
Обработка информации в цнс
Миелиновые аксоны ганглиозных клеток сетчатки направляются в головной мозг в составе двух зрительных нервов (рис.23.7). Правый и левый зрительные нервы сливаются у основания черепа, образуя зрительный перекрест (хиазму). Здесь нервные волокна, идущие от медиальной половины сетчатки каждого глаза переходят на контрлатеральную сторону, а волокна от латеральных половин сетчаток продолжаются ипсилатерально.
После перекреста аксоны ганглиозных клеток в составе зрительного тракта следуют в латеральные коленчатые тела (ЛКТ), где образуют синаптические контакты с нейронами ЦНС. Аксоны нервных клеток ЛКТ в составе т.н. зрительной лучистости достигают нейронов первичной зрительной коры (поле 17 по Бродману). Далее по внутрикорковым связям возбуждение распространяется во вторичную зрительную кору (поля 18б 19) и ассоциативные зоны коры.
Сенсорные пути зрительной системы организованы по ретинотопическому принципу - возбуждение от соседних ганглиозных клеток достигает соседних точек ЛКТ и коры. Поверхность сетчатки как бы проецируется на поверхность ЛКТ и коры.
Большая часть аксонов ганглиозных клеток заканчиваются в ЛКТ, часть же волокон следует в верхние бугры двухолмия, гипоталамус, претектальную область ствола мозга, ядро зрительного тракта. Связь между сетчаткой и верхними буграми четверохолмия служит для регуляции движений глаз. Проекция сетчатки в гипоталамус служит для сопряжения эндогенных циркадных ритмов с суточными колебаниями уровня освещенности.
Связь между сетчаткой и претектальной областью ствола исключительно важна для регуляции просвета зрачка и аккомодации. Нейроны ядер зрительного тракта, которые также получают синаптические входы от ганглиозных клеток, связаны с вестибулярными ядрами ствола мозга. Эта проекция позволяет оценивать положение тела в пространстве на основании зрительных сигналов, а также служит для осуществления сложных глазодвигательных реакций (нистагм).
Обработка зрительной информации в ЛКТ
Нейроны ЛКТ имеют рецептивные поля округлой формы. Электрические реакции этих клеток аналогичны таковым ганглиозных клеток. В ЛКТ существуют нейроны, которые возбуждаются при наличии в их рецептивном поле границы свет/темнота (контрастные нейроны) или при передвижении этой границы в пределах рецептивного поля (детекторы движения). Обработка зрительной информации в первичной зрительной коре В зависимости от реакции на световые стимулы нейроны коры подразделяют на несколько классов. Нейроны с простым рецептивным полем. Наиболее сильное возбуждение такого нейрона происходит при освещении его рецептивного поля световой полоской определенной ориентации. Частота нервных импульсов, генерируемых таким нейроном уменьшается при изменении ориентации световой полоски.
Нейроны со сложным рецептивным полем
Максимальная степень возбуждения нейрона достигается при передвижении светового стимула в пределах ON зоны рецептивного поля в определенном направлении. Передвижение светового стимула в другом направлении или выход светового стимула за пределы ON зоны вызывает более слабое возбуждение
Нейроны со сверхсложным рецептивным полем
Максимальное возбуждение такого нейрона достигается при действии светового стимула сложной конфигурации. Например, известны нейроны, наиболее сильное возбуждение которых развивается при пересечении двух границ между светлым и темным в пределах ON зоны рецептивного поля.
Несмотря на огромно количество экспериментальных данных о закономерностях реагирования клеток на разные зрительные стимулы, к настоящему времени нет полной теории, объясняющей механизмы обработки зрительной информации в головном мозге. Мы не можем объяснить, каким образом разнообразные электрические реакции нейронов сетчатки, ЛКТ и коры обеспечивают распознавание образов и другие феномены зрительного восприятия.
Регуляция движений глаз
Часть волокон ганглиозных клеток следуют к нейронам верхних бугров четверохолмия (средний мозг), которые связаны с ядрами глазодвигательного, блокового и отводящего нервов, нейроны которых иннервируют поперечнополосатые мышечные волокна мышц глаза. Нервные клетки верхних бугров получат синаптические входы от вестибюлярных рецепторов, проприорецепторов мышц шеи, что позволяет организму координировать движения глаз с перемещениями тела в пространстве.
Зрительная система обладает замечательной способность распознавать объект при самых разных вариантах его изображения. Мы можем узнавать образ (знакомое лицо, букву и т. п.), когда некоторых его частей недостает, когда он содержит лишние элементы, когда он по-разному ориентирован в пространстве, имеет разные угловые размеры, повернут к нам разными сторонами и т.п.. Постоянство формы и размеров. Как правило, мы воспринимаем окружающие предметы неизменными по форме и размерам. Хотя на самом деле их форма и размеры на сетчатке не являются постоянными. Например, велосипедист в поле зрения всегда кажется одинаковым по величине независимо от расстояния до него. Колеса велосипеда воспринимаются как круглые, хотя на самом деле их изображения на сетчатке могут быть узкими эллипсами. Это явление демонстрирует роль опыта в видении окружающего мира. Нейрофизиологические механизмы этого феномена в настоящее время неизвестны.
Восприятие глубины пространства
Изображение окружающего мира на сетчатке является плоским. Однако, мы видим мир объемным. Существует несколько механизмов, которые обеспечивают построение 3-мерного пространства на основании плоских изображений, сформированных на сетчатке.
Поскольку глаза расположены на некотором расстоянии друг от друга, то изображения, формирующиеся на сетчатке левого и правого глаза, несколько различаются друг от друга. Чем ближе расположен объект по отношению к наблюдателю, тем больше будут различаться эти изображения. Чем дальше расположен объект, тем меньше угловой размер его изображения на сетчатке. Оценивая угловые размеры знакомых предметов, мы можем оценивать расстояние до них. Перекрывание изображений также помогает оценить их взаимное расположение в пространстве. Изображение близкого предмета может перекрывать изображение удаленного, но не наоборот. При смещении головы наблюдателя изображения наблюдаемых объектов на сетчатке также будут смещаться (явление параллакса). При одном и том же смещении головы изображения близких объектов будут смещаться сильнее, чем изображения удаленных
Восприятие неподвижности пространства
Если, закрыв один глаз, нажать пальцем на второе глазное яблоко, то мы увидим, что мир вокруг нас смещается в сторону. В обычных условиях окружающий мир неподвижен, хотя изображение на сетчатке постоянно "прыгает" за счет перемещения глазных яблок, поворотов головы, изменения положения тела в пространстве. Восприятие неподвижности окружающего пространства обеспечивается тем, что при обработке зрительных образов учитывается информация о движении глаз, движениях головы и положении тела в пространстве. Зрительная сенсорная система умеет "вычитать" собственные движения глаз и тела из перемещения изображения на сетчатке. Рецептивное поле область, занимаемая совокупностью всех рецепторов, стимуляция которых приводит к изменению активности определенного элемента: афферентного волокна (Р. п. нерва) или сенсорного нейрона (Р. п. нейрона). Последнее оказывается более сложным, особенно для центральных нейронов, поскольку в зависимости от конкретных характеристик стимула Р. п. может оказаться различным. Понятие Р. п. используется и для обозначения зоны расположения чувствительных элементов, стимуляция которых приводит к возникновению специализированного рефлекса, Р. п. рефлекса, или рефлексогенная зона (например, интероцептивные рефлексы сердечно-сосудистой системы развиваются как следствие активации синокаротидной зоны, барорецепторной зоны дуги аорты и т.д.).
Проводниковый отдел начинается в сетчатке (первый нейрон – биполярный, второй нейрон – ганглиозные клетки). Далее - зрительные нервы, частичный перекрест в хиазме и зрительные тракты. В каждом зрительном тракте содержатся нервные волокна, идущие от внутренней (носовой) поверхности сетчатки глаза одноименной стороны и от наружной половины сетчатки другого глаза. Зрительный тракт заканчивается в подкорковых центрах зрения: латеральном коленчатом теле (главный подкорковый центр), подушке таламуса и верхних холмиках среднего мозга (третий нейрон).
От них зрительные нервные волокна направляются в кору полушарий большого мозга. 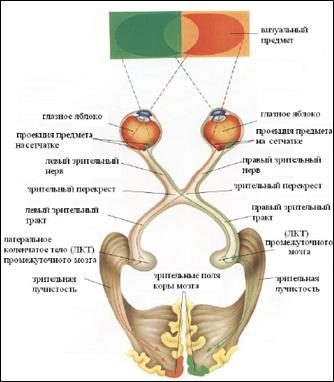
Аксоны третьего нейрона латерального коленчатого тела (в основном) и таламуса образуют зрительную лучистость, которая проходит через заднюю часть задней ножки внутренней капсулы и заканчивается в коре шпорной борозды.
Волокна от ядер верхних холмиков соединяются с парасимпатическим ядром III пары черепных нервов. От этого ядра импульс передается через ресничный узел к сфинктеру зрачка и вызывает его сужение. Другая часть волокон от верхних холмиков направляется в спинной мозг к клеткам передних рогов, обеспечивая автоматические рефлекторные движения в ответ на зрительные раздражения.
В наружных (или латеральных) коленчатых телах, куда приходят волокна из сетчатки, есть рецептивные поля, которые также имеют округлую форму, но меньше по размеру, чем в сетчатке. Ответы нейронов здесь носят фазический характер, но более выражены, чем в сетчатке. На уровне наружных коленчатых тел происходит процесс взаимодействия афферентных сигналов, идущих от сетчатки глаза, с эфферентными из области коркового отдела зрительного анализатора. С участием ретикулярной формации здесь происходит взаимодействие со слуховой и другими сенсорными системами, что обеспечивает процессы избирательного зрительного внимания путем выделения наиболее существенных компонентов сенсорного сигнала.
Центральный,или корковый, отделзрительного анализатора расположен в затылочной доле (поля 17, 18, 19).
Поле 17 осуществляет специализированную, но более сложную, чем в сетчатке и в наружных коленчатых телах, переработку информации. РП нейронов зрительной коры небольших размеров имеют вытянутые, почти прямоугольные, а не округлые формы. Наряду с этим имеются сложные и сверхсложные рецептивные поля детекторного типа. Эта особенность позволяет выделять из цельного изображения лишь отдельные части линий с различным расположением и ориентацией, при этом проявляется способность избирательно реагировать на эти фрагменты.
В каждом участке коры сконцентрированы нейроны, которые образуют колонку, проходящую по глубине через все слои вертикально, при этом происходит функциональное объединение нейронов, выполняющих сходную функцию.
Разные свойства зрительных объектов (цвет, форма, движение) обрабатываются в разных частях зрительной коры параллельно.
В зрительной коре существуют функционально различные группы клеток – простые и сложные.
Простые клеткисоздают РП, которое состоит из возбудительной и тормозной зон. Сложные клеткиявляются детекторами угла, наклона и движения линий в поле зрения.В одной колонке могут располагаться как простые, так и сложные клетки. Сложные клетки расположены в более поверхностных слоях поля 17, в полях 18 и 19 простые клетки являются исключением, там расположены сложные и сверхсложные клетки.
В зрительной коре часть нейронов образует цветооппонентные рецептивные поля ( одни нейроны реагируют on-ответом на красное освещение и off-ответом на зеленое, реакция других – обратная).
Т. о., на основании полученных данных полагают, что процессы в колбочках более соответствуют трехкомпонентной теории цветоощущения, тогда как для нейронных сетей сетчатки и вышележащих зрительных центров подходит теория контрастных цветов Геринга.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Обработка информации в ЦНС. Рецептивные поля
Миелиновые аксоны ганглиозных клеток сетчатки направляются в головной мозг в составе двух зрительных нервов (рис.23.7). Правый и левый зрительные нервы сливаются у основания черепа, образуя зрительный перекрест (хиазму). Здесь нервные волокна, идущие от медиальной половины сетчатки каждого глаза переходят на контрлатеральную сторону, а волокна от латеральных половин сетчаток продолжаются ипсилатерально.
После перекреста аксоны ганглиозных клеток в составе зрительного тракта следуют в латеральные коленчатые тела (ЛКТ), где образуют синаптические контакты с нейронами ЦНС. Аксоны нервных клеток ЛКТ в составе т.н. зрительной лучистости достигают нейронов первичной зрительной коры (поле 17 по Бродману). Далее по внутрикорковым связям возбуждение распространяется во вторичную зрительную кору (поля 18б 19) и ассоциативные зоны коры.
Сенсорные пути зрительной системы организованы по ретинотопическому принципу - возбуждение от соседних ганглиозных клеток достигает соседних точек ЛКТ и коры. Поверхность сетчатки как бы проецируется на поверхность ЛКТ и коры.
Большая часть аксонов ганглиозных клеток заканчиваются в ЛКТ, часть же волокон следует в верхние бугры двухолмия, гипоталамус, претектальную область ствола мозга, ядро зрительного тракта. Связь между сетчаткой и верхними буграми четверохолмия служит для регуляции движений глаз. Проекция сетчатки в гипоталамус служит для сопряжения эндогенных циркадных ритмов с суточными колебаниями уровня освещенности.
Связь между сетчаткой и претектальной областью ствола исключительно важна для регуляции просвета зрачка и аккомодации. Нейроны ядер зрительного тракта, которые также получают синаптические входы от ганглиозных клеток, связаны с вестибулярными ядрами ствола мозга. Эта проекция позволяет оценивать положение тела в пространстве на основании зрительных сигналов, а также служит для осуществления сложных глазодвигательных реакций (нистагм).
Обработка зрительной информации в ЛКТ
Нейроны ЛКТ имеют рецептивные поля округлой формы. Электрические реакции этих клеток аналогичны таковым ганглиозных клеток. В ЛКТ существуют нейроны, которые возбуждаются при наличии в их рецептивном поле границы свет/темнота (контрастные нейроны) или при передвижении этой границы в пределах рецептивного поля (детекторы движения). Обработка зрительной информации в первичной зрительной коре В зависимости от реакции на световые стимулы нейроны коры подразделяют на несколько классов. Нейроны с простым рецептивным полем. Наиболее сильное возбуждение такого нейрона происходит при освещении его рецептивного поля световой полоской определенной ориентации. Частота нервных импульсов, генерируемых таким нейроном уменьшается при изменении ориентации световой полоски.
Нейроны со сложным рецептивным полем
Максимальная степень возбуждения нейрона достигается при передвижении светового стимула в пределах ON зоны рецептивного поля в определенном направлении. Передвижение светового стимула в другом направлении или выход светового стимула за пределы ON зоны вызывает более слабое возбуждение
Нейроны со сверхсложным рецептивным полем
Максимальное возбуждение такого нейрона достигается при действии светового стимула сложной конфигурации. Например, известны нейроны, наиболее сильное возбуждение которых развивается при пересечении двух границ между светлым и темным в пределах ON зоны рецептивного поля.
Несмотря на огромно количество экспериментальных данных о закономерностях реагирования клеток на разные зрительные стимулы, к настоящему времени нет полной теории, объясняющей механизмы обработки зрительной информации в головном мозге. Мы не можем объяснить, каким образом разнообразные электрические реакции нейронов сетчатки, ЛКТ и коры обеспечивают распознавание образов и другие феномены зрительного восприятия.
Регуляция движений глаз
Часть волокон ганглиозных клеток следуют к нейронам верхних бугров четверохолмия (средний мозг), которые связаны с ядрами глазодвигательного, блокового и отводящего нервов, нейроны которых иннервируют поперечнополосатые мышечные волокна мышц глаза. Нервные клетки верхних бугров получат синаптические входы от вестибюлярных рецепторов, проприорецепторов мышц шеи, что позволяет организму координировать движения глаз с перемещениями тела в пространстве.
Зрительная система обладает замечательной способность распознавать объект при самых разных вариантах его изображения. Мы можем узнавать образ (знакомое лицо, букву и т. п.), когда некоторых его частей недостает, когда он содержит лишние элементы, когда он по-разному ориентирован в пространстве, имеет разные угловые размеры, повернут к нам разными сторонами и т.п Постоянство формы и размеров. Как правило, мы воспринимаем окружающие предметы неизменными по форме и размерам. Хотя на самом деле их форма и размеры на сетчатке не являются постоянными. Например, велосипедист в поле зрения всегда кажется одинаковым по величине независимо от расстояния до него. Колеса велосипеда воспринимаются как круглые, хотя на самом деле их изображения на сетчатке могут быть узкими эллипсами. Это явление демонстрирует роль опыта в видении окружающего мира. Нейрофизиологические механизмы этого феномена в настоящее время неизвестны.
Виды тестов. Тесты-опросники
Опросники - обширная группа психодиагностических методик, задания в которых представлены в виде вопросов или утверждений. Эти вопросы или утверждения не имеют правильных вариантов ответа и характеризуются только их частотой и направленностью. Эта группа методик предоставлена для получения объективных и субъективных данных об испытуемых. .
Различные концептуальные подходы к освещению проблемы.
Личностные особенности подросткового возраста изучали многие зарубежные и русские психологи. На данную проблему существует множество точек зрения, рассмотрим лишь некоторые: Е. Доуан Для среднего подростка совершенно не характерны ни Дж. Адельсон личностные кризисы, ни крах “я” - концепции, ни тенденции от ранее приобретенных ценнос .
Защитные психические механизмы
Что такое "защитные механизмы"? В связи с тем, что как уже описывалось, согласно 3.Фрейду, в человеческой психике происходит постоянный конфликт между уровнями личности, для смягчения вызываемой этим конфликтом чувства напряженности и вины человеческая психика выработала ряд защитных механизмов, призванных бессознательно подав .
- На головну
- Контакти
- 0
- 1
- 2
- 3
- 4
- 5
Основные понятия и ключевые термины: ПОЛУШАРИЯ БОЛЬШОГО МОЗГА. КОРА ПОЛУШАРИЙ.
Вспомните! Что такое большой мозг?
Издавна считают, что формирование мозга человека завершается в раннем детстве, но, как оказалось, на самом деле это не так. Проведя целый ряд экспериментов, учёные Великобритании установили, что лобные кости изменяются до 40 лет, что позволяет мозгу человека постоянно расти и развиваться, особенно лобовым частям. А изменяется ли количество извилин полушарий большого мозга у человека в течение жизни?

Каковы особенности строения и функций полушарий большого мозга?
ПОЛУШАРИЯ БОЛЬШОГО МОЗГА
мозга, которые имеют выраженную складчатую поверхность, обеспечивают обработку информации и формирование сложных форм поведения человека. У каждого человека поверхность полушарий разная. Неповторимость этой поверхности обусловлена большим количеством малых и больших борозд и извилин. Складки полушарий начинают формироваться ещё в зародышевом развитии. У новорождённых уже есть первичные, вторичные и третичные борозды, но они продолжают развиваться и после рождения, особенно до 1-2 лет. К 7-12 годам борозды и извилины уже имеют такой вид, как у взрослого человека.
На поверхности полушарий есть три крупнейшие борозды: центральная (1), боковая (2) и затылочно-теменная (7). Они выделяют в каждом полушарии 4 основных части: лобную (4), теменную (5), затылочную (6) и височную (3) (ил. 85).

Отличительным признаком полушарий большого мозга человека является наличие хорошо развитой коры, которая образована серым веществом и покрывает всю их поверхность. Но основную массу конечного мозга составляет белое вещество, расположенная под корой.
Ещё одной важной характеристикой полушарий большого мозга человека является распределение функций между ними, или функциональная специализация. Левое полушарие отвечает за такие важные функции, как речь, чтение, письмо, счёт, анализ, интеллект. Правое полушарие отвечает
за образное восприятие, воображение, чувства, синтез и интуицию. В каждое полушарие нервные импульсы поступают с противоположной стороны тела и пространства: правое полушарие управляет органами левой части тела и получает информацию слева, а левая - наоборот. Для успешной работы нужна слаженная работа обоих полушарий мозга. У каждого человека одно из полушарий является председательствующим (доминантным): у правшей - левое полушарие, у левшей - правое.
Итак, особенности внешнего строения полушарий большого мозга человека связаны с сильно выраженной складчатой поверхностью, которая имеет индивидуальный характер, закладывается ещё до рождения и развивается к периоду половой зрелости.
Каково строение коры полушарий большого мозга?
КОРА БОЛЬШИХ ПОЛУШАРИЙ - слой серого вещества на поверхности полушарий большого мозга, которая является центром обработки информации. Она составляет примерно 44 % от объёма полушарий. Толщина коры - 3-5 мм, а её общая площадь за счёт борозд и извилин составляет 2000 - 2500 см 2 . Количество нейронов коры может изменяться в пределах 10 9 - 10 10 , и каждый имеет от 7 до 10 тыс. связей с соседними клетками, определяет гибкость, устойчивость и надежность функций коры. Кора больших полушарий, несмотря на незначительную толщину, имеет сложное строение. Нейроны коры связаны между собой и с ниже расположенными клетками многочисленными синапсами, образуя шесть горизонтальных слоёв и вертикальные нейрональные колонки. Именно такая колонка и является основной структурно-функциональной единицей коры полушарий человека, осуществляет обработку информации. Нейрональная колонка - это совокупность вертикально расположенных нейронов в коре головного мозга, которая проходит через её слои. Малые колонки содержат от 80 до 120 нейронов почти во всех зонах мозга. Всего в коре человека насчитывается около 2х10 8 таких колонок. Колонки соединяются между собой горизонтальными связями и образуют нейронные ансамбли.
Пространство между телами и отростками нервных клеток коры заполнено нейроглией и капиллярами. Нейроны коры делятся на 2 основных типа: пирамидальные (около 80 % всех клеток коры) и звёздчатые (около 20 %). Большие пирамидальные клетки (клетки Беца) открыл в 1874 году украинский анатом В. Бец (1834-1894).
Серое вещество коры полушарий имеет свое продолжение в нижнем белом веществе. Скопление серого вещества в белом, что является эволюционно старой частью полушарий большого мозга, называется подкоркой. К подкорковым структурам относят ядра промежуточного мозга, среднего мозга, ретикулярную формацию, лимбическую систему и др.


Итак, кора полушарий большого мозга состоит из серого вещества и имеет сложную трёхмерную организацию нейронов, функционирующих как единое целое благодаря многочисленным связям.
Какую роль выполняет кора в обработке информации?
В коре большого мозга различают три вида участков - чувствительные, двигательные и ассоциативные зоны. Чувствительные поля получают информацию от различных рецепторов и являются корковыми участками её анализа, двигательные - посылают командные импульсы к рабочим органам, ассоциативные - соединяют между собой двигательные и чувствительные зоны, анализируют и хранят информацию, именно с ними связаны обучение, речь, мышление, сознание. Основными полями, которые отличаются строением и функциями, являются кожно-мышечная, двигательная, слуховая, обонятельная, вкусовая, зрительная зоны.
Кора лобной доли играет определяющую роль в обучении, регуляции поведения в нестандартных ситуациях, использовании накопленного опыта человека, выявлении чувств. В лобной доле левого полушария расположен двигательный центр речи, обеспечивающий способность человека писать предложения и произносить слова. Спереди от центральной борозды расположена двигательная зона коры, которая обеспечивает движения мышц и суставов тела.
В коре теменной доли позади центральной борозды расположена зона кожно-мышечной чувствительности, к которой поступает информация от кожи, суставов и мышц и с которой связаны ощущения прикосновения, боли и температуры. Но большая часть этой доли, как и лобной, занята ассоциативными полями, которые в значительной мере определяют поведение человека.
В коре затылочной доли расположены зрительные зоны, обеспечивающие чёткое восприятие зрительных объектов, их запоминание и распознавание символических зрительных образов (букв, цифр, мимики и т. д.).
В коре височной доли расположены обонятельные, слуховые и вкусовые зоны.

Итак, кора является самым высоким центром обработки информации; обеспечивает осуществление рефлексов, составляющих основу ВНД (память, эмоции, мышление и т. д.).
Самостоятельная работа с иллюстрацией
Примените знание английского языка и определите названия, расположение и значение чувствительных, двигательных и ассоциативных зон коры в долях полушарий большого мозга.


Blue Brain Project - проект по компьютерному моделированию коры мозга человека, начатый в 2005 году. Над проектом совместно работают компания IBM и Швейцарский технический институт. Проект использует супермощный компьютер Blue Gene для моделирования нейронных сетей. В конце 2006 года удалось смоделировать одну нервную колонку коры молодой крысы. При этом было задействовано 8 192 процессора для моделирования 10 000 нейронов и около 3 х 10 7 синапсов. С 2009 года в США начал действовать ещё один проект Human Connectome Project, целью которого является описание нейрональных путей человека. Какое значение имеют эти проекты для человека?

Владимир Алексеевич Бец (1834-1894) - украинский анатом и гистолог, профессор кафедры анатомии Киевского университета св. Владимира. Большое значение в изучении головного мозга имеют совершённые им научные исследования микроскопического строения коры. Он первым описал шестислойную структуру коры головного мозга (1874) и открыл в ней пирамидальные клетки (клетки Беца). Чем пирамидальные клетки отличаются от других нейронов?

Вопросы для самоконтроля
1. Что такое полушария большого мозга? 2. Назовите доли больших полушарий. 3. Что такое кора полушарий большого мозга? 4. Что является структурно-функциональной единицей коры полушарий? 5. Какие три вида функциональных участков выделяют в коре полушарий? 6. Приведите примеры чувствительных, ассоциативных и двигательных зон коры полушарий.
7. Назовите особенности строения и функций полушарий большого мозга.
8. Опишите строение коры полушарий большого мозга. 9. Какую роль выполняет кора в обработке информации?
10. Как строение коры полушарий взаимосвязана с её функциями?
Идею использования простых физиологических показателей для оценки индивидуальных различий по интеллекту предложил выдающийся биолог Ф. Гальтон (цит. по: Ждан, 2004). Он рассматривал интеллект как биологическое образование, которое можно количественно оценить с помощью физиологических индикаторов. Экспериментальное воплощение эти идеи нашли в ряде работ, где в качестве коррелята интеллекта и частично способа его измерения предлагалось рассматривать время выполнения простых заданий. В современной психологии этот подход четко сформулирован П. Верноном: некоторая часть индивидуальных различий в успешности выполнения тестов интеллекта объясняется различиями в том, насколько быстро индивид может обрабатывать информацию; последнее сравнительно независимо от любых приобретенных знаний или навыков. Таким образом, понятие психической скорости или скорости выполнения умственных действий
Гипотезы о связи умственных способностей и переработки информации в ЦНС. В качестве объекта исследования для установления связей с психометрическим интеллектом могут выступать разные показатели этой скорости, характеризующие отдельные этапы или операции деятельности человека. Это может быть время простой реакции, время реакции выбора и время опознания (inspectin time). По некоторым представлениям скорость проведения нервного импульса должна коррелировать с показателями психометрического интеллекта. Считается, что скорость проведения нервного импульса отражает эффективность функционирования нервной системы: чем выше скорость, тем быстрее нервная система обрабатывает информацию. Ускоренная обработка информации в нервной системе рассматривается как физиологическое условие увеличения умственных возможностей индивида (Рийсдик, Бумсма, 2001).
Согласно другой теории, общие способности могут быть связаны с точностью и безошибочностью передачи информации от нейрона к нейрону (Eysenck, 1982). Иначе говоря, предполагается, что индивиды, нейроны которых быстро проводят информацию вдоль аксона, во-первых, передают информацию эффективно и точно через синапсы, во-вторых, будут более успешно обрабатывать информацию по сравнению с теми, у кого скорость распространения нервных импульсов меньше и при передаче информации происходит больше искажений.
В этом контексте была сформулирована так называемая миели-новая гипотеза (Miller, 1994), согласно которой более высокий интеллект связан с большими размерами мозга, которые, в свою очередь, зависят от толщины миелиновой оболочки. Толстая миелино-вая оболочка, во-первых, обеспечивает более высокую скорость проведения импульса, во-вторых, препятствует вмешательству в этот процесс посторонних сигналов. Тем самым она повышает надежность передачи информации. Подтверждают эту гипотезу положительные корреляции между размером мозга и показателями интеллекта, которые были получены при применении метода ядерно-маг-нитного резонанса.
Время как фактор эффективности. Как уже отмечалось, скорость выполнения умственных действий приобретает роль фактора, объясняющего происхождение индивидуальных различий в познавательной деятельности и показателях интеллекта. Показано, что показатель интеллекта связан с временем реакции, взятом в разных вариантах оценки, отрицательной корреляцией, составляющей в среднем -0,3 (Deary et al., 2010). Помимо этого, хронометрию процессов переработки информации можно провести, используя латентности компонентов ВП, интерпретируемые как маркеры времени выполнения отдельных когнитивных операций. Закономерно, что существует целый ряд исследований взаимосвязи показателей ВП и интеллекта.
Для сопоставления с показателями IQ привлекаются и другие параметры ВП: различные варианты амплитудных оценок, вариативность, асимметрия. Наибольшую известность в связи с этим приобрели исследования взаимосвязи некоторых особенностей ВП, синаптической передачи и формирования энграмм памяти (Hendrickson, 1982; пит. по: Айзенк, 1995). Предполагалось, что при обработке информации на уровне синапсов в коре мозга могут возникать ошибки. Чем большее число таких ошибок продуцирует индивид, тем ниже показатели его интеллекта. Количественно оценить число этих ошибок невозможно, но они проявляются в индивидуальных особенностях конфигурации ВП. Согласно этой концепции индивиды, безошибочно обрабатывающие информацию, должны продуцировать высокоамплитудные и имеющие сложную форму ВП, т.е. с дополнительными пиками и колебаниями (рис. 17.1).
ВП упрощенной формы характерны для индивидов с низкими показателями интеллекта. Эти предположения получили статистическое подтверждение при сопоставлении ВП и показателей интеллекта по тестам Векслера и Равена. Таким образом, есть основания утверждать, что эффективность передачи информации на нейронном уровне определяется двумя параметрами: скоростью и точностью (безошибочностью). Оба параметра можно рассматривать как характеристики биологического интеллекта.
Не только вызванные потенциалы, но и индивидуальные особенности ЭЭГ дают основания для прогноза интеллекта (Разумникова, 2009). Это также подтверждают работы А.Н. Лебедева с соавторами (1997). С использованием статистических средств обработки данных ими было показано, что интеллектуальные способности индивида можно прогнозировать по таким показателям ЭЭГ, как частота альфа-ритма, степень синхронизации колебаний в лобных областях, соотношение мощности колебаний в альфа- и тета-диапазонах.


Рис. 17.1. Вызванные потенциалы у шести испытуемых с высокими (слева) и шести испытуемых с низкими (справа) показателями IQ
(по Eysenck, 1995)
что он ограничивается меньшим числом локусов активации по сравнению с менее успешными испытуемыми. Не исключено, что причина возникшего противоречия в степени сложности заданий. По мере усложнения заданий число локусов активации у индивидуумов с более высоким IQ может увеличиваться (Deary et al., 2010).
Топографические факторы. Роль топографических факторов в обеспечении мышления и интеллекта можно рассматривать по крайней мере в двух аспектах. Первый зависит от морфологических и функциональных особенностей отдельных структур мозга, которые связаны с высокими умственными достижениями. Второй касается особенностей взаимодействия между структурами мозга, при которых возможна высокоэффективная умственная деятельность.
Мышление и интеллект представляют собой свойство мозга как целого, поэтому особое значение приобретает анализ взаимодействия различных регионов мозга, при котором достигается высокоэффективная умственная деятельность. В этом контексте представляет интерес модель теменно-лобной интеграции (P-FIT model), объясняющая корковый механизм решения проблемы. Р. Юнг и Р. Хайер (Jung, Haier, 2007) провели метаанализ 37 эмпирических исследований, выполненных разными вариантами нейровизуализации. Теоретически они исходили из того, что человек собирает и обрабатывает информацию, объединяя данные зрения и слуха. Поэтому особенно важна ранняя сенсорная обработка в экстра-стриарной коре (поля 18 и 19) и веретеновидной извилине (поле 37), отвечающих за формирование визуального опыта, а также зона Вернике (поле 22), отвечающая за смысловую интерпретацию акустических стимулов. Обработанная сенсорная информация направляется в теменную кору, а точнее — в супрамаргинальную извилину (поле 40), верхнюю теменную (поле 7), ангулярную извилину (поле 39) (рис. 17.2). В этих центрах осуществляется смысловая обработка информации, включающая символизацию, абстракцию и уточнение проблемы. Далее теменная кора взаимодействует с лобными областями (поля 6, 9, 10, 45—47). Эти зоны обеспечивают выбор подходящего решения проблемы. Если решение найдено, оно поступает в переднюю поясную кору (поле 32), которая ограничивает возможность дальнейшего выбора и тормозит конкурирующие ответы. Авторы подчеркивают: конечный результат зависит от точности функционирования дугообразного пучка передающих информацию нервных волокон.

Рис. 17.2. Корковые поля, обозначенные по карте Бродмана, связанные с высокими показателями выполнения тестов интеллекта и мышления. Эти поля образуют теменно-лобную интеграционную систему; темные кружки — доминирующие центры левого полушария; серые кружки — доминирующие двусторонние центры; белая стрелка — дугообразный пучок, связывающий центры
Брока и Вернике (по Jung, Haier, 2007)
Межполушарные отношения. Проблема специализации полушарий в познавательной деятельности человека хорошо изучена (см. п. 5.4 и 8.5). Аналитическая, знаково опосредованная стратегия познания характерна для работы левого полушария, синтетическая, образно опосредованная — правого. Закономерно, что функциональные свойства полушарий, а точнее, степень их индивидуальной выраженности, могут служить физиологическим условием высоких достижений в решении задач разного типа (вербально-логических или пространственных).
Исходно предполагалось, что условием высоких достижений в умственной деятельности является развитие функций доминантного левого полушария, однако в настоящее время все большее значение в этом плане придается функциям субдоминантного правого полушария. Возникла гипотеза эффективного билатерального взаимодействия как физиологической основы общей одаренности. Чем лучше праворукий человек использует возможности своего субдоминантного правого полушария, тем больше он способен: одновременно обдумывать разные вопросы; привлекать больше ресурсов для решения интересующей его проблемы; одновременно сравнивать свойства объектов, вычленяемые познавательными стратегиями каждого из полушарий. Гипотеза билатерального взаимодействия и эффективного использования всех возможностей левого и правого полушарий в интеллектуальной деятельности представляется оптимальной, поскольку она, во-первых, адресуется к работе мозга как целого и, во-вторых, использует представления о ресурсах мозга.
Читайте также:
