
Возбуждение — деятельное состояние живой ткани, в которое она приходит под влиянием раздражения.
Торможение — такое состояние, когда деятельность ткани или органа ослабляется или полностью прекращается.
Раздражители.При раздражении мышца приходит в состояние возбуждения, которое распространяется по мышце и вызывает ее сокращение. По биологическому действию раздражители бывают адекватные и неадекватные. Адекватный — это такой раздражитель, к которому данный орган или ткань приспособились в процессе эволюции. Для мышцы адекватным раздражителем является нервный импульс. Неадекватными — будут такие раздражители, действию которых ткань или орган в естественных условиях обычно не подвергаются. Так, сокращение мышцы можно вызвать механическим раздражением (укол, кислота, ток) и т.д.
Все раздражители по своей силе деляг на пороговые, подпороговые и сверхпороговые. Пороговыми называют минимальные раздражители, которые могут вызвать возбуждение. Подпороговые — это раздражители, сила которых меньше пороговой, сверхпороговые — раздражители более сильные, чем пороговые.
Биоэлектрические явления.Возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки (рис. 4.19).
Потенциал покоя.Мембрана нервной или мышечной клетки в состоянии покоя с наружной стороны заряжена положительно, а с внутренней — отрицательно. Разность зарядов между наружной и внутренней поверхностью клеточной мембраны в состоянии физиологического покоя клетки называется потенциалом покоя, или мембранным потенциалом.
Потенциал действия. Под влиянием раздражения проницаемость клеточной мембраны для ионов натрия повышается в сотни
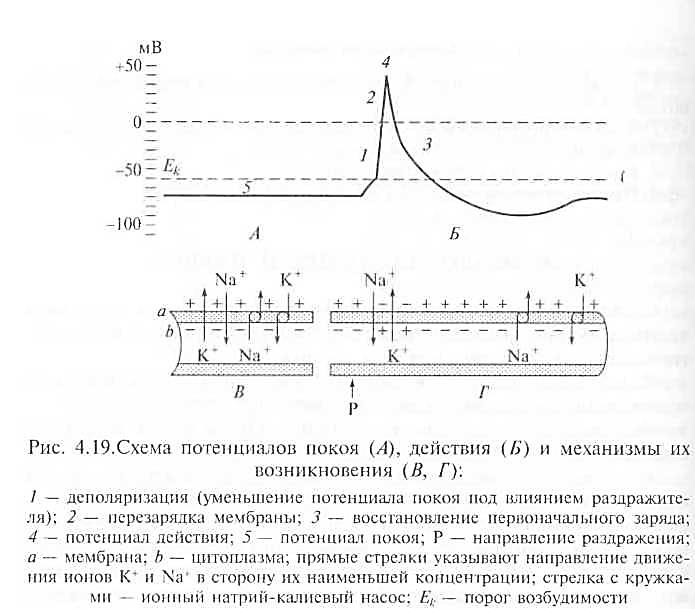 |
Рис. 4.19.Схема потенциалов покоя (А), действия (5) и механизмы их
возникновения (В, Г):
1 — деполяризация (уменьшение потенциала покоя под влиянием раздражите-
ля); 2 — перезарядка мембраны; 3 — восстановление первоначального заряда;
4 — потенциал действия; 5 — потенциал покоя; Р — направление раздражения;
а — мембрана; b — цитоплазма; прямые стрелки указывают направление движе-
ния ионов К + и в сторону их наименьшей концентрации; стрелка с кружка-
ми — ионный натрий-калиевый насос; Ек — порог возбудимости
раз. Происходит перезарядка клеточной мембраны: внутренняя ее сторона заряжается положительно, а наружная — отрицательно. Затем значительно повышается проницаемость мембраны для ионов калия, они начинают быстро перемещаться из клеток во внеклеточную жидкость и вновь восстанавливают первоначальный заряд мембраны. Пикообразное колебание потенциала, возникающее в результате кратковременной перезарядки мембраны и последующего восстановления ее заряда, называется потенциалом действия. Такие колебания потенциала продолжаются 1 — 5 мс.
Проведение возбуждения. Круговой ток, проходя через участок покоя, раздражает его и вызывает появление потенциала действия (рис. 4.20). Когда в одном из участков нервной клетки, нервного или мышечного волокна возникает возбуждение и появляется потенциал действия, между возбужденным и соседним невозбужденным участком возникает разность потенциалов. Эта разность потенциалов является причиной появления круговых (локальных) токов. Внутри клетки ток идет от возбужденного участка к участку покоя, по внешней стороне — от участка покоя к возбужденному.
В участке, который ранее был возбужден, происходит восста
Рис. 4.20. Схема распрост-
ранения возбуждения:
А — волокна мышц и безмякотных нервных волокон; Б —
мякотное нервное волокно; 1 — мембрана; 2 — волокно;
а, Ь, с — перехваты Ранвье; заштрихованы возбужденные
участки; светлые — участки покоя; стрелками показано движение круговых токов

новление потенциала покоя. Данный процесс многократно повторяется и обусловливает распространение импульсов возбуждения. Скорость проведения возбуждения по скелетным мышцам 12 —15 м/с, гладким мышцам — 1 — 18 см/с, в безмякотных нервах — 0,5 — 3 м/с.
Основные свойства живой ткани.Любая живая клетка обладает свойствами раздражимости, возбудимости и лабильности (функциональной подвижности).
Раздражимость. С раздражимостью связаны все процессы роста и размножения клеток и тканей. На разнообразные воздействия внешней или внутренней среды — раздражения протоплазма живой клетки отвечает специфической реакцией, при которой происходят усиление или ослабление обмена веществ, количественное и качественное его изменение. Эта реакция на раздражение получила название раздражимости, или реактивности.
Возбудимость — свойство нервной или мышечной ткани отвечать на действие раздражителя возбуждением. Для измерения величины возбудимости определяют порог возбудимости и хронаксию. Порогом возбудимости называется наименьшая сила раздражителя, способная вызвать возбуждение; его выражают в вольтах постоянного тока.
Лабильность — скорость, с которой в ткани возникает и успевает закончиться полный период отдельного импульса возбуждения. Мера лабильности — это максимальное число импульсов возбуждения за 1 св ответ на такое же максимальное число раздражений. Высокой лабильностью обладают мякотные нерпы, у без- мякотных нервов она низкая. Так, максимальный ритм возбуждений мякотного нервного волокна 500, безммкотного — 200. Максимальный ритм возбуждений скелетной мышцы 200 импульсов в 1 с, а гладкой мышцы — в десятки раз меньше.
При очень частых раздражениях сокращения мышцы уменьшаются или даже совсем прекращаются. Такая частота называется пессимальной, или пессимумом. Пессимум возникает вследствие того, что возбуждение еще не закончилось и ткань находится в состоянии абсолютной или относительной рефрактерное™, а на нее действует новое раздражение. Частые раздражения, превышающие меру лабильности, вызывают не возбуждение, а торможение.
Учение Н. Е. Введенского о парабиозе устанавливает связь между возбуждением и торможением. Возбуждение и торможение — это различные реакции ткани на раздражение, исход которогозависит от лабильности. При высокой лабильности возникает возбуждение, снижение лабильности вызывает торможение.
ФИЗИОЛОГИЯ МЫШЦ
Скелетные мышц, входящие в состав опорно-двигательного аппарата позвоночных животных, состоят из группы мышечных пучков, каждый из них составлен из тысяч мышечных волокон, которые представляют собой клетки цилиндрической формы длиной до 12 см и диаметром 10 — 100 мкм. Каждое волокно окружено оболочкой сарколеммой и содержит тонкие нити — миофибрил-лы (рис. 4.21). Поперечные мембраны делят каждую миофибриллу на отдельные участки — саркомеры. Сократимым веществом мышечного волокна являются миофибриллы, состоящие из множества (около 2500) тонких и толстых белковых нитей — протофибрилл. Толстые протофибриллы образованы из белка миозина, тонкие — из актина. Нити актина прикреплены к мембране саркомера; они образуют светлые участки миофибриллы. В темных участках находятся нити миозина. Акгиновые нити частично входят своими концами в промежутки между миозиновыми нитями. Нити актина и миозина соединяются между собой многочисленными поперечными мостиками, которые образованы скрученными в спираль отростками — мостиками миозиновой нити. Чередование нитей в миофибрилле обусловливает ее поперечную исчерченность.

Рис. 4.21. Схема чередова-
ния нитей в миофибрилле:
А — электронно-микроскопическая фотография мио-
фибриллы; Б — строение миофибриллы; Z — поперечная
мембрана, разделяющая миофибриллу на отдельные уча-
стки — саркомеры; 1 — миозиновая нить, 2 — актиновая нить
Гладкие мышцы, образующие стенки внутренних органов, состоят из клеток различной формы длиной от 30 до 500 мкм и диаметром 2—10 мкм; у клеток имеются боковые отростки, благодаря которым клетки гладких мышц группируются в длинные пучки. Они, в свою очередь, при помощи тяжей соединяются друг с другом, обеспечивая деятельность мышцы как единой системы. Сократительным аппаратом гладких мышц являются миофибриллы, в которых содержатся тонкие актиновьге нити с прикрепленными к ним короткими отростками миозиновых нитей, называемых димерами.
Свойства скелетных мышц.Проведение возбуждения в мышцах происходит изолированно, т. е. потенциалы действия не переходят с одного мышечного волокна на другое. Возбудимость скелетныхмышц меньше возбудимости нервов. Нервное волокно оканчивается в середине мышечного волокна, поэтому возбуждение распространяется в обе стороны со скоростью 4—15 м/с.
Скелетная мышца является упругим телом. Если к мышце подвесить груз, то она растягивается, это свойство называется растяжимостью. Эластичностью мышцы называется возвращение мышцы к первоначальной своей длине после растяжения, когда будет удален груз. Пластичностью мышцы называется свойство сохранять удлиненную форму после удаления груза, вызвавшего ее растяжение. В скелетных мышцах лучше выражено свойство эластичности, чем пластичности.
Виды мышечных сокращений.При нанесении на мышцу одиночного раздражения она отвечает одиночным сокращением. Записывая это сокращение на двигающейся ленте какого-либо самописца (например, в простейшем случае на кимограф), можно отметить три периода: латентный — период от раздражения до начала сокращения, период сокращения и период расслабления. У икроножной мышцы лягушки одиночное сокращение длится 0,1 с, у млекопитающих — 0,04 — 0,1 с. В латентный период в мышце происходят процессы, при которых освобождается энергия для мышечного сокращения.
Если к мышце поступает несколько частых импульсов возбуждения, наступает длительное сокращение мышцы, которое называется тетаническим, или тетанусом. В зависимости от частоты возбуждений тетанус будет зубчатым или гладким (рис. 4.22).
Зубчатый тетанус наблюдается при такой частоте импульсов возбуждений, когда каждый импульс действует на мышцу в тот момент, когда она уже начинает расслабляться. Если же импульсы возбуждения настолько частые, что они воздействуют на мышцу до начала ее расслабления, то получатся длительные непрерывные сокращения мышцы — гладкий тетанус.
 |
Рис. 4.22. Сокращение скелетной мышцы при различной частоте раздра-
жения:
А — одиночное сокращение в ответ на одно раздражение; Б — зубчатый тетанус
(до 15 раздражений в 1 с); В — гладкий тетанус (свыше 25 раздражений); М —
механограмма — запись сокращения мышц; Р — частота раздражений; 1 — фаза
сокращения; 2 — фаза расслабления
Механизм мышечного сокращения (теория скольжения).В состоянии покоя тонкие нити актина лежат частично своими концами в промежутках между толстыми миозиновыми нитями. При возбуждении мышцы под влиянием потенциала действия нити актина сдвигаются в промежутке между нитями миозина, т. е. скользят навстречу друг другу, вызывая сокращение миофибрилл (рис. 4.23). Считают, что скольжение происходит в результате укорочения мостиков — боковых отростков миозиновых нитей под влиянием ионов кальция. Укорачиваясь, мостики тянут актиновую нить навстречу миозиновой.
Работа и сила мышц.При сокращении мышца укорачивается и тем самым совершает работу. Она будет наибольшей при средних нагрузках и среднем ритме сокращений.
Силу мышцы определяют по максимальному сокращению ее при поднятии максимального груза. Наибольшей силой обладают мышцы с косыми волокнами, имеющими перистое строение.
Утомление мышц.В процессе сокращений мышцы утомляются, при этом понижаются их возбудимость, лабильность и величина сокращения. В утомленной мышце понижается содержание гликогена и накапливаются продукты обмена. В опытах на нервно-мышечном препарате Н. Е. Введенский установил, что прежде всего утомляются синапсы в связи с их низкой лабильностью. В целом организме утомление в первую очередь наступает в нервных центрах коры больших полушарий. И. М.Сеченов доказал, что быстрое восстановление работоспособности утомленных мышц наступает не при полном их покое, а при работе других, до этого не сокращавшихся мышц. Импульсы от вновь вовлеченных в работу мышц повышают возбудимость нервных центров, возбуждение одних нервных центров снижает и даже снимает утомление других центров. Утомление зависит от состояния симпатической нервной системы и желез внутренней секреции: утомленная мышца вновь
Рис. 4.23. Схема расслабленной (А) и сократившейся (Б) миофибриллы
 |
начинает сокращаться при раздражении симпатического нерва или введении адреналина, усиливающих обмен. Тренировка мышц позволяет отсрочить наступление утомления. При ней увеличивается объем мышц в результате утолщения мышечных волокон, повышается содержание гликогена, АТФ и креатинфосфата, ускоряются восстановительные процессы.
Тонус мышц. Тонус скелетных мышц играет важную роль для поддержания определенного положения тела в пространстве и деятельности двигательного аппарата. Скелетные мышцы в покое расслабляются не полностью, а находятся в некотором напряжении, т.е. тонусе. Он обусловлен поступлением к мышцам редких нервных импульсов, возбуждающих мышечные волокна не одновременно, а поочередно.
Гладкие мышцы обладают способностью длительно находиться в тонусе. Длительные тонические сокращения этих мышц особенно отчетливо выражены в сфинктерах полых органов, в стенках кровеносных сосудов.
У животных различают три вида мышц: скелетные поперечно- исчерченные, сердечную поперечно-исчерченную (см. гл. 24) и гладкие, которые различны по строению и физиологическим свойствам.
Поперечно-исчерченные скелетные мышцы
Скелетные мышцы вместе со скелетом составляют аппарат движения организма, который обеспечивает поддержание позы и перемещение отдельных частей тела и всего тела в пространстве. Наряду с этим скелетные мышцы и скелет вырабатывают тепловую (70%) и механическую (30%) энергию упругих деформаций и электрическую энергию, выполняют защитную функцию, защищая внутренние органы от повреждений. Масса мышц может составлять около 50% массы тела (у крупного рогатого скота и лошади — 42—47%, у овцы — 34%, у свиньи — 31%). По структурной организации (имеют соединительнотканную строму и мышечную паренхиму разной степени развития) и физиологической роли мышцы делятся на динамические, статодинамические и статические.
Строение. Скелетные мышцы (мышечная ткань, или мышечная паренхима) состоят из мышечных волокон, которые объединяются в мышечные пучки. Мышечное волокно — это специализированная клетка цилиндрической формы, длинной до 10—12 см и диаметром 10—100 мк. Каждое мышечное волокно состоит из оболочки (сарколеммы) и цитоплазмы (саркоплазмы). В саркоплазме располагаются все компоненты животной клетки, вдоль оси мышечного волокна — тонкие нити — миофибриллы, а в них — протофибриллы — нити белков миозина и актина. Они являются сократительным аппаратом мышечного волокна. Миофибриллы объединяются в группы — колонки по 4—20 штук в каждой. Каждая миофибрилла Z-мембранами разделена приблизительно на 20 тыс. саркомеров длиной около
2,5 км. На обоих концах саркомеров к Z-мембране прикреплены около 2 тыс. тонких актиновых нитей, которые своими концами входят между миозиновыми толстыми нитями.
Механизм сокращения. Сокращение мышцы связано с взаимодействием актина и миозина (рис. 16.2). Взаимодействие актина и миозина тормозится системой мышечных белков. На поверхности актиновых нитей имеется два белка — тропонин и тропомиозин. Поступление импульса к мышце сопровождается выходом из саркоплазматического ретикулума мышечного волокна ионов Са 2+ . Они взаимодействуют с белком тропонином, образуют комплекс, он толкает тропомиозин в желобки между двумя цепями актина, у актиновых нитей освобождается поверхность для взаимодействия

Рис. 16.2. Скелетная мышца и ее составные части:
А — мышца; б — часть мышцы, увеличенная, чтобы показать мышечное волокно; В — часть мышечного волокна, увеличенная, чтобы показать миофибрилл; Г — часть миофибрилла, увеличенная, чтобы показать актиновые и миозиновые волокна; Д — часть миофибрилла, увеличенная, чтобы показать саркомер, ограниченный с двух сторон мембранами (Z-линии): / — изотропные участки, образованные актиновыми нитями; А — анизотропные участки, образованные миозиновыми нитями; Н-зона — в центральной части анизотропного участка, актиновые и миозиновые нити не перекрывают друг друга; Е — миозиновые и актиновые протофибриллы; Ж — взаимодействие между миозином и актином при сокращении мышц: а-б — фазы взаимодействия, в-а — отсутствие взаимодействия; Ак — актин, Мз — миозин, Тн — тропонин, Пв — поперечные выступы (головки) специального белка миозиновых нитей
с миозиновыми нитями. За счет гребковых движений головок (специального белка) миозиновых нитей актиновые нити втягиваются на миозиновые, мышца укорачивается. После прекращения поступления нервных импульсов кальциевый насос транспортирует Са 2+ в систему саркоплазматического ретикулума, взаимодействие актиновых и миозиновых нитей прекращается, происходит отсоединение поперечных мостиков миозина от актина, мышца расслабляется.
Деятельность кальциевого насоса энергетически обеспечивается расходом АТФ, что имеет место, как при укорочении, так и расслаблении мышцы. Непосредственным источником энергии для сокращения мышц является АТФ, энергия которой обеспечивает перемещение поперечных мостиков. Расщепление АТФ до АДФ и фосфата с обязательным освобождением энергии — обязательное условие следующего прикрепления поперечного мостика к актину.
Свойства. Поперечно-исчерченным скелетным мышцам присущи основные свойства возбудимых тканей: возбудимость и проводимость, а также в определенной степени упругость, растяжимость, эластичность, пластичность. Возбуждение в белых волокнах распространяется со скоростью 12—15 м/с, в красных — 3—4 м/с.
Возбуждение мышцы внешне проявляется в сокращении. В ответ на одиночное раздражение мышца отвечает одиночным сокращением. Оно осуществляется очень быстро (0,09—0,1 с). В одиночном сокращении различают три фазы: скрытую, укороченую и расслабленую (рис. 16.3).

Рис. 16.3. Одиночное сокращение мышцы:
А — момент раздражения мышцы, а-б— скрытый период, б-в — укорочение, в-г — расслабление, г-д — последовательные эластические колебания
Один мотонейрон через свой аксон в мышце иннервирует несколько мышечных волокон. Один нейрон и иннервируемые им мышечные волокна составляют моторную единицу. Моторные единицы имеют разную возбудимость, вовлекаются в ответную реакцию при разной силе импульсов. Поэтому по мере увеличения силы импульсов сила сокращения мышцы увеличивается.
В естественных условиях к мышце поступает, как правило, не один импульс, а серия импульсов. На серию импульсов мышца отвечает длительным сокращением (рис. 16.4). Оно называется тетани- ческим (или длительным) сокращением. Различают гладкий тетанус, который возникает при частых ритмах раздражения, когда каждый очередной импульс застает мышцу в фазе укорочения в ответ на предыдущий импульс, и зубчатый тетанус, возникающий при редких ритмах раздражения, когда каждый очередной импульс застает мышцу в начале фазы расслабления.
Сокращение мышцы, которое не сопровождается изменением тонуса (напряжения), называется изотоническим. Сокращение

Рис. 16.4. Сокращение скелетной мышцы при разной частоте раздражения:
I — сокращение мышцы; II — отметка частоты раздражения; а — одиночные сокращения, б — зубчатый тетанус, в — гладкий тетанус
мышцы, когда она повышает тонус (напряжение), но не может укорачиваться из-за чрезмерной нагрузки, называется изометрическим.
Поперечно-исчерченные скелетные мышцы относятся к произвольным мышцам, т.е. они сокращаются по воле животного. Основная деятельность скелетных мышц связана с обеспечением позы, устойчивого положения тела и перемещения отдельных частей и организма в целом. Отдельные группы мышц осуществляют свою сократительную деятельность в связи с определенными приспособительными реакциями.
Сила мышц. Мышцы характеризуются определенной силой. Она измеряется по максимальному грузу, который мышца в состоянии поднять, либо по максимальному напряжению, которое она может развить в условиях изометрического сокращения. Одиночное мышечное волокно способно развить напряжение 200 мг. Сила мышцы зависит не от длины мышечных волокон, а от их количества, т.е. от ее поперечного сечения. Сила мышцы с косо расположенными волокнами (статодинамическая мышца) значительно больше, чем сила мышцы той же толщины, но с продольно расположенными волокнами (динамическая мышца).
Отношение максимальной силы мышцы к ее анатомическому поперечнику называется относительной силой.
Отношение максимальной силы мышцы к ее физиологическому поперечнику (поперечный разрез мышцы перпендикулярно ходу ее волокна называется физиологическим поперечником мышцы) называется абсолютной силой.
Мощность мышцы равна произведению мышечной силы на скорость ее укорочения.
Работа мышц. Удержание или перемещение тяжести за счет мышечного сокращения называется работой мышц. Если мышцы обеспечивают позу (определенное положение тела без движения), то это называется статической работой, если движение, то это называется динамической работой. Обе работы дополняют друг друга.
Сокращаясь, мышца действует на кость как на рычаг и производит механическую работу. Величина механической работы определяется как произведение величины груза на расстояние, на которое перемещен груз. Измеряется она в джоулях (Дж).
Величина работы мышцы зависит от величины нагрузки и ритма работы. Увеличение перемещаемого груза увеличивает производимую работу, но до известного предела, после которого величина работы уменьшается. Наиболее производительной оказывается работа, совершаемая при средней нагрузке и при среднем ритме сокращений. При частых или редких сокращениях, а также при слишком большой или малой нагрузках механическая работа мышц снижается. Законы средней нагрузки и среднего ритма мышц были установлены И.М. Сеченовым.
Утомление мышц. Мышцы не могут работать беспрерывно. Длительная работа приводит к снижению их работоспособности.
Временное снижение работоспособности мышц, отмечаемое при длительной работе и исчезающее после отдыха, называется утомлением мышц. Принято различать два вида утомления мышц: истинное и ложное.
При ложном утомлении утомляется не мышца, а особый механизм передачи импульсов с нерва на мышцу в месте их контакта, называемый синапс. В синапсе истощаются резервы медиатора.
При истинном утомлении в мышце происходят следующие процессы:
- а) накапливаются недоокисленные продукты распада питательных веществ вследствие недостаточного поступления кислорода;
- б) истощаются запасы источников энергии для мышечного сокращения.
Утомление проявляется уменьшением силы сокращения мышцы и степени расслабления мышцы.
Если мышца на некоторое время прекращает работу и находится в состоянии покоя, то:
- 1) восстанавливается работа синапса;
- 2) с кровью из мышцы удаляются продукты обмена, а доставляются к ней питательные вещества; мышца вновь приобретает способность сокращаться и производить работу.
Читайте также:
