
Носители информации нервный импульс и медиаторы
Но во многих учебниках авторы как раз любят употреблять это загадочное выражение: "обработка информации". Жаль, что оно почти ни о чём не говорит студентам.
Поэтому я решил дать здесь краткий, но очень важный материал о реальных носителях информации в нервной системе.
И первое, с чего надо начать, говоря о работе нервной системы и её структур, - это перейти от понятия "информация" к более конкретному понятию "возбуждение".
С точки зрения физиологии при описании работы нервной системы следует везде, где только это возможно, заменять абстрактный термин "информация" на термин, характеризующий вполне материальный физиологический процесс: "нервное возбуждение". Описание физиологических механизмов психических процессов - это, как правило, описание пути движения нервного возбуждения по нервным структурам. Например, именно таким способом описываются механизмы рефлексов через работу их рефлекторных дуг ( Рефлексы ).
Однако популярное в последнее время высказывание о том, что "мысль материальна" - в корне не верно! Мысль - идеальна, это один из вариантов существования информации. Но вот носители мысли - безусловно, материальны! Это нервные структуры и нервные процессы. Идеальный информационный процесс мышления идёт на материальных носителях .
Материальные носители информации в нервной системе:
Локальные электрические потенциалы: ВПСП, ТПСП, дендритные потенциалы, ГП (генераторные потенциалы) и РП (рецепторные потенциалы), а также следовые потенциалы после прохождения нервного импульса.
Выбросы нейротрансмиттеров (медиаторов и модуляторов).
Мембранные структуры нервных клеток (молекулярные рецепторы, ионные каналы).
Внутренние структуры цитоплазмы и ядра нервных клеток: ДНК и РНК (нуклеиновые кислоты) и белки.
Сохранять информацию можно только на одном из таких носителей, никаких прочих "ячеек памяти" или "ячеек хранения информации" в нервной системе не обнаружено.
Другим вариантом сохранения информации будет изменение структуры самого носителя информации, существующее более длительно и статично. Например, структуры мембраны могут измениться на короткий или длительный срок, или внутриклеточные структуры, включая ядерные, могут измениться, что приведёт к изменению свойств клетки. Это и будет сохранением информации.
Такое изменение клеточных структур под влиянием полученной информации называется пластичностью, и оно лежит в основе важнейших психических процессов.
Пластичность нервных клеток лежит в основе следующих явлений:
Память, научение, условные рефлексы и доминанта – это пластические перестройки синапсов.
Информация сохраняется (фиксируется, запоминается) в изменённых нейронных связях, а не в каком-то особом "хранилище информации".
Этапы обработки информации (транформации возбуждения)
Ввод информации в нервную систему осуществляется с помощью раздражения рецепторов. Итак, первый этап - это раздражение.
Рецепторы производят трансдукцию – преобразование (транформацию) раздражения в нервное возбуждение. Трансдукция — это первая трансформация информации в нервной системе, первая её обработка. Заметьте, что часть информации при этом неизбежно теряется. Этот процесс также можно назвать восприятием, рецепцией, кодированием, т.к. раздражение воспринимается и переводится в форму нервных импульсов, т.е. во внутренний код нервной системы. Подробности смотрите тут: Физиология восприятия , восприятие , кодирование
Возбуждение, порождённое (генерированное) рецепторами, доставляется по афферентным нейронам в низший нервный центр. Это всего лишь проведение возбуждения, без его переработки.
В низшем нервном центре происходит вторая трансформация информации. Возбуждение видоизменяется и разделяется на несколько потоков. Подробности смотрите тут: пути сенсорного возбуждения
Потоки возбуждения, выходящие из низших центров:
а) сенсорно-перцептивный - в вышележащий центр и далее в высший нервный центр коры для создания сенсорного образа и его анализа,
б) модулирующий - в модулирующую (активирующую и тормозящую) систему мозга для изменения активности мозговых структур,
в) эффекторно-рефлекторный - на эффекторы для ответной реакции (рефлекса).
5. В высшем нервном центре происходит третья трансформация информации. Пришедшее в него сенсорное возбуждение преобразуется и разделяется на множество потоков. Формируется перцептивный сенсорный образ раздражителя.
Изучаемые вопросы:
Тема 9.2 – нервная регуляция процессов жизнедеятельности.
Расписание: 6 часа теория + 6 часа практика
Раздел: А Классификация нервной системы
Общие принципы строения ЦНС
1.Классификация нервной системы
1. Общие принципы строения ЦНС:
3. Виды нейронов:
- по локализации - по функции
6. Нервный центр-понятие
7. Виды нервных волокон
8. Нервы - строение, виды.
10. Виды синапсов:
- по виду контакта
- по способу передачи сигналов
11. Виды химических синапсов:
12. Механизмы передачи в синапсах
Раздел: Б Спинной мозг
2. Оболочки спинного мозга
3. Локализация чувствительных нейронов
4. Сегмент-понятие виды
5. Зоны Захарьина-Геда
6. Спинномозговые корешки:
8. Виды рефлексов
9. Нервная деятельность:
-виды (высшая и низшая)
-структуры ее осуществляющие
10. Универсальные процессы нервной деятельности:
-носители информации (нервный импульс и медиаторы)
-принцип нервной деятельности
11. Интегративный характер нервной деятельности и его структурно- функциональные основы:
12. - Проводящие пути спинного мозга:
13. Нервные центры спинного мозга
14. Функции спинного мозга:
15. Рефлексы спинного мозга:
16. Рефлекторна дуга как система нейронов и их отростков, контактирующих посредством синапсов.
17. Рефлекторные дуги простых и сложных соматических рефлексов спинного мозга (сухожильных и кожно-мышечных)
Раздел В. Спинномозговые нервы
4. Нервные волокна, их образующие:
-чувствительные - дендриты чувствительных нейронов спинальных ганглиев
-двигательные - аксоны двигательных нейронов спинного мозга
-вегетативные- аксоны вегетативных нейронов спинного мозга
5. Ветви спинномозговых нервов - функциональные виды нервных волокон, идущих в их составе.
6. Серая соединительная ветвь
7. Грудные спинномозговые нервы.
8. Сплетение передних ветвей спинномозговых нервов
- шейное
Плечевое
Поясничное
Крестцово-копчиковое
9. Нервные стволы
10.Области иннервации, сплетений
Раздел Г. Головной мозг
3. Продолговатый мозг:
4. Мост
5. Мозжечок
- внешнее и внутреннее строение,
6. Четверохолмие
-и нижние бугры, их микроструктура.
7. Промежуточный мозг;
структуры его образующие:
8. Ствол мозга:
9. Ретикулярная формация:
10. Лимбическая система:
-интеграция эмоций и вегетативных реакций
11. Проводящие пути головного мозга
12. Конечный мозг
- внешнее и внутреннее строение
13. Базальные ядра
Кора головного мозга.
14. Проекционные зоны коры
15. Ассоциативные поля, их функции
16. Послойное строение коры
17. Экранный принцип функционирования коры.
18. Условные рефлексы.
19.Условно-рефлекторная деятельность коры
Оболочки головного мозга и межоболочечные пространаства
Полости головного мозга
-желудочки их сообщение друг с другом
- со спинномозговым каналом,
- субарахноидальным пространством головного и спинного мозга.
Ликвор
-состав, образование, движение, функции
Черепные нервы
-Функциональные виды черепных нервов
-и парасимпатических волокон черепных нервов
Обонятельные нервы
-выход из полости носа в полость черепа
Место контакта с обонятельным мозгом. Функция
Зрительный нерв
-выход из полости глазницы в полость мозга.
Глазодвигательный нерв
-выход из полости глазницы в полость мозга, области иннервации.
Блоковой нерв
-выход из полости глазницы в полость мозга, области иннервации.
Отводящий нерв
-выход из полости глазницы в полость мозга, области иннервации.
Тройничный нерв
-место выхода из полости черепа
-области иннервации: чувствительных, двигательных и парасимпатических волокон 1-й, 2-й и 3-й ветвей
Лицевой нерв
-расположение в височной кости
-место выхода из полости черепа
Преддверно-улитковый нерв
Языко-глоточный нерв
-место выхода из полости черепа
-области иннервации чувствительных волокон
Блуждающий нерв
-место выхода из полости черепа
-области иннервации двигательных чувствительных и парасимпатических волокон
Добавочный нерв
-место выхода из полости черепа
-вид его волокон
Подъязычный нерв
–место выхода из полости черепа.
Раздел Д. Классификация вегетативной нервной системы
1. Область иннервации и функции вегетативной нервной системы
2. Центральные и периферические отделы вегетативной нервной системы
3. Отличия: вегетативной нервной системы от соматической, симпатической от парасимпатической
4. Симпатические стволы и нервные сплетения.
5. Вегетативная рефлекторная дуга
6. Медиаторы в синапсах
7. Влияние симпатической и парасимпатической нервной системы на:
-секрецию бронхиальных желез
-секрецию пищеварительного тракта
Секрецию потовых желез
Раздел Е. Понятие о высшей нервной деятельности.
2. Условные рефлексы
3. Принципы рефлекторной теории И.П. Павлова
4. Особенности образования условных рефлексов, механизмы
5. Виды условных рефлексов
6. Торможение условных рефлексов
7. Структурно-функциональные основы особенностей психической деятельности человека (1 и 2 сигнальные системы)
8. Физиологические основы индивидуальной психической деятельности
9. Типы высшей нервной деятельности человека
10. Формы психической деятельности:
- ( физиологические основы перечисленных форм психической деятельности)
Глава 14 ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Дата добавления: 2018-06-01 ; просмотров: 231 ;
Связь миллиардов нейронов мозга осуществляется посредством медиаторов. Химическое вещество можно отнести к числу медиаторов лишь в том случае, если оно удовлетворяет ряду критериев. В нервных волокнах должны содержаться ферменты, необходимые для синтеза этого вещества. При раздражении нервов это вещество должно выделяться, реагировать со специфическим рецептором на постсинаптической клетке и вызывать биологическую реакцию. Должны существовать механизмы, быстро прекращающие действие этого вещества.
Всем этим критериям удовлетворяют два вещества – ацетилхолин и но-радреналин. Содержащие их нервы называют соответственно холинерги-ческими и адренергическими. В соответствии с этим все эфферентные системы делят на холинорецепторы и адренорецепторы.
Ряд других химических веществ удовлетворяют многим, но не всем перечисленным критериям. К таким медиаторам относят дофамин, адреналин, серотонин, октопамин, гистамин, ГАМК и др.
Обширная группа холинорецепторов весьма неоднородна как в структурном, так и в функциональном отношении. Объединяют их медиатор ацетилхолин и общая схема строения синапса.
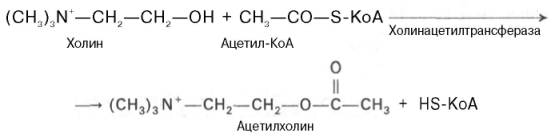
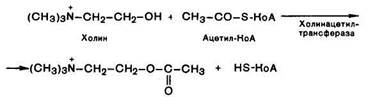
Ацетилхолин представляет собой сложный эфир уксусной кислоты и холина. Он синтезируется в нервной клетке из холина и активной формы ацетата – ацетилкоэнзима А при помощи специального фермента холин-ацетилтрансферазы (холинацетилазы):
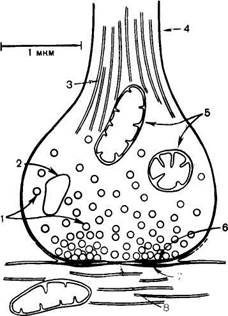
Синапс можно представить себе как узкое пространство (щель), ограниченное с одной стороны пресинаптической, а с другой – постсинапти-ческой мембраной (рис. 19.4). Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других истончена и имеет отверстия для сообщения цитоплазмы аксона с синаптическим пространством. Постсинапти-ческая мембрана менее плотная, не имеет отверстий. Подобным образом построены и нервно-мышечные синапсы, но они имеют более сложное строение мембранного комплекса.
В общих чертах картину участия ацетилхолина в осуществлении передачи нервного импульса возбуждения можно представить следующим образом. В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30–80 нм, которые содержат нейромедиаторы. Эти пузырьки покрыты оболочкой, которая образована белком клатрином (мол. масса 180000). В холинергических синапсах каждый пузырек диаметром 80 нм содержит
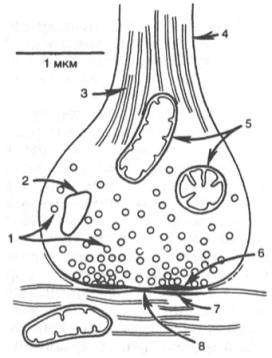
Рис. 19.4. Схематическое изображение синапса (по Мецлеру).
1 - синаптические пузырьки; 2 - лизосо-ма; 3 - микрофибриллы (нейрофибрил-лы); 4 - аксон; 5 - митохондрии; 6 - пре-синаптическое утолщение мембраны; 7 - постсинаптическое утолщение мембраны; 8 - синаптическая щель (около 20 нм).
В холинергических синапсах это происходит двумя путями. Первый путь заключается в том, что ацетилхолин подвергается ферментативному гидролизу. Второй путь – это энергозависимый активный транспорт ацетилхо-лина в нейрон, где он накапливается для последующего повторного использования.
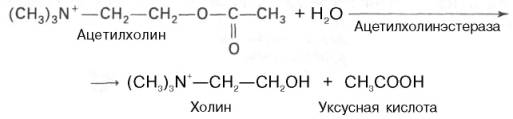
В большинстве отделов головного мозга гидролиз ацетилхолина осуществляется ацетилхолинэстеразой (истинная холинэстераза, которая гид-ролизует ацетилхолин быстрее, чем иные эфиры холина). В нервной ткани существуют и другие эстеразы, которые способны гидролизовать ацетил-холин, но значительно медленнее, чем, например, бутирилхолин. Эти эстеразы называются холинэстеразой (или псевдохолинэстеразой). К числу холинергических систем относятся моторные нейроны, образующие нервно-мышечные соединения, все преганглионарные нейроны автономной нервной системы и постганглионарные нейроны парасимпатической нервной системы. Большое количество холинергических симпатических областей обнаружено также в головном мозге. В зависимости от чувствительности к той или иной группе химических соединений холинергические нейроны делятся на мускариновые (активируемые мускарином) и никотиновые (активируемые никотином). Мускариновые рецепторы ацетилхо-лина, имеющиеся во многих нейронах автономной нервной системы, специфически блокируются атропином. Никотиновые синапсы присутствуют в ганглиях и скелетных мышцах. Их ингибиторами являются кураре и активный компонент этого яда D-тубокурарин.
Необходимо подчеркнуть, что в адренорецепторах существует два вида рецепторов для норадреналина: α- и β-адренергические рецепторы. Эти рецепторы можно отличить друг от друга по специфическим реакциям, которые они вызывают, а также по тем специфическим агентам, которые способны блокировать данные реакции.
Установлено, что как только β-адренергический рецептор, расположенный на наружной поверхности мембраны эффекторной клетки, начинает взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Этот сложный ряд последовательных реакций может быть заблокирован пропраноло-лом – веществом, препятствующим связыванию норадреналина с β-адре-нергическим рецептором.
Известно, что в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (—NH2) у норадреналина, серотонина, дофамина и адреналина, тем самым инактивируя указанные медиаторы. В последние годы было показано, что, помимо ферментативного превращения, существует и другой механизм быстрой инактивации, точнее удаления, медиаторов. Оказалось, что норадреналин быстро исчезает из синаптической щели в результате вторичного поглощения симпатическими нервами; вновь оказавшись в нервном волокне, медиатор, естественно, не может воздействовать на постсинаптические клетки. Конкретный механизм этого явления пока не вполне ясен.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности использующими серотонин в качестве медиатора. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами. Исследования, проведенные с ингибитором синтеза серотонина n-хлорфенилаланином, а также с другими ингибиторами, дают основания считать, что серотонин влияет на процессы сна. Выявлено также, что торможение кортикостероидами секреторной активности гипофиза оказывается менее эффективным у тех животных, мозг которых беднее серотонином.
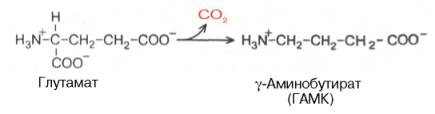
Важным нейромедиатором, выполняющим тормозные функции, является γ-аминомасляная кислота (ГАМК), количество которой в головном мозге во много раз больше, чем других нейромедиаторов. Так, в гипоталамусе суммарное содержание ацетилхолина, норадреналина, дофамина и серотонина не превышает 10 мкг/г, в то время как ГАМК в этом отделе головного мозга более 600 мкг/г. ГАМК увеличивает проницаемость постсинаптических мембран для ионов К + и тем самым отдаляет мембранный потенциал от порогового уровня, при котором возникает потенциал действия; таким образом, ГАМК – это тормозной нейромедиатор. ГАМК образуется при декарбоксилировании глутамата в реакции, катализируемой глутаматдекарбоксилазой:
В терапевтической практике применяется большое количество лекарственных средств, которые действуют через систему медиаторов. Многие лекарственные препараты, успешно применяемые при лечении гипертонии, влияют на накопление и выделение адренергических медиаторов. Например, резерпин – понижающее артериальное давление средство специфически тормозит процесс переноса катехоламинов в специальные гранулы нейронов и тем самым делает эти амины доступными действию эндогенной МАО.
Многие антидепрессанты (вещества, снимающие депрессию) увеличивают содержание катехоламинов в синаптической щели, т.е. количество медиаторов для стимулирования рецептора возрастает. К таким веществам, в частности, относятся имипрамин (блокирует поглощение норадреналина нервными волокнами), амфетамин (одновременно способствует выделению норадреналина и блокирует его поглощение), ингибиторы МАО (подавляют метаболизм катехоламинов) и др. В связи с этим возникла катехол-аминовая гипотеза депрессивных состояний, согласно которой психическая депрессия связана с недостатком катехоламинов в мозге.
В начале 50-х годов фармакологи выяснили, что известный галлюциноген диэтиламин лизергиновой кислоты (ЛСД) не только сходен по химическому строению с серотонином, но и нейтрализует некоторые его фармакологические эффекты (блокируя рецепторы серотонина). Поэтому было высказано предположение, что нарушение обмена серотонина может быть причиной возникновения особых психических заболеваний.
Считают, что такие антипсихотические средства, как аминазин (хлор-промазин) и галоперидол, усиливая синтез катехоламинов, способны блокировать дофаминовые рецепторы в мозге.
Связь миллиардов нейронов мозга осуществляется посредством медиаторов — ацетилхолин и норадреналин. Содержащие их нервы называют соответственно холинергическими и адренергическими. В соответствии с этим все эфферентные системы делят на холинорецепторы и адренорецепторы.
К другим медиаторам относятся дофамин, адреналин, серотонин, октопамин, гистамин, ГАМК и др.
Обширная группа холинорецепторов весьма неоднородна как в структурном, так и в функциональном отношении. Объединяют их медиатор — ацетилхолин — и общая схема строения синапса.
 |
Ацетилхолин представляет собой сложный эфир уксусной кислоты и холина. Он синтезируется в нервной клетке из холина и активной формы ацетата — ацетилкоэнзима А при помощи специального фермента холинацетилтрансферазы (холйнацетилазы):
| Схематическое изображение синапса (по Мецлеру). 1 — синаптические пузырьки; 2 — лизосома; 3 — микрофибриллы (нейрофибриллы); 4 - аксон; 5 - митохондрии; 6 — пресинаптическое утолщение мембраны; 7 - постсинаптическое утолщение мембраны; 8 — синаптическая щель (около 20 нм). |
Синапс можно представить себе как узкое пространство (щель), ограниченное с одной стороны пресинаптической, с другой — постсинаптической мембраной. Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других — истончена и имеет отверстия, с помощью которых цитоплазма аксона может сообщаться с синаптическим пространством. Постсинаптическая мембрана менее плотная, не имеет отверстий.
В адренорецепторах существует два вида рецепторов для норадреналина: α- и β-адренергические рецепторы.
Как только β-адренергический рецептор (расположенный на наружной поверхности мембраны эффекторной клетки) начнет взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Известно, что в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (-NH2) у норадреналина, серотонйна, дофамина и адреналина, тем самым инактивируя упомянутые медиаторы.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности использующими серотонин в качестве медиатора. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами.
Важным нейромедиатором, выполняющим тормозные функции, является ГАМК, количество которой в головном мозге во много раз выше, чем других нейромедиаторов.
Синапс
Как передается возбуждение от одного нейрона другому или от нейрона, например, на мышечное волокно? Этой проблемой интересуются не только профессиональные нейробиологи, но и врачи, особенно фармакологи. Знание биологических механизмов необходимо для лечения некоторых заболеваний, а также для создания новых лекарств и препаратов. Дело в том, что одними из основных мест воздействия этих веществ на организм человека являются места передачи возбуждения с одного нейрона на другой (или на другую клетку, например клетку сердечной мышцы, стенки сосудов и пр.). Отросток нейрона аксон направляется к другому нейрону и образует на нем контакт, который называют синапсом (в переводе с греческого - контакт; см. рис. 2.3). Именно синапс хранит многие тайны мозга. Нарушение этого контакта, например, веществами, блокирующими его работу, приводит к тяжелейшим последствиям для человека. Это место приложения действия наркотиков. Примеры будут приведены ниже, а сейчас рассмотрим, как устроен и как работает синапс.
Трудности этого исследования определяются тем, что сам синапс очень маленький (его диаметр не более 1 мкм). Один нейрон получает такие контакты, как правило, от нескольких тысяч (3 - 10 тыс.) других нейронов. Каждый синапс надежно закрыт специальными клетками глии, поэтому исследовать его очень непросто. На рис. 2.12 показана схема синапса, как это представляет себе современная наука. Несмотря на свою миниатюрность, он устроен весьма сложно. Одним из его основных компонентов являются пузырьки, которые находятся внутри синапса. Эти пузырьки содержат биологически очень активное вещество, которое называется нейротрансмиттером, или медиатором (передатчиком).
Вспомним, что нервный импульс (возбуждение) с огромной скоростью продвигается по волокну и подходит к синапсу. Этот потенциал действия вызывает деполяризацию мембраны синапса (рис. 2.13), однако это не приводит к генерации нового возбуждения (потенциала действия), а вызывает открывание специальных ионных каналов, с которыми мы еще не знакомы. Эти каналы пропускают ионы кальция внутрь синапса. Ионы кальция играют очень большую роль в деятельности организма. Специальная железа внутренней секреции - паращитовидная (она находится поверх щитовидной железы) регулирует содержание кальция в организме. Многие заболевания связаны с нарушением обмена кальция в организме. Например, его недостаток приводит к рахиту у маленьких детей.
На мембране одного нейрона могут одновременно находиться два вида синапсов: тормозные и возбудительные. Все определяется устройством ионного канала мембраны. Мембрана возбудительных синапсов пропускает как ионы натрия, так и ионы калия. В этом случае мембрана нейрона деполяризуется. Мембрана тормозных синапсов пропускает только ионы хлора и гиперполяризуется. Очевидно, что если нейрон заторможен, потенциал мембраны увеличивается (гиперполяризация). Таким образом, нейрон благодаря воздействию через соответствующие синапсы может возбудиться или прекратить возбуждение, затормозиться. Все эти события происходят на соме и многочисленных отростках дендрита нейрона, на последних находится до нескольких тысяч тормозных и возбудительных синапсов.
В качестве примера разберем, как действует в синапсе медиатор, который называется ацетилхолином. Этот медиатор широко распространен в головном мозге и в периферических окончаниях нервных волокон. Например, двигательные импульсы, которые по соответствующим нервам приводят к сокращению мышц нашего тела, оперируют ацетилхолином. Ацетилхолин был открыт в 30-х годах австрийским ученым О. Леви. Эксперимент был очень прост: изолировали сердце лягушки с подходящим к нему блуждающим нервом. Было известно, что электрическая стимуляция блуждающего нерва приводит к замедлению сокращений сердца вплоть до полной его остановки. О. Леви простимулировал блуждающий нерв, получил эффект остановки сердца и взял из сердца немного крови. Оказалось, что если эту кровь добавить в желудочек работающего сердца, то оно замедляет свои сокращения. Был сделан вывод: при стимуляции блуждающего нерва выделяется вещество, останавливающее сердце. Это и был ацетилхолин. Позже был открыт фермент, который расщеплял ацетилхолин на холин (жир) и уксусную кислоту, в результате чего прекращалось действие медиатора. Этим исследованием впервые была установлена точная химическая формула медиатора и последовательность событий в типичном химическом синапсе. Эта последовательность событий сводится к следующему.
Потенциал действия, пришедший по пресинаптическому волокну к синапсу, вызывает деполяризацию, которая включает кальциевый насос, и ионы кальция поступают в синапс; ионы кальция связываются белками мембраны синаптических пузырьков, что приводит к активному опорожнению (экзоцитозу) пузырьков в синаптическую щель. Молекулы медиатора связываются (узнающим центром) соответствующими рецепторами постсинаптической мембраны, при этом открывается ионный канал. Через мембрану начинает протекать ионный ток, что приводит к возникновению на ней постсинаптического потенциала. В зависимости от характера открытых ионных каналов возникает возбудительный (открываются каналы для ионов натрия и калия) или тормозной (открываются каналы для ионов хлора) постсинаптический потенциал.
Ацетилхолин весьма широко распространен в живой природе. Например, он находится в стрекательных капсулах крапивы, в стрекательных клетках кишечнополостных животных (например, пресноводной гидры, медузы) и пр. В нашем организме ацетилхолин выбрасывается в окончаниях двигательных нервов, управляющих мышцами, из окончаний блуждающего нерва, который управляет деятельностью сердца и других внутренних органов. Человек давно знаком с антагонистом ацетилхолина - это яд кураре, которым пользовались индейцы Южной Америки при охоте на животных. Оказалось, что кураре, попадая в кровь, вызывает обездвиживание животного, и оно погибает фактически от удушья, но кураре не останавливает сердце. Исследования показали, что в организме существуют два типа рецепторов к ацетилхолину: один успешно связывает никотиновую кислоту, а другой - мускарин (вещество, которое выделено из гриба рода Muscaris). На мышцах нашего тела находятся рецепторы никотинового типа к ацетилхолину, тогда как на сердечной мышце и нейронах головного мозга - рецепторы к ацетилхолину мускаринового типа.
К биогенным аминам относят также адреналин и норадреналин, которые содержатся в синапсах нейронов автономной нервной вегетативной системы. Во время стресса под влиянием специального гормона - адренокортикотропного (подробнее см. ниже) - из клеток коры надпочечников в кровь также выбрасываются адреналин и норадреналин.
В настоящее время при классификации медиаторных веществ принято выделять: первичные медиаторы, сопутствующие медиаторы, медиаторы-модуляторы и аллостерические медиаторы. Первичными медиаторами считают те, которые действуют непосредственно на рецепторы постсинаптической мембраны. Сопутствующие медиаторы и медиаторы-модуляторы могут запускать каскад ферментативных реакций, которые, например, фосфорилируют рецептор для первичного медиатора. Аллостерические медиаторы могут участвовать в кооперативных процессах взаимодействия с рецепторами первичного медиатора.
Одними из самых распространенных являются медиаторы, принадлежащие к группе биогенных аминов. Эта группа медиаторов достаточно надежно идентифицируется микрогистологическими методами. Известны две группы биогенных аминов: катехоламины (дофамин, норадреналин и адреналин) и индоламин (серотонин). Функции биогенных аминов в организме весьма многообразны: медиаторная, гормональная, регуляция эмбриогенеза.
Основным источником норадренергических аксонов являются нейроны голубого пятна и прилежащих участков среднего мозга (рис. 2.14). Аксоны этих нейронов широко распространяются в мозговом стволе, мозжечке, в больших полушариях. В продолговатом мозге крупное скопление норадренергических нейронов находится в вентролатеральном ядре ретикулярной формации. В промежуточном мозге (гипоталамусе) норадренергические нейроны наряду с дофаминергическими нейронами входят в состав гипоталамо-гипофизарной системы. Норадренергические нейроны в большом количестве содержатся в нервной периферической системе. Их тела лежат в симпатической цепочке и в некоторых интрамуральных ганглиях.
Дофаминергические нейроны у млекопитающих находятся преимущественно в среднем мозге (так называемая нигро-неостриарная система), а также в гипоталамической области. Дофаминовые цепи мозга млекопитающих хорошо изучены. Известны три главные цепи, все они состоят из однонейронной цепочки. Тела нейронов находятся в мозговом стволе и отсылают аксоны в другие области головного мозга (рис. 2.15).
Одна цепь очень проста. Тело нейрона находится в области гипоталамуса и отсылает короткий аксон в гипофиз. Этот путь входит в состав гипоталамо-гипофизарной системы и контролирует систему эндокринных желез.
Вторая дофаминовая система также хорошо изучена. Это черная субстанция, многие клетки которой содержат дофамин. Аксоны этих нейронов проецируются в полосатые тела. Эта система содержит примерно 3/4 дофамина головного мозга. Она имеет решающее значение в регулировании тонических движений. Дефицит дофамина в этой системе приводит к болезни Паркинсона. Известно, что при этом заболевании происходит гибель нейронов черной субстанции. Введение L-DOPA (предшественника дофамина) облегчает у больных некоторые симптомы заболевания.
Третья дофаминергическая система участвует в проявлении шизофрении и некоторых других психических заболеваний. Функции этой системы пока изучены недостаточно, хотя сами пути хорошо известны. Тела нейронов лежат в среднем мозге рядом с черной субстанцией. Они проецируют аксоны в вышележащие структуры мозга, мозговую кору и лимбическую систему, особенно к фронтальной коре, к септальной области и энторинальной коре. Энторинальная кора, в свою очередь, является главным источником проекций к гиппокампу.
Согласно дофаминовой гипотезе шизофрении, третья дофаминергическая система при этом заболевании сверхактивна. Эти представления возникли после открытия веществ, снимающих некоторые симптомы заболевания. Например, хлорпромазин и галоперидол имеют разную химическую природу, но они одинаково подавляют активность дофаминергической системы мозга и проявление некоторые симптомов шизофрении. У больных шизофренией, в течение года получавших эти препараты, появляются двигательные нарушения, получившие название tardive dyskinesia (повторяющиеся причудливые движения лицевой мускулатуры, включая мускулатуру рта, которые больной не может контролировать).
Серотонин почти одновременно открыли в качестве сывороточного сосудосуживающего фактора (1948) и энтерамина, секретируемого энтерохромаффиновыми клетками слизистой оболочки кишечника. В 1951 г. было расшифровано химическое строение серотонина и он получил новое название - 5-гидрокситриптамин. В организме млекопитающих он образуется гидроксилированием аминокислоты триптофана с последующим декарбоксилированием. 90% серотонина образуется в организме энтерохромаффиновыми клетками слизистой оболочки всего пищеварительного тракта. Внутриклеточный серотонин инактивируется моноаминоксидазой, содержащейся в митохондриях. Серотонин внеклеточного пространства окисляется перулоплазмином. Большая часть вырабатываемого серотонина связывается с кровяными пластинками и по кровяному руслу разносится по организму. Другая часть действует в качестве местного гормона, способствуя авторегулированию кишечной перистальтики, а также модулируя эпителиальную секрецию и всасывание в кишечном тракте.
Серотонинергические нейроны широко распространены в центральной нервной системе (рис. 2.16). Они обнаруживаются в составе дорсального и медиального ядер шва продолговатого мозга, а также в среднем мозге и варолиевом мосту. Серотонинергические нейроны иннервируют обширные области мозга, включающие кору больших полушарий, гиппокамп, бледный шар, миндалину, область гипоталамуса. Интерес к серотонину был привлечен в связи с проблемой сна. При разрушении ядер шва животные страдали бессонницей. Сходный эффект оказывали вещества, истощающие хранилище серотонина в мозге.
Самая высокая концентрация серотонина обнаружена в эпифизе (pineal gland). Серотонин в эпифизе превращается в мелатонин, который участвует в пигментации кожи, а также влияет у многих животных на активность женских гонад. Содержание как серотонина, так и мелатонина в эпифизе контролируется циклом свет - темнота через нервную симпатическую систему.
Другую группу медиаторов ЦНС составляют аминокислоты. Уже давно известно, что нервная ткань с ее высоким уровнем метаболизма содержит значительные концентрации целого набора аминокислот (перечислены в порядке убывания): глутаминовой кислоты, глутамина, аспарагиновой кислоты, гамма-аминомасляной кислоты (ГАМК).
Глутамат в нервной ткани образуется преимущественно из глюкозы. У млекопитающих больше всего глутамата содержится в конечном мозге и мозжечке, где его концентрация примерно в 2 раза выше, чем в стволе мозга и спинном мозге. В спинном мозге глутамат распределен неравномерно: в задних рогах он находится в большей концентрации, чем в передних. Глутамат является одним из самых распространенных медиаторов в ЦНС.
Постсинаптические рецепторы к глутамату классифицируются в соответствии с аффинностью (сродством) к трем экзогенным агонистам - квисгулату, каинату и N-метил-D-аспартату (NMDA). Ионные каналы, активируемые квисгулатом и каинатом, подобны каналам, которые управляются никотиновыми рецепторами - они пропускают смесь катионов (Na + и. К + ). Стимуляция NMDA-рецепторов имеет сложный характер активации: ионный ток, который переносится не только Na + и К + , но также Са ++ при открывании ионного канала рецептора, зависит от потенциала мембраны. Вольтзависимая природа этого канала определяется разной степенью его блокирования ионами Mg ++ с учетом уровня мембранного потенциала. При потенциале покоя порядка - 75 мВ ионы Mg ++ , которые преимущественно находятся в межклеточной среде, конкурируют с ионами Са ++ и Na + за соответствующие каналы мембраны (рис. 2.17). Вследствие того, что ион Mg ++ не может пройти через пору, канал блокируется всякий раз, как попадает туда ион Mg ++ . Это приводит к уменьшению времени открытого канала и проводимости мембраны. Если мембрану нейрона деполяризовать, то количество ионов Mg ++ , которые закрывают ионный канал, снижается и через канал беспрепятственно могут проходить ионы Са ++ , Na + и. К + . При редких стимуляциях (потенциал покоя изменяется мало) глутаматергического рецептораВПСП возникает преимущественно за счет активации квисгулатных и каинатных рецепторов; вклад NMDA-рецепторов незначителен. При длительной деполяризации мембраны (ритмическая стимуляция) магниевый блок удаляется, и NMDA-каналы начинают проводить ионы Са ++ , Na + и. К + . Ионы Са ++ через вторичные посредники могут потенцировать (усиливать) минПСП, что может привести, например, к длительному увеличению синаптической проводимости, сохраняющейся часами и даже сутками.
Из тормозных медиаторов ГАМК является самой распространенной в ЦНС. Она синтезируется из L-глутаминовой кислоты в одну стадию ферментом декарбоксилазой, наличие которой является лимитирующим фактором этого медиатора. Известно два типа ГАМК-рецепторов на постсинаптической мембране: ГАМКА (открывает каналы для ионов хлора) и ГАМКБ (открывает в зависимости от типа клетки каналы для. К + или Са ++ ). На рис. 2.18 показана схема ГАМК-рецептора. Интересно, что в его состав входит бензодиазипиновый рецептор, наличием которого объясняют действие так называемых малых (дневных) транквилизаторов (седуксена, тазепама и др.). Прекращение действия медиатора в ГАМК-синапсах происходит по принципу обратного всасывания (молекулы медиатора специальным механизмом поглощаются из синаптической щели в цитоплазму нейрона). Из антагонистов ГАМК хорошо известен бикукулин. Он хорошо проходит через гематоэнцефалический барьер, оказывает сильное воздействие на организм даже в малых дозах, вызывая конвульсии и смерть. ГАМК обнаруживается в ряде нейронов мозжечка (в клетках Пуркинье, клетках Гольджи, корзинчатых клетках), гиппокампа (в корзинчатых клетках), в обонятельной луковице и черной субстанции.
Идентификация ГАМК-цепей мозга трудна, так как ГАМК - обычный участник метаболизма в ряде тканей организма. Метаболическая ГАМК не используется как медиатор, хотя в химическом отношении их молекулы одинаковы. ГАМК определяется по ферменту декарбоксилазы. Метод основан на получении у животных антител к декарбоксилазе (антитела экстрагируют, метят и вводят в мозг, где они связываются с декарбоксилазой).
Другим известным тормозным медиатором является глицин. Глицинергические нейроны находятся главным образом в спинном и продолговатом мозге. Считают, что эти клетки выполняют роль тормозных интернейронов.
Ацетилхолин - один из первых изученных медиаторов. Он чрезвычайно широко распространен в нервной периферической системе. Примером могут служить мотонейроны спинного мозга и нейроны ядер черепных нервов. Как правило, холинергические цепи в мозге определяют по присутствию фермента холинэстеразы. В головном мозге тела холинергических нейронов находятся в ядре перегородки, ядре диагонального пучка (Брока) и базальных ядрах. Нейроанатомы считают, что эти группы нейронов формируют фактически одну популяцию холинергических нейронов: ядро педнего мозга, nucleus basalis (оно расположено в базальной части переднего мозга) (рис. 2.19). Аксоны соответствующих нейронов проецируются к структурам переднего мозга, особенно в новую кору и гиппокамп. Здесь встречаются оба типа ацетилхолиновых рецепторов (мускариновые и никотиновые), хотя считается, что мускариновые рецепторы доминируют в более рострально распоженных мозговых структурах. По данным последних лет складывается впечатление, что ацетилхолиновая система играет большую роль в процессах, связанных с высшими интегративными функциями, которые требуют участия памяти. Например, показано, что в мозге больных, умерших от болезни Альцгеймера, наблюдается массивная утрата холинергических нейронов в nucleus basalis.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также:
