
Нервный путь тактильной сенсорной системы
Сенсорная (чувствительная) информация играет очень важную роль в жизнедеятельности человека. Она поступает в нервную систему различными путями. Через кожный покров и от органов чувств идет поток внешней (экстероцептивной) информации, сигнализирующий о состоянии внешней среды. От
В передаче и анализе сенсорной информации принимают участие многие нервные структуры. Совокупность всех нервных образований ЦНС и ПНС, осуществляющих восприятие и анализ сенсорной информации, исходящей из внешней и внутренней сред организма, И. П. Павлов назвал анализаторами.
Анализаторы имеют общий план строения (рис. 98). В каждом из них выделяют три отдела, перечисленные ниже.
 |
| Рис. 98. Общий план строения анализатора. |
• Рецепторный отдел, ответственный за опознание специфических раздражителей и преобразование их воздействия в нервное возбуждение. Различают экстерорецепторы (экстероцепторы), воспринимающие раздражения из внешней среды, проприорецепторы (проприоцепторы), воспринимающие раздражения, возникающие в мышцах и суставах, и интерорецепторы (интероцепторы), воспринимающие раздражения от внутренних органов и сосудов.
• Проводниковый отдел, обеспечивающий многоэтапную передачу
нервного возбуждения по соответствующим нервам и трактам через
ряд ядерных (подкорковых) нервных центров. Проводниковый отдел
любого анализатора представлен не только различными ядрами ство
ла мозга и таламуса и их проекциями к соответствующим областям
коры мозга, но и такими образованиями, как ретикулярная форма
ция, структуры лимбической системы, мозжечок, которые принима
ют непосредственное участие в обработке сенсорной информации.
По мере передачи сенсорной информации от одного нервного центра
к другому осуществляется ее последовательный анализ, в результате
чего в организме возникает ощущение, или чувствование.
• Корковый отдел (корковый конец анализатора), находящийся в коре
мозга. Каждый анализатор имеет свою преимущественную локализа
цию в коре мозга. Так, корковое ядро двигательного анализатора
Таким образом, чувствование, а вместе с ним и восприятие ощущения, представляют собой сложные многоэтапные процессы, при реализации которых имеет место функциональное объединение (интеграция) различных структур мозга. На уровне рецепторов происходит опознание раздражений (рецепция), поступающих из внешней среды и внутренней среды организма. По мере проведения сенсорной информации в нервной системе через ряд промежуточных ядерных центров идет ее анализ и перераспределение между различными отделами мозга, т. е. осуществляется само чувствование. Однако ощущение как форма субъективного переживания воспринимаемой сенсорной информации возникает лишь на уровне коры мозга.
Восприятие ощущения как психический процесс субъективного отражения действительности включает не только опознание различных раздражителей и субъективное переживание их воздействий, но и соотнесение их с памятью, эмоциями и другими показателями интегративной деятельности мозга. Однако эта сфера лежит уже за пределами анатомических знаний.
Сенсорная информация от туловища и конечностей по чувствительным волокнам спинномозговых нервов поступает к спинному мозгу, от которого по восходящим путям направляется в головной мозг. При этом восходящие проекционные связи спинного мозга с головным начинаются либо вне спинного мозга от нейронов спинномозговых узлов, либо от нейронов, расположенных в задних столбах спинного мозга. Сенсорная информация от органов головы и частично шеи поступает непосредственно в головной мозг по чувствительным волокнам черепных нервов; при этом восходящие проекционные волокна начинаются в их сенсорных ядрах.
Общей особенностью сенсорных путей является многоэтапная передача возбуждения через различные ядерные центры, в которых происходит последовательный анализ информации. В стволе мозга сенсорные проводящие пути располагаются в его покрышке и, направляясь к коре мозга, обязательно проходят через промежуточный мозг, через его зрительные бугры (таламу-сы), в ядрах которых залегают подкорковые центры всех видов чувствительности, кроме слуховой. В них происходит переключение сенсорных путей; при этом сенсорная информация проходит частичную обработку (анализ и синтез) перед тем, как будет направлена к коре больших полушарий.
Среди сенсорных путей различают: пути протопатической чувствительности (наиболее древней и связанной с передачей сенсорной информации через ядра ретикулярной формации); пути глубокой чувствительности, связанной с передачей проприоцептивной и интероцептивной сенсорной информации; пути поверхностной, или эпикритической, чувствительности, связанной с проведением нервных импульсов, вызванных воздействием осязательного, болевого, температурного раздражителей.
Сенсорные пути специальных видов чувствительности (зрения, слуха, обоняния и вкуса) рассматриваются в гл. 6.
Организм человека содержит большое разнообразие рецепторных клеток воспринимающих воздействия различных факторов окружающей среды. Каждый вид рецепторов обладает специфичностью по отношению к конкретному раздражителю, т. е. воспринимает определенный вид раздражения; иными словами, рецепторы имеют сенсорную модальность. Основные типы рецепторов в организме человека перечислены в табл. 8.
Таблица 8.Характеристика нервных рецепторов*
| Сенсорная модальность | Tun нервного окончания | Локализация рецепторов |
| Экстероцепторы | ||
| • Тактильная (прикосновение) | Тельца Мейснера, тельца Меркеля | Кожа |
| поверхностный слой | ||
| • Болевая (ноцицептивная) | Свободные нервные окончания | поверхностный слой |
| • Давление | Тельца Пачини | средний слой |
| • Температурная | Тельца Руффини, колбы Краузе | глубокий слой |
| Проприоцепторы | ||
| • Растяжение мышц | Нервно-мышечное веретено | Брюшко мышцы |
| • Напряжение мышц | Нервно-сухожильный орган | Сухожилие мышцы |
| • Суставное чувство | Механорецепторы | Суставная капсула, связки |
| Интероцепторы | ||
| • Хеморецепция | Хеморецепторы | Каротидный синус |
| • Осморецепция | Осморецепторы | Гипоталамус |
| • Барорецепция | Барорецепторы, тельца Пачини | Кровеносные сосуды, внутренние органы |
| • Болевая | Ноцицепторы | Внутренние органы |
Характеристика рецепторов специальных видов чувствительности приведена в гл. 6.
Рецепторы, воспринимающие сигналы с поверхности кожного покрова, называют экстероцепторами (экстерорецепторами). К ним относятся тактильные (осязательные) рецепторы, термо- и барорецепторы кожи и слизистых оболочек (рис. 99). В зависимости от конечного пункта назначения — кора мозга или подкорковые образования — сенсорное восприятие, связанное с экстероцепторами, может быть осознанным и бессознательным.
Рецепторы, чувствительные к сигналам, поступающим из различных участков тела (от мышц, суставов, связок и т. д.), называют проприоцептора-ми (проприорецепторами); они поставляют важную соматосенсорную информацию о положении тела в пространстве и взаимном расположении частей тела. К ним относятся различные механорецепторы. Обычно восприятие проприоцептивных раздражений в полной мере человеком не осознается.
Рис. 99. Разновидности экстерорецепторов в кожном покрове.
Рецепторы, расположенные во внутренних органах тела и в сосудах, называют интероцепторами (интерорецепторами).
Практически все рецепторы, кроме своей специфической сенсорной модальности, могут воспринимать болевые сигналы, работая как ноцицепто-ры — болевые рецепторы.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Кожа и слизистые оболочки человека обладают тремя видами чувствительности: тактильной — чувство давления, прикосновения и вибрации; температурной — чувство тепла и холода и болевой (ноцицептивной) — чувство боли. Восприятие всех этих видов чувствительность связано с наличием в коже и слизистых оболочках различных рецепторных образований, которые дают начало нескольким видам соматосенсорной системы: тактильной, температурной, болевой и висцеральной.
В коже и слизистых оболочках располагаются различные рецепторные образования. Наиболее простым типом сенсорных рецепторов являются свободные нервные окончания. Более сложную организацию имеют морфологически дифференцированные образования (осязательные образования Меркеля и Мейснера, тельца Пачини и др.) Большинству специализированных концевых образований присуща предпочтительная чувствительность к определенным видам раздражений, и только свободные нервные окончания воспринимают несколько стимулов, являясь полимодальными рецепторами.
Тактильная сенсорная система. Обеспечивает ощущения прикосновения, давления, вибрации и щекотки.
Периферический отдел. Представлен различными рецепторными образованиями, раздражение которых приводит к формированию специфических ощущений. На поверхности кожи, лишенной волос, а также на слизистых оболочках на прикосновение реагируют специальные рецепторные клетки (тельца Мейснера), расположенные в сосочковом слое кожи и в слизистой оболочке. На коже, покрытой волосами, на прикосновение реагируют рецепторы волосяного фолликула. На давление реагируют рецепторные образования (диски Меркеля), расположенные небольшими группами в глубоких слоях кожи и слизистых оболочек. Адекватным стимулом для них служит прогибание эпидермиса при действии механического стимула на кожу или слизистую оболочку.
Вибрацию — быстрые изменения давления — воспринимают тельца Пачини. Они располагаются в слизистой оболочке и не покрытых волосами частях кожи, в подкожной жировой ткани, а также в суставных сумках, сухожилиях. Тельца Пачини напоминают луковицу. Каждое тельце состоит из многослойной наружной капсулы, внутренней колбы и заключенной в ней части афферентного нервного волокна. В каждое тельце входит одно афферентное волокно. Механический стимул, действуя на тельце Пачини, трансформируется в электрическую энергию рецепторного потенциала, который в нервном волокне вызывает потенциал действия (рис. 7.2.36). Тельца Пачини обладают очень быстрой адаптацией и реагируют на ускорение при смещении кожи.
Щекотание воспринимают свободно лежащие, неинкапсулиро- ванные нервные окончания, расположенные в поверхностных слоях кожи. Для данного вида рецепторов характерна низкая специфичность реакции на стимулы разной интенсивности. С активацией рецепторов этой группы связывают ощущение щекотки, что и дало название самим рецепторам —рецепторы щекотки.

Рис. 7.2.36. Трансформация механического стимула в нервный процесс в тельце Пачини:
РП — рецепторный потенциал; ПД — потенциал действия
Выделяют фазные и статические тактильные рецепторы. Фазные тактильные рецепторы возбуждаются при динамическом раздражении. Они обладают высокой чувствительностью, коротким латентным периодом, быстро адаптируются. Статические тактильные рецепторы возбуждаются, в основном, от статического раздражения. Они менее чувствительны, чем фазные, имеют более длительный латентный период, медленно адаптируются.
Проводниковый отдел. От большинства механорецепторов покровных тканей информация поступает в центральную нервную систему по волокнам группы А и лишь от рецепторов щекотки — по С-волокнам.
Первый нейрон находится в чувствительных ганглиях. Второй нейрон расположен в задних рогах спинного мозга или в ядрах тройничного нерва продолговатого мозга (для челюстно-лицевой области). Оттуда импульсы идут по специфическому и неспецифическому путям в таламус. Специфический путь обеспечивает быструю передачу сигналов в кору о прикосновении и давлении, дифференцированных по силе и месту действия. Афферентные сигналы, идущие по неспецифическому пути, вовлекают в процесс восприятия тактильных стимулов структуры гипоталамуса и лимбического комплекса. Участие этих образований ЦНС привносит вегетативный и эмоциональный компоненты в формирование тактильного восприятия в целом (рис. 7.2.37).
Центральный отдел. Центральный отдел тактильной сенсорной системы локализуется в задней центральной извилине соматосенсорной области коры большого мозга. Центральные анализаторы тактильной системы позволяют распознать пространственное по-

Рис. 7.2.37. Лемнисковый путь тактильной чувствительности
ложение раздраженных рецепторов, а также оценить интенсивность одиночного тактильного стимула.
По порогу тактильная чувствительность бывает пространственной и абсолютной. Под пространственным порогом понимают наименьшее расстояние между двумя точками кожи или слизистой оболочки, при одновременном раздражении которых возникает ощущение двух прикосновений (различительная способность). Наибольшей различительной способностью обладают кончик языка, губы, ладонная поверхность пальцев рук, а наименьшей — голень, спина, бедро, плечо. Пространственный порог определяют методом эстезиометрии с помощью циркуля Вебера.
Нарастание давления в одной точке выше величины нижнего порога тактильного рецептора влечет за собой увеличение интенсивности ощущения до верхнего предела, который является вместе с тем нижним порогом болевых ощущений. Уровень абсолютной тактильной чувствительности определяют эстезиометрически с помощью набора волосков и щетинок разного размера или приборов, строго дозирующих величину механической нагрузки.
Температурная сенсорная система. Она обеспечивает формирование ощущений тепла и холода при действии низких и высоких температурных раздражителей внешней или внутренней среды на организм. Эта система имеет первостепенное значение для поддержания постоянства температуры тела человека, в том числе за счет сложных поведенческих приспособительных реакций.
Периферический отдел. Представлен двумя видами рецепторов: холодовыми (тельца Руффини) и тепловыми (колбы Краузе).
Рецепторы холода расположены в коже и слизистой оболочке поверхностно, а рецепторы тепла находятся более глубоко. Наибольшее число термочувствительных рецепторов сосредоточено в области губ и век. В покровных тканях тепловых рецепторов точек примерно в 10 раз меньше, чем холодовых, а на периферии роговицы и конъюнктивы глаза они вообще отсутствуют.
Свободные нервные окончания, реагирующие на изменение температуры, являются первично-чувствующими полимодалъными рецепторами. Большинство терморецепторов имеют локальные рецептивные поля и реагируют на отклонение температуры повышением частоты генерируемых импульсов, которое наблюдается в течение всего времени действия стимула.
Проводниковый отдел. Афферентные импульсы от рецепторов холода идут по миелинизированным волокнам группы А, а от рецепторов тепла — по немиелинизированным волокнам группы С. С этим связана большая скорость распространения информации от холодовых рецепторов, чем от тепловых. Первый нейрон афферентного звена локализуется в чувствительных ганглиях. Клетки задних рогов спинного мозга или ядра тройничного нерва представляют второй нейрон. Нервные волокна, отходящие от вторых нейронов, в составе спиноталамического тракта доходят до специфических ядер зрительного бугра, где находится третий нейрон. Отсюда возбуждение поступает в четвертые нейроны коры полушарий большого мозга.
Центральный отдел. Центральный отдел температурной сенсорной системы локализуется в области задней центральной извилины коры большого мозга (рис. 7.2.38).
Восприятие температурных раздражителей. В определенном температурном диапазоне человек не испытывает ощущений тепла или холода. Этот диапазон получил название зоны комфорта, или нейтральной зоны. При отклонении температуры кожи за пределы зоны комфорта возникает различные по интенсивности ощущения тепла или холода (рис. 7.2.39).
Различают статические и динамические реакции терморецепторов. Реакция рецепторов на продолжительное действие измененной температуры называется статической. Уровень статической реакции зависит от длительности температурного раздражения и величины отклонения от границы зоны комфорта. При длительном воздействии температурных факторов внешней среды и малых отклонениях температуры кожи происходит медленная частичная адаптация рецепторов с сохранением низкого уровня их статической реакции. При больших отклонениях температуры среды от зоны комфорта адаптивные возможности рецепторов покровных тканей уменьшаются, а уровень статической реакции терморецепторов резко повышается.

Рис. 7.2.38. Проводниковый и корковый отделы температурной сенсорной системы: латеральный и вентральный спиноталамические тракты

Рис. 7.2.39. Температурные зоны реагирования терморецепторов (цит. по Н. Hensel, D.R. Kenshalo, J. Physiol: 204: 99,1969)
Динамические реакции терморецепторов определяются тремя параметрами: исходной температурой, скоростью изменения температуры внешней среды, а также площадью кожи или слизистой оболочки, на которую действует температурный фактор.
Исходная температура кожи определяет уровень возбудимости терморецепторов: чем ниже температура кожи или слизистой оболочки, тем выше возбудимость холодовых и ниже — тепловых рецепторов, и наоборот. При большой скорости изменения температуры внешней среды возбудимость терморецепторов кожи быстро изменяется. При малой скорости изменения температуры среды возбудимость рецепторов изменяется медленно. В этих условиях возможно приспособление к воздействию температурного фактора, проявляющееся в снижении возбудимости терморецепторов (аккомодации).
Между интенсивностью температурных ощущений и размером поверхности, на которую действует температурный стимул, существует прямо пропорциональная зависимость: чем больше площадь воздействия температурного фактора, тем температурные ощущения сильнее, и наоборот. Этот феномен связывают с наличием пространственной суммации на разных уровнях проводникового отдела температурной сенсорной системы. Наличие пространственной суммации подтверждается опытом с двусторонней стимуляцией тыльной поверхности обеих рук. При воздействии на обе руки температурные ощущения будут выше, чем при обогревании или охлаждении одной руки.
Температурную чувствительность исследуют методом термоэсте- зиометрии, путем определения плотности расположения рецепторов, а также их функциональной мобильности.
Болевая сенсорная система. См. модульная ед. 8.3.
Внутренние (висцеральные) сенсорные системы. Осуществляют анализ и синтез информации о состоянии внутренней среды организма и участвует в регуляции работы внутренних органов. Выделяют следующие сенсорные системы:
- • давления в кровеносных сосудах и давления (наполнения) во внутренних полых органах — периферическим отделом этой системы являются механорецепторы;
- • температуры;
- • химического состава внутренней среды организма;
- • осмотического давления внутренней среды.
Рецепторы этих сенсорных систем, имеющие общее название ин- тероцепторы, расположены в различных органах, сосудах, слизистых оболочках и ЦНС.
Периферический отдел. Интероцепторы внутренней сенсорной системы представлены рецепторами, реагирующими на различные виды раздражителей. К механорецепторам относятся все рецепторы, для которых адекватным стимулом является давление, а также растяжение, деформация стенок сосудов, сердца, легких, желудочно- кишечного тракта и других внутренних органов. К хеморецепторам относят всю совокупность рецепторов, реагирующих на химические раздражители. Это рецепторы аортального и каротидного клубочков, слизистых и серозных оболочек пищеварительного тракта и органов дыхания, а также хеморецепторы головного мозга. Осморецепторы расположены в аортальном и каротидном синусах, в артериальных сосудах и вблизи капилляров, а также в печени и других органах. Терморецепторы локализованы в слизистых оболочках пищеварительного тракта, органов дыхания, мочевого пузыря; серозных оболочках; в стенках кровеносных сосудов, а также в ядрах гипоталамуса. Болевые рецепторы (ноцицепторы) широко распространены в органах и тканях, но особенно много их находится в соединительнотканных оболочках, покрывающих внутренние органы, а также в плевре и брюшине.
Проводниковый отдел. Возбуждение, возникшее в интероцепторах, распространяется к нервным центрам по афферентным волокнам соматических и висцеральных нервов (блуждающего, большого, малого чревного, тазового). С этим связано возникновение сома- товисцеральных, висцеро-висцеральных и висцеросоматических рефлексов. Соматические афферентные волокна осуществляют иннервацию строго определенных участков тела и связаны с определенными сегментами спинного мозга, что создает возможность четкой локализации области раздражения и формирования определенных рефлексов. Такая же сегментарная иннервация характерна для плевры и брюшины.
Висцеральные рефлексы могут возникать под влиянием возбуждений, поступающих по соматическим и собственным висцеральным афферентным волокнам.
Организация афферентного звена висцеральных (вегетативных) рефлексов имеет свои особенности. Она характеризуется:
- • наличием прямых (основных, или сегментарных) и непрямых (дополнительных, или надсегментарных) афферентных путей;
- • множественностью и многосегментарностью афферентных путей от внутренних органов;
- • двусторонней воронкой: в одном органе перекрываются зоны иннервации афферентных путей нескольких отделов (сегментов) ЦНС, а в одном отделе (сегменте) ЦНС сходятся афферентные пути от нескольких органов;
- • неодинаковыми зонами иннервации: различные висцеральные нервы иннервируют значительно различающиеся по размеру области. Например, блуждающий нерв иннервирует органы грудной и брюшной полостей вплоть до малого таза, в то время как другой парасимпатический нерв — тазовый — иннервирует органы значительно меньшей области — малого таза.
Главным коллектором чувствительных путей парасимпатической нервной системы служит блуждающий нерв. Афферентные парасимпатические нейроны локализуются преимущественно в верхнем узле блуждающего нерва. Аксоны этих нейронов входят в продолговатый мозг. Другим крупным афферентным парасимпатическим нервом является синусный нерв, в состав которого входят волокна, идущие от рецепторов каротидного клубочка, расположенного у места ветвления общей сонной артерии. Чувствительные волокна синусного нерва это отростки афферентных нейронов верхнего и нижнего яремных узлов. Центральные отростки этих нейронов направляются к чувствительному ядру продолговатого мозга. Афферентные возбуждения от органов и тканей челюстно-лицевой области, участвующие в формировании рефлексов парасимпатической системы, проходят в составе тройничного, лицевого, языкоглоточного нервов и заканчиваются в чувствительном ядре продолговатого мозга, откуда они переключаются на другие парасимпатические структуры ствола мозга.
Обработка афферентных висцеральных возбуждений, поступающих по проводящим путям спинного мозга, происходит в над- сегментарных структурах (рис. 7.2.40. Уровни взаимодействий в автономной нервной системе(?2)). Так, часть аксонов нейронов спинного мозга в составе задних столбов достигает продолговатого мозга, где образует синапсы в соответствующих ядрах. В них расположены вторые нейроны афферентного пути висцерального рефлекса, отростки которых формируют путь из продолговатого мозга в ядра таламуса. Аксоны интернейронов спинного мозга, к которым подходят афферентные волокна от внутренних органов, в составе афферентных трактов также достигают нейронов ядер таламуса.
Потоки возбуждений от внутренних органов по внутрицент- ральным восходящим путям достигают ретикулярной формации среднего мозга. В ретикулярной формации проекции афферентных висцеральных и соматических нервов распределены диффузно. Поэтому от одной и той же точки можно регистрировать ответы, вызванные раздражением не только висцеральных, но и соматических чувствительных нервов. Взаимодействие соматических и висцеральных возбуждений активирует ретикулярные нейроны. Участие ретикулярной формации в регуляции вегетативных процессов опосредуется ее влияниями на симпатический отдел автономной нервной системы, гипоталамические структуры и гипофиз.
По восходящим путям возбуждение достигает ядер гипоталамуса, который служит главным подкорковым центром интеграции висцеральных функций. Он управляет всеми гомеостатическими функциями организма, используя соматические, висцеральные и эндокринные механизмы. Информация об изменениях гомеостаза поступает в гипоталамус по восходящим спинобульбарным путям, а также от рецепторов, контролирующих химический состав, температуру, осмотическое давление крови, протекающей через капилляры гипоталамуса. Висцеральными функциями гипоталамус управляет не прямо, а опосредованно, через центры автономной нервной системы ретикулярной формации ствола и спинного мозга.
Кора больших полушарий мозга — высший интегративный центр регуляции функций организма. В коре головного мозга чувствительные импульсы от внутренних органов проецируются к сенсорным областям и ассоциативным зонам. Представительства симпатических и парасимпатических нервов находятся в близко расположенных зонах коры, что обеспечивает четкую координацию процессов управления висцеральными функциями. Проекции этих нервов также имеют зоны перекрытия на уровне специфических ядер таламуса. На этих уровнях происходит взаимодействие висцеральных и соматических возбуждений, что отражается на процессах восприятия информации от внутренних органов. Например, в нормальных условиях в дневное время более мощный поток соматических возбуждений маскирует возбуждения от висцеральных структур, что не позволяет нам ощущать деятельность своих внутренних органов. В ночное время или при усилении активности интероцепторов в случаях развития нарушений во внутренних органах поток возбуждений от них преобладает над афферентацией от скелетных мышц, что проявляется в неприятных ощущениях в грудной или брюшной полости.
Благодаря тактильной сенсорной системе осуществляется контакт грудного ребенка с матерью, выполнение различных игровых, образовательных и трудовых операций, интимные отношения между мужчиной и женщиной, ощущение комфорта от носимой одежды. В принципе, такие примеры, доказывающие значение тактильной рецепции, можно перечислять еще и еще. Скажем только одно – нарушение этого вида чувствительности существенно снижает адаптационные возможности человека и лишает его многих радостей жизни. Особое место тактильная сенсорная система занимает у млекопитающих, выполняя жизненно важные функции – тактильное исследование окружающей среды, питание, звукообразование и т.д.
Тактильная чувствительность резко повышена на участках тела, которые покрыты волосами. Это объясняется тем, что волосы играют роль рычажков и усиливают передачу раздражения в несколько раз, а так как 95% поверхности человеческого тела покрыто волосами, на некоторых участках едва заметными, то любое прикосновение к поверхности тела резко усиливается.
Различают следующие основные механорецепторы – свободные неинкапсулированные нервные окончания, свободные нервные окончания волосяных фолликулов, диски Меркеля, тельца Руффини, тельца Мейснера и тельца Пачини. Строение указанных рецепторов различно, они распределены неравномерно и локализуются на разной глубине кожи. Первые два типа рецепторов относятся к первичным (являются окончаниями дендрита чувствительного нейрона), остальные – ко вторичным (представляют собой инкапсулированные специализированные клетки, трансформирующие механическое воздействие в рецепторный потенциал, который передается на дендрит чувствительного нейрона). Рассмотрим отдельные виды рецепторов (рис. 4.1).
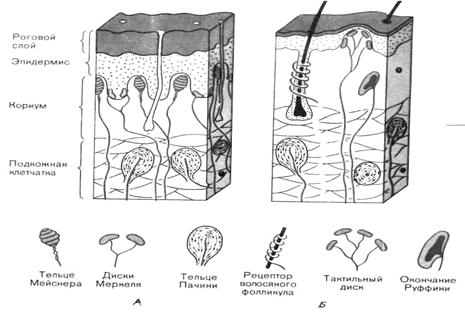
Рис. 4.1. Схема строения и положения механорецепторов в коже, на не покрытых волосами (А) и волосистых (Б) участках кожи.
Свободные неинкапсулированные нервные окончания являются самыми распространенными рецепторами кожи. Они находятся в основном в сосочковом слое дермы – обычно идут вдоль мелких сосудов и представляют собой разветвления дендрита афферентных нейронов. Первоначально их считали рецепторами боли, но в настоящее время их рассматривают как полимодальные рецепторы, отвечающие и на болевые, и на температурные, и на механические стимулы. Это медленно адаптирующиеся рецепторы: продолжают отвечать весь период времени, пока действует стимул.
Свободные нервные окончания волосяных фолликулов также представляют собой разветвления дендрита афферентного нейрона, которые оплетают волосяную сумку. Обычно фолликул получает волокна от нескольких чувствительных нейронов, но в то же время один и тот же дендрит чувствительного нейрона иннервирует несколько волосяных фолликулов. Волос, как было уже отмечено выше, служит рычагом, усиливающим раздражение нервных окончаний, что и объясняет высокую чувствительность волос к прикосновению. Указанные рецепторы реагируют преимущественно на легкое прикосновение и осуществляют пространственное и временное тактильное различение.
Диски (клеточные комплексы) Меркеля представляют собой видоизмененные эпителиальные клетки, с которыми образуют синапсы дендриты афферентных нейронов. Находятся в базальном слое эпидермиса и частично в сосочковом слое дермы в виде небольших скоплений. Особенно много дисков Меркеля в участках кожи с высокой чувствительностью, т.е. в голой коже пальцев рук и на губах. В покрытой волосами коже они лежат в особых колоколообразных тельцах, выдающихся над окружающей кожной поверхностью (в каждом таком образовании, называемом тактильным тельцем Пинкуса-Игго, находится 30-50 клеток Меркеля, связанных с афферентными нервными волокнами). При высоте около 0,1 мм и диаметре 0,2-0,4 мм эти тельца едва различимы невооруженным глазом.
Поскольку диски Меркеля медленно адаптирующиеся рецепторы, то их рассматривают в качестве пропорционального датчика, т.е. генерация рецепторного потенциала в них тем активнее, чем сильнее действует раздражитель. Эти рецепторы рассматривают как рецепторы давления, или силы, поскольку они воспринимают изменение силы механического воздействия.
Тельца (окончания) Руффини располагаются в волосистой части кожи – в глубоких слоях эпидермиса и в сосочковом слое дермы. Представляют собой веретеновидной формы капсулу, образованную плотно переплетенными коллагеновыми волокнами. Внутри такой капсулы содержится жидкость, в которую погружены разветвления дендрита чувствительного нейрона. Как и диски Меркеля, являются рецепторами давления, или силы.
Тельца Мейснера (тактильное мейснеровое тельце) представляют собой капсулу конусовидной или овальной формы. Капсула ориентирована перпендикулярно поверхности кожи. Ее стенки образованы многочисленными пластинчатыми клетками, между которыми параллельно поверхности кожи располагаются терминали дендрита чувствительного нейрона. Эти рецепторы находятся в сосочках собственно кориума, в участках, лишенных волос (кожа ладоней, подошв, пальцев рук и ног, а также губы, сосочки молочных желез и половые органы). В покрытой волосами коже их меньше и здесь они принимают форму рецепторов волосяных фолликулов. Реагируют на скорость изменения силы, т.е. являются датчиком скорости.
Тельца Пачини (пластинчатые тельца, тельца Фатера-Пачини) – наиболее крупные и наиболее распространенные в организме рецепторы, а потому они являются наиболее изученными рецепторами кожи. Располагаются в гиподерме и частично в глубоких слоях дермы. Кроме того, они обнаружены и в сухожилиях и фасциях мышц, в надкостнице, в суставных сумках. Тельце Пачини напоминает луковицу, состоит из наружной капсулы, внутренней колбы и заключенного в нее дендрита афферентного нейрона. Пространство между наружной капсулой и внутренней колбой, а также внутри колбы заполнено жидкостью (ликвором). Возбуждение этих рецепторов происходит при кратковременном (в течение 0,1 мс) смещении капсулы на 0,5 мкм.
Тельца Пачини считаются датчиком ускорения. Благодаря своим свойствам они воспринимают самую незначительную деформацию кожи при соприкосновении с различными предметами и почвой, т.е. с какой скоростью смещается кожа.
Таким образом, большинство тактильных рецепторов локализовано, как правило, в глубоких слоях эпидермиса и сосочковом слое кориума.
Всех их можно разделить на три главных типа:
- рецепторы давления (датчики силы), которые ведут себя как пропорциональные датчики, т.е. генерация рецепторного потенциала в них тем активнее, чем сильнее действует раздражитель. Поэтому их еще называют пропорциональными рецепторами. Это свободные неинкапсулированные нервные окончания, диски Меркля, тельца Руффини;
- рецепторы прикосновения (датчики скорости) реагируют на скорость изменения силы, т.е. скорость вдавления стимула, поэтому осуществляют пространственное и временное тактильное различение. Это тельца Мейсмера и рецепторы волосяных фолликулов;
- рецепторы вибрации (датчики ускорения) – тельца Пачини - реагируют на изменение скорости механического воздействия.
Однако при этом необходимо подчеркнуть, что механические стимулы, обычно действующие на кожу, одновременно возбуждают в той или иной степени несколько типов механорецепторов в зависимости от характера стимула. Поэтому возникающие ощущения нельзя приписать рецепторам одного типа. Соответственно в повседневной жизни трудно определить различия между ощущениями давления и прикосновения.
Принцип работы механорецепторов . Независимо от того, что одни рецепторы реагируют на изменение силы, вторые – на скорость изменения этой силы, а третьи – на вторую производную изменения силы, во всех случаях принцип работы рецептора состоит в том, что под влиянием механического стимула в мембране рецептора изменяется ионная проницаемость, что приводит к появлению рецепторного потенциала. Он вызывает выделение медиатора, что сопровождается появлением генераторного потенциала в дендритах чувствительного нейрона, благодаря которому изменяется частота генерации потенциалов действия.
Различия в восприятии механических стимулов определяются скоростью адаптации тактильных рецепторов. Так, быстро адаптирующиеся рецепторы являются датчиками скорости, или ускорения, а медленно адаптирующиеся – датчиками изменения силы. В то же время скорость адаптации определяется структурой рецепторов – наличие сложно организованной капсулы рецептора повышает скорость его адаптации (укорачивает рецепторный потенциал), поскольку такая капсула хорошо проводит быстрые и гасит медленные изменения давления. Адаптация механорецепторов кожи имеет большое значение – благодаря этому свойству рецепторов мы перестаем ощущать постоянное давление одежды, привыкаем носить на роговице глаз контактные линзы, а на носу – очки и т.д.
Сенсорная информация от механорецепторов кожи передается в кору больших полушарий по нервным волокнам тонкого и клиновидного пучков, образующих лемнисковую систему. Тонкий пучок, или пучок Голля, несет информацию от рецепторов нижних конечностей и нижней части туловища, а клиновидный пучок, или Бурдаха, - от рецепторов верхних конечностей и верхней части туловища. Оба пути состоят из афферентных нейронов (они находятся в спинномозговых узлах), аксоны которых вступают в задние рога спинного мозга и, не прерываясь, направляются в составе задних столбов до тонкого (ядро Голля) и клиновидного (ядро Бурдаха) ядер продолговатого мозга.
Аксоны нейронов этих ядер полностью перекрещиваются на уровне олив, образуя медиальный лемниск (петлю), и, поднимаясь дальше, заканчиваются в специфических ядрах таламуса, которые из-за своего анатомического положения называют вентробазальным комплексом. Нейроны специфических ядер таламуса посылают свои аксоны в соматосенсорные зоны коры больших полушарий (SI и SII). Поражение задних столбов клинически проявляется в потере мышечно-суставного чувства, вибрационной и тактильной чувствительности на стороне поражения при сохранении болевой и температурной чувствительности.
Лемнисковая система проводит точную (по силе и месту воздействия) и сложную (о давлении, прикосновении, вибрации и движении в суставах) информацию с большой скоростью (до 80 км/с).
Для всех составляющих лемнисковой системы (задних столбов спинного мозга, тонкого и клиновидного ядер, таламических ядер и корковых областей) выявлена четкая топографическая организация проекции кожи, т.е. каждый участок кожи контрлатеральной части тела занимает определенную зону, площадь которой зависит от функционального значения этой части тела.
Часть импульсов от рецепторов давления кожи и слизистой передается в кору больших полушарий и по вентральному спиноталамическому тракту, который проходит в составе передних столбов спинного мозга. Однако в спиноталамической системе на уровне таламических ядер правильная соматотопическая организация представительства поверхности тела отсутствует.
Особое место в передаче сенсорной информации от механорецепторов отводится V паре черепно-мозговых нервов - тройничному нерву, который в своих трех ветвях содержит афференты, идущие от лица и полости рта. Он иннервирует кожу, зубы, слизистую полости рта, язык и роговицу. Ко времени рождения тройничная система уже хорошо развита и проводит сенсорные сигналы, которые запускают пищевое поведение. Именно она обеспечивает первое сенсорное знакомство новорожденного с окружающим миром.
Информация от нейронов специфических ядер таламуса первоначально поступает в две проекционные соматосенсорные зоны коры больших полушарий (SI и SII). В частности, информация от нейронов вентробазального комплекса направляется (контрлатерально) в первую проекционную зону, которая у приматов и человека находится в постцентральной извилине (SI). От нейронов задней группы ядер таламуса информация преимущественно поступает (контрлатерально и ипсилатерально) во вторую проекционную соматосенсорную зону коры (SII), которая расположена в области сильвиевой (латеральной) борозды (рядом со слуховой зоной). От этих двух проекционных соматосенсорных зон информация поступает в передние и задние ассоциативные зоны коры.
Первая проекционная соматосенсорная область, локализованная в постцентральной извилине (первичные зоны — 1-е и 3-е поля по Бродману, вторичные зоны — 2-е и 5-е поля), является, по сути, местом окончания лемнискового пути и ядром тактильного анализатора. Она отличается от других областей коры очень высокой степенью топографической организации (проекции различных областей поверхности тела на соматосенсорную кору осуществляются по принципу "точка в точку"). Такое явление называют соматотопией, или топографическим представительством. О наличии соматотопии свидетельствует тот факт, что при раздражении поверхности кожи короткими точечными прикосновениями первичный ответ в соматосенсорной коре локализуется в строго ограниченном участке.
Показано, что размеры представительства соответствующих участков поверхности кожи у человека и приматов в постцентральной извилине связаны не с величиной поверхности их тела, а зависят от биологической значимости информации, воспринимающейся тем или иным участком кожи. Например, у человека представительство губ, лица и кистей рук в этой зоне по площади намного больше, чем представительство туловища и нижних конечностей. Это дает специфический рисунок сенсорного гомункулюса — отражения тела человека в постцентральной извилине (рис. 4.2). Характер этого рисунка свидетельствует о высокой чувствительности и тонком различении для этих частей тела, а также их биологической значимости. Так, при осмотре какого-либо предмета, когда нам надо выяснить форму, наличие шероховатостей и т.д., мы поглаживаем этот предмет, т.е. касаемся его поверхности кожей нашей ладони.
Когда кора в области SI разрушена повреждением или удалена с терапевтической целью, возникает дефицит восприятия. Стимуляция кожи еще может быть воспринята как таковая, но способность локализовать ее и распознать пространственные детали раздражителя нарушается. Так, например, больные не могли на ощупь отличить овал от параллелепипеда (в клинике это нарушение носит название астерогнозис). Степень дефицита зависит от размеров поврежденного участка коры. Здесь снова выступает соматотопическая организация. Однако надо сказать, что после достаточно долгого промежутка времени подобные нарушения становятся слабее. Такое улучшение, по-видимому, обусловлено способностью других областей коры (например, 5-го поля теменной коры, соседнего с SI) принять на себя функции SI.
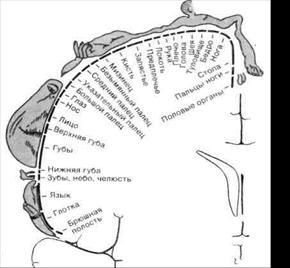
Рис.4.2. Схема чувствительного гомункулюса (соматотопическая организация соматосенсорной коры SI человека).
Разрез полушарий (на уровне постцентральной извилины) во фронтальной плоскости. Обозначения показывают пространственное представительство поверхности тела в коре, установленное на основании локальной стимуляции мозга бодрствующих больных.
В целом роль соматосенсорной зоны коры состоит в интегральной оценке соматосенсорных сигналов, во включении их в сферу сознания, полисенсорный синтез и в сенсорное обеспечение выработки новых двигательных навыков. Удаление или повреждение соматосенсорных зон коры приводит к нарушению способности локализовать тактильные ощущения, а их электростимуляция вызывает ощущение давления, прикосновения, вибрации и зуда.
Читайте также:
