
Общая физиология мышечной и нервной системы
Основоположники научных направлений и лауреаты Нобелевской премии в области физиологии
| Уильям Гарвей |

Физиология человека и животных,как наука о жизнедеятельности здорового организма и функциях его составных частей — клеток, тканей, органов и систем, зародилась в XVII столетии. Основоположником экспериментальной физиологии является английский врач, анатом, физиолог и эмбриолог Уильям Гарвей (1578— 1657), который в результате многолетних наблюдений и экспериментов создал учение о кровообращении (см.стр386). История физиологии, как и любой другой области знания, неразрывно связана с именами ученых, своими научными поисками и открытиями способствовавших прогрессу в изучении Природы, в данном случае — жизнедеятельности организма человека и животных. Этим объясняется впервые предпринимаемая попытка представить развитие физиологии в виде совокупности данных, характеризующих вклад известных ученых и лауреатов Нобелевской премии в разработку физиологии клетки, общей физиологии нервной и мышечной систем, физиологии центральной нервной системы, физиологии органов чувств и физиологии висцеральных систем.
Физиология клетки
Выдающимся достижением в физиологии клетки является обоснование в конце 40-х — 50-х годах XX столетия мембранной теории возникновения биоэлектрических потенциалов (А.Ходжкин, Э.Хаксли и Б.Катц). В 1963 г. Нобелевской премии удостоены австралийский нейрофизиолог Джон К.Экклс (р. 1903) и английские физиологи Эндрю Ф.Хаксли (р. 1917) и Алан Л.Ходжкин (р. 1914) за изучение ионных механизмов возбуждения и торможения в периферических и центральных частях оболочек нервных клеток.
Д.Экклс впервые осуществил внутриклеточное отведение электрических процессов в клетках центральной нервной системы, определил электрофизиологические характеристики возбуждающих и тормозящих постсинаптических потенциалов в отдельных нервных клетках, открыл пресинаптическое торможение. Э.Хаксли и А.Ходжкин показали роль ионов натрия в генезе мембранного потенциала действия, а также установили, что в состоянии покоя концентрация ионов калия внутри нервной клетки выше, чем снаружи, а концентрация ионов натрия, наоборот, выше снаружи. Ходжкин впервые измерил абсолютную величину мембранного потенциала и описал динамику изменений этой величины во время генерации нервного импульса. Хаксли принадлежит открытие ныне широко известного натриевого насоса в механизме генерации и передачи нервного импульса, создание теории мышечного сокращения.
В 1974 г. Нобелевская премия присуждена за исследования структурной и функциональной организации клетки. Ее лауреатами стали бельгийские ученые — биолог Альберт Клод (1899- 1983) и биохимик Кристиан Р. Де Дюв (р.1917), а также американский физиолог и цитолог Георг Э.Паладе (р. 1912). Изучая субклеточные фракции, А.Клод показал, что с митохондриями связана активность основных ферментов окисления, а также выделил фракцию субклеточных частиц, обогащенных РНК (микросомы Клода). Р.Де Дюв открыл новый класс субклеточных частиц, названных им лизосомами, выяснил их природу и развил концепцию об их функции, определил участие лизосом в физиологических и патологических процессах в клетке. Г.Паладе принадлежит открытие и описание рибосом.
Российский биохимик Владимир Александрович Энгельгардт (1894-1984) установил (совм.с М.Н.Любимовой), что сократительный белок мышцы — миозин, обладает аденозин-трифосфатазной активностью. Авторы показали, что при взаимодействии искусственно приготовленных миозиновых нитей с АТФ изменяются их механические свойства. Эти данные были развиты американским биохимиком Альбертом Сент-Дьердьи (1893-1986), который обнаружил в мышце белок актин и показал, что актомиозиновые нити укорачиваются под влиянием АТФ. В результате этих открытий и дальнейших исследований выявилось единство принципа функционирования, химической динамики и энергетики обладающих подвижностью различных клеток организма.
Общая физиология нервной и мышечной систем
Итальянский натуралист Джованни А.Борелли (1608-1679) связал процесс сокращения мышц при их движении с деятельностью нервов. Он установил роль межреберных мышц в акте дыхания и впервые представил движение сердца как мышечное сокращение.
В 1771 г. итальянский физик и анатом Луиджи Гальвани (1737-1798) открыл в мышцах электрические токи, которые он назвал "животным электричеством". Ему принадлежит разработка теории,
согласно которой мышцы и нервы заряжены электричеством наподобие лейденской банки. Гальвани является основоположником электрофизиологии.
Впервые охарактеризовал действия электрического тока на возбудимые ткани немецкий физиолог Эмиль Дю Буа-Реймон (1818-1896). Он открыл явление физического электротона, показал, что поперечное сечение нерва электроотрицательно по отношению к его длин-нику (ток покоя), установил, что "отрицательное колебание" тока покоя является выражением деятельного состояния тканей. Ряд открытий принадлежит ученикам Дю Буа- Реймона. Лудимар Германн (1838- 1914) объяснил происхождение токов покоя в нерве и мышце, создал теорию распространения возбуждения по нерву. Он экспериментально определил скорость распространения волны сокращения в мышцах человека. Эдуард Ф.В. Пфлюгер (1829-1910) сформулировал законы физиологического электротона, сокращения и полярный закон, составившие основу представлений о процессах возбуждения в живых тканях. Рудольфу П.Г.Гейденгайну (1834-1897) удалось зарегистрировать выделение тепла при одиночном мышечном сокращении и обнаружить зависимость теплообразования в мышцах от кровообращения, нагрузки, интенсивности раздражения и др. Юлиус Бернштейн (1839-1917) показал, что волна сокращения и ток действия в скелетной мышце распространяются с одинаковой скоростью. В 1902 г. он предложил мембранную теорию происхождения биоэлектрических потенциалов в возбудимых тканях, оказавшую существенное влияние им последующее развитие электрофизиологии.
Немецкий физиолог Герман
| Герман Л.Ф.Гельмгольц |
Л.Ф.Гельмгольц (1821-1894) обнаружил и измерил продолжительность одиночного сокращения мышцы, а также разработал теорию ее длительного тетанического сокращения. Он впервые определил скорость распространения возбуждения в нервах. Измерив теплообразование в мышце при ее сокращении, Гельмгольц заложил основы учения об энергетике мышечной работы. Немецкий физиолог Адольф Фик (1829-1901) показал, что безазотистые вещества, в первую очередь, углеводы (а не белки) являются источником энергии мышечной деятельности.
Проблемы общей физиологии нервной и мышечной систем успешно разрабатывались в России. Николай Евгеньевич Введенский (1852-1922) открыл ритмический характер процесса возбуждения и доказал неутомляемость нерва, установил закономерности оптимума и пессимума частоты и силы раздражения, на основе которых ввел в физиологию понятие лабильности и определил ее для разных тканей. Введенский предложил теорию нервного торможения как качественной модификации процесса воз-
 |
буждения. Ему принадлежит также заслуга открытия периэлектротона и создания учения о парабиозе.
| Николай Евгеньевич Введенский |
Александр Иванович Бабухин (1835-1891) показал, что нервное волокно проводит возбуждение в обоих направлениях (закон двустороннего проведения). Открытие и описание явления католической депрессии связано с работами Бронислава Фортунатовича Ве-риго (1860-1925), который установил, что гальванический ток блокирует проведение импульсов по двигательным и чувствительным нервным волокнам. Василий Яковлевич Данилевский (1852-1939) доказал факт увеличения теплообразования в мышце при ее сокращении. На основе работ Г.Гельмгольца, Р.Гейденгайна, Данилевского и других ученых было сформулировано представление о химических источниках энергии мышечного сокращения.
Василий Юрьевич Чаговец (1873-1941) впервые предложил ионную теорию происхождения электрических явлений в живом организме. Близкие к его теории взгляды высказывал американский физиолог Жак Леб (1859-1924). В 1906 г. Чаговец предложил конденсаторную теорию раздражения тканей и доказал, что возбуждающее действие электрического тока обусловлено конденсаторным накоплением ионов на полупроницаемых мембранах живых тканей.
Нобелевская премия за 1922 г. присуждена английскому физиологу Арчибальду В.Хиллу (1886-1977) и немецкому биохимику Отто Ф.Мейергофу (1884-1951). А.Хиллу принадлежит открытие явления скрытого теплообразования в мышцах, а также определение количества тепла, выделяемого мышцей в состоянии покоя и при сокращении. Совместно с А.Доунингом и Р.Джерардом он обнаружил эффект теплообразования в нерве при его возбуждении. Мейергоф описал связь анаэробного распада и аэробного синтеза углеводов в работающей и покоящейся мышце, проследил путь превращения молочной кислоты (цикл Пастера-Мейергофа). Совместно с немецким биохимиком Карлом Ломаном (1898-1978) Мейергоф открыл аденозинтрифосфорную кислоту (АТФ) — они установили ее формулу и впервые расчитали количество энергии, выделяемой при расщеплении этого соединения. В дальнейшем, АТФ была признана универсальным источником энергии в организме.
Одним из достижений физиологии XX века считается открытие медиаторов (нейротрансмиттеров) и создание учения о химическом механизме передачи нервного импульса в синапсах. Основы этого учения были заложены австрийским физиологом Отто Леей (1873— 1961) и английским физиологом Генри Х.Дейлом (1875-1968), удостоенных в 1936 г. Нобелевской премии "за открытие химической природы передачи нервной реакции".
В 1970 г. Нобелевской премией было отмечено "открытие сигнальных веществ в контактных органах нервных клеток и механизмов их накопления, освобождения и дезактивации". Речь шла об исследованиях, ознаменовавших новый этап в развитии учения о медиаторах, выполненных шведским физиологом Ульфом фон Эйлером (1905- 1983), американским фармакологом Джулиусом Аксельро-дом (р. 1912) и английским физиологом и биофизиком Бернардом Катцом (р. 1911). У.Эйлер, изучая процесс передачи нервных импульсов в синаптической нервной системе, установил, что медиатором в этом процессе служит норадреналин. Д.Аксельрод показал механизм действия веществ, блокирующих проведение нервного импульса в синапсах. Б.Катцу принадлежит открытие механизма выделения ацетилхолина в нервно-мышечной передаче возбуждения. Физиологические свойства нервных волокон и, в частности, закономерности изменения возбудимости и рефрактерности нервов при распространении возбуждения исследовал английский физиолог Кейт Люкас (1879-1916), который доказал, что закон "все или ничего" распространяется и на деятельность нервно- мышечного аппарата.
 |
| Алексей Алексеевич Ухтомский |
Развивая учение Н.Е.Введенского о лабильности и парабиозе Алексей Алексеевич Ухтомский (1875-1942) показал, что лабильность органов и тканей не постоянна, приспособление организмов к меняющимся условиям среды достигается в результате перестройки различных органов и систем на новый уровень лабильности. Александр Филиппович Самойлов (1867-1930) установил, что при передаче импульса в нерве преобладают физические, а в передаточном звене (синапсе) — химические процессы. Он доказал, что в основе центрального торможения лежит выделение химического вещества. Даниил Семенович Воронцов (1886-1965) показал, что возбудимость нерва, утраченная под воздействием одновалентных катионов, восстанавливается анодом, а изменения возбудимости, вызванные применением двухвалентных катионов, восстанавливаются катодом (феномен Воронцова). Воронцову принадлежит открытие так называемой следовой электроотрицательности, развивающейся после потенциала действия нерва, а также доказательство
Согласно современным представлениям, нервная и мышечная ткани могут находиться в трех основных состояниях — физиологическом покое, возбуждении и торможении.
Физиологический покой — это такое состояние, когда ткань или орган не проявляет признаков присущей им деятельности.
Возбуждение — деятельное состояние живой ткани, в которое она приходит под влиянием раздражения.
Торможение — такое состояние, когда деятельность ткани или органа ослабляется или полностью прекращается.
Раздражители.При раздражении мышца приходит в состояние возбуждения, которое распространяется по мышце и вызывает ее сокращение. По биологическому действию раздражители бывают адекватные и неадекватные. Адекватный — это такой раздражитель, к которому данный орган или ткань приспособились в процессе эволюции. Для мышцы адекватным раздражителем является нервный импульс. Неадекватными — будут такие раздражители, действию которых ткань или орган в естественных условиях обычно не подвергаются. Так, сокращение мышцы можно вызвать механическим раздражением (укол, кислота, ток) и т.д.
Все раздражители по своей силе деляг на пороговые, подпороговые и сверхпороговые. Пороговыми называют минимальные раздражители, которые могут вызвать возбуждение. Подпороговые — это раздражители, сила которых меньше пороговой, сверхпороговые — раздражители более сильные, чем пороговые.
Биоэлектрические явления.Возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки (рис. 4.19).
Потенциал покоя.Мембрана нервной или мышечной клетки в состоянии покоя с наружной стороны заряжена положительно, а с внутренней — отрицательно. Разность зарядов между наружной и внутренней поверхностью клеточной мембраны в состоянии физиологического покоя клетки называется потенциалом покоя, или мембранным потенциалом.
Потенциал действия. Под влиянием раздражения проницаемость клеточной мембраны для ионов натрия повышается в сотни
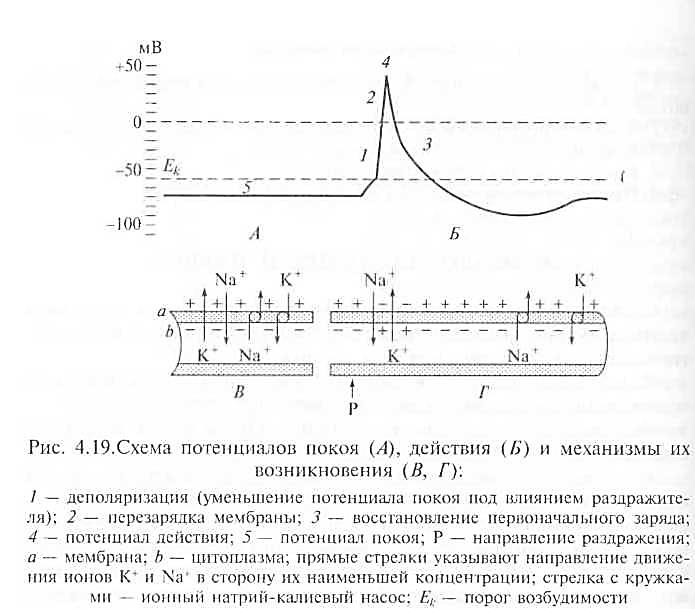 |
Рис. 4.19.Схема потенциалов покоя (А), действия (5) и механизмы их
возникновения (В, Г):
1 — деполяризация (уменьшение потенциала покоя под влиянием раздражите-
ля); 2 — перезарядка мембраны; 3 — восстановление первоначального заряда;
4 — потенциал действия; 5 — потенциал покоя; Р — направление раздражения;
а — мембрана; b — цитоплазма; прямые стрелки указывают направление движе-
ния ионов К + и в сторону их наименьшей концентрации; стрелка с кружка-
ми — ионный натрий-калиевый насос; Ек — порог возбудимости
раз. Происходит перезарядка клеточной мембраны: внутренняя ее сторона заряжается положительно, а наружная — отрицательно. Затем значительно повышается проницаемость мембраны для ионов калия, они начинают быстро перемещаться из клеток во внеклеточную жидкость и вновь восстанавливают первоначальный заряд мембраны. Пикообразное колебание потенциала, возникающее в результате кратковременной перезарядки мембраны и последующего восстановления ее заряда, называется потенциалом действия. Такие колебания потенциала продолжаются 1 — 5 мс.
Проведение возбуждения. Круговой ток, проходя через участок покоя, раздражает его и вызывает появление потенциала действия (рис. 4.20). Когда в одном из участков нервной клетки, нервного или мышечного волокна возникает возбуждение и появляется потенциал действия, между возбужденным и соседним невозбужденным участком возникает разность потенциалов. Эта разность потенциалов является причиной появления круговых (локальных) токов. Внутри клетки ток идет от возбужденного участка к участку покоя, по внешней стороне — от участка покоя к возбужденному.
В участке, который ранее был возбужден, происходит восста
Рис. 4.20. Схема распрост-
ранения возбуждения:
А — волокна мышц и безмякотных нервных волокон; Б —
мякотное нервное волокно; 1 — мембрана; 2 — волокно;
а, Ь, с — перехваты Ранвье; заштрихованы возбужденные
участки; светлые — участки покоя; стрелками показано движение круговых токов

новление потенциала покоя. Данный процесс многократно повторяется и обусловливает распространение импульсов возбуждения. Скорость проведения возбуждения по скелетным мышцам 12 —15 м/с, гладким мышцам — 1 — 18 см/с, в безмякотных нервах — 0,5 — 3 м/с.
Основные свойства живой ткани.Любая живая клетка обладает свойствами раздражимости, возбудимости и лабильности (функциональной подвижности).
Раздражимость. С раздражимостью связаны все процессы роста и размножения клеток и тканей. На разнообразные воздействия внешней или внутренней среды — раздражения протоплазма живой клетки отвечает специфической реакцией, при которой происходят усиление или ослабление обмена веществ, количественное и качественное его изменение. Эта реакция на раздражение получила название раздражимости, или реактивности.
Возбудимость — свойство нервной или мышечной ткани отвечать на действие раздражителя возбуждением. Для измерения величины возбудимости определяют порог возбудимости и хронаксию. Порогом возбудимости называется наименьшая сила раздражителя, способная вызвать возбуждение; его выражают в вольтах постоянного тока.
Лабильность — скорость, с которой в ткани возникает и успевает закончиться полный период отдельного импульса возбуждения. Мера лабильности — это максимальное число импульсов возбуждения за 1 св ответ на такое же максимальное число раздражений. Высокой лабильностью обладают мякотные нерпы, у без- мякотных нервов она низкая. Так, максимальный ритм возбуждений мякотного нервного волокна 500, безммкотного — 200. Максимальный ритм возбуждений скелетной мышцы 200 импульсов в 1 с, а гладкой мышцы — в десятки раз меньше.
При очень частых раздражениях сокращения мышцы уменьшаются или даже совсем прекращаются. Такая частота называется пессимальной, или пессимумом. Пессимум возникает вследствие того, что возбуждение еще не закончилось и ткань находится в состоянии абсолютной или относительной рефрактерное™, а на нее действует новое раздражение. Частые раздражения, превышающие меру лабильности, вызывают не возбуждение, а торможение.
Учение Н. Е. Введенского о парабиозе устанавливает связь между возбуждением и торможением. Возбуждение и торможение — это различные реакции ткани на раздражение, исход которогозависит от лабильности. При высокой лабильности возникает возбуждение, снижение лабильности вызывает торможение.
ФИЗИОЛОГИЯ МЫШЦ
Скелетные мышц, входящие в состав опорно-двигательного аппарата позвоночных животных, состоят из группы мышечных пучков, каждый из них составлен из тысяч мышечных волокон, которые представляют собой клетки цилиндрической формы длиной до 12 см и диаметром 10 — 100 мкм. Каждое волокно окружено оболочкой сарколеммой и содержит тонкие нити — миофибрил-лы (рис. 4.21). Поперечные мембраны делят каждую миофибриллу на отдельные участки — саркомеры. Сократимым веществом мышечного волокна являются миофибриллы, состоящие из множества (около 2500) тонких и толстых белковых нитей — протофибрилл. Толстые протофибриллы образованы из белка миозина, тонкие — из актина. Нити актина прикреплены к мембране саркомера; они образуют светлые участки миофибриллы. В темных участках находятся нити миозина. Акгиновые нити частично входят своими концами в промежутки между миозиновыми нитями. Нити актина и миозина соединяются между собой многочисленными поперечными мостиками, которые образованы скрученными в спираль отростками — мостиками миозиновой нити. Чередование нитей в миофибрилле обусловливает ее поперечную исчерченность.

Рис. 4.21. Схема чередова-
ния нитей в миофибрилле:
А — электронно-микроскопическая фотография мио-
фибриллы; Б — строение миофибриллы; Z — поперечная
мембрана, разделяющая миофибриллу на отдельные уча-
стки — саркомеры; 1 — миозиновая нить, 2 — актиновая нить
Гладкие мышцы, образующие стенки внутренних органов, состоят из клеток различной формы длиной от 30 до 500 мкм и диаметром 2—10 мкм; у клеток имеются боковые отростки, благодаря которым клетки гладких мышц группируются в длинные пучки. Они, в свою очередь, при помощи тяжей соединяются друг с другом, обеспечивая деятельность мышцы как единой системы. Сократительным аппаратом гладких мышц являются миофибриллы, в которых содержатся тонкие актиновьге нити с прикрепленными к ним короткими отростками миозиновых нитей, называемых димерами.
Свойства скелетных мышц.Проведение возбуждения в мышцах происходит изолированно, т. е. потенциалы действия не переходят с одного мышечного волокна на другое. Возбудимость скелетныхмышц меньше возбудимости нервов. Нервное волокно оканчивается в середине мышечного волокна, поэтому возбуждение распространяется в обе стороны со скоростью 4—15 м/с.
Скелетная мышца является упругим телом. Если к мышце подвесить груз, то она растягивается, это свойство называется растяжимостью. Эластичностью мышцы называется возвращение мышцы к первоначальной своей длине после растяжения, когда будет удален груз. Пластичностью мышцы называется свойство сохранять удлиненную форму после удаления груза, вызвавшего ее растяжение. В скелетных мышцах лучше выражено свойство эластичности, чем пластичности.
Виды мышечных сокращений.При нанесении на мышцу одиночного раздражения она отвечает одиночным сокращением. Записывая это сокращение на двигающейся ленте какого-либо самописца (например, в простейшем случае на кимограф), можно отметить три периода: латентный — период от раздражения до начала сокращения, период сокращения и период расслабления. У икроножной мышцы лягушки одиночное сокращение длится 0,1 с, у млекопитающих — 0,04 — 0,1 с. В латентный период в мышце происходят процессы, при которых освобождается энергия для мышечного сокращения.
Если к мышце поступает несколько частых импульсов возбуждения, наступает длительное сокращение мышцы, которое называется тетаническим, или тетанусом. В зависимости от частоты возбуждений тетанус будет зубчатым или гладким (рис. 4.22).
Зубчатый тетанус наблюдается при такой частоте импульсов возбуждений, когда каждый импульс действует на мышцу в тот момент, когда она уже начинает расслабляться. Если же импульсы возбуждения настолько частые, что они воздействуют на мышцу до начала ее расслабления, то получатся длительные непрерывные сокращения мышцы — гладкий тетанус.
 |
Рис. 4.22. Сокращение скелетной мышцы при различной частоте раздра-
жения:
А — одиночное сокращение в ответ на одно раздражение; Б — зубчатый тетанус
(до 15 раздражений в 1 с); В — гладкий тетанус (свыше 25 раздражений); М —
механограмма — запись сокращения мышц; Р — частота раздражений; 1 — фаза
сокращения; 2 — фаза расслабления
Механизм мышечного сокращения (теория скольжения).В состоянии покоя тонкие нити актина лежат частично своими концами в промежутках между толстыми миозиновыми нитями. При возбуждении мышцы под влиянием потенциала действия нити актина сдвигаются в промежутке между нитями миозина, т. е. скользят навстречу друг другу, вызывая сокращение миофибрилл (рис. 4.23). Считают, что скольжение происходит в результате укорочения мостиков — боковых отростков миозиновых нитей под влиянием ионов кальция. Укорачиваясь, мостики тянут актиновую нить навстречу миозиновой.
Работа и сила мышц.При сокращении мышца укорачивается и тем самым совершает работу. Она будет наибольшей при средних нагрузках и среднем ритме сокращений.
Силу мышцы определяют по максимальному сокращению ее при поднятии максимального груза. Наибольшей силой обладают мышцы с косыми волокнами, имеющими перистое строение.
Утомление мышц.В процессе сокращений мышцы утомляются, при этом понижаются их возбудимость, лабильность и величина сокращения. В утомленной мышце понижается содержание гликогена и накапливаются продукты обмена. В опытах на нервно-мышечном препарате Н. Е. Введенский установил, что прежде всего утомляются синапсы в связи с их низкой лабильностью. В целом организме утомление в первую очередь наступает в нервных центрах коры больших полушарий. И. М.Сеченов доказал, что быстрое восстановление работоспособности утомленных мышц наступает не при полном их покое, а при работе других, до этого не сокращавшихся мышц. Импульсы от вновь вовлеченных в работу мышц повышают возбудимость нервных центров, возбуждение одних нервных центров снижает и даже снимает утомление других центров. Утомление зависит от состояния симпатической нервной системы и желез внутренней секреции: утомленная мышца вновь
Рис. 4.23. Схема расслабленной (А) и сократившейся (Б) миофибриллы
 |
начинает сокращаться при раздражении симпатического нерва или введении адреналина, усиливающих обмен. Тренировка мышц позволяет отсрочить наступление утомления. При ней увеличивается объем мышц в результате утолщения мышечных волокон, повышается содержание гликогена, АТФ и креатинфосфата, ускоряются восстановительные процессы.
Тонус мышц. Тонус скелетных мышц играет важную роль для поддержания определенного положения тела в пространстве и деятельности двигательного аппарата. Скелетные мышцы в покое расслабляются не полностью, а находятся в некотором напряжении, т.е. тонусе. Он обусловлен поступлением к мышцам редких нервных импульсов, возбуждающих мышечные волокна не одновременно, а поочередно.
Гладкие мышцы обладают способностью длительно находиться в тонусе. Длительные тонические сокращения этих мышц особенно отчетливо выражены в сфинктерах полых органов, в стенках кровеносных сосудов.
Классификация мышц.
Скорость передвижения животных, возможность сохранять позу, осуществлять гомеостатическое регулирование кровообращения, дыхания и других процессов зависит от мышечной системы.
Все мышцы организма являются эффекторами и служат объектом иннервации.
Все мышцы делят на скелетные, (поперечнополосатые, исчерченные) и гладкие.
Поперечнополосатые мышцы формируют двигательные аппараты скелета, глазодвигательный, жевательный и др. аппараты. К этой же группе относится сердечная мышца. Скелетные мышцы подвержены волевому контролю, управление их происходит соматической нервной системой. Сердечная мышца управляется автономной нервной системой, иннервирована симпатическими и парасимпатическими нервными волокнами, и не подлежит произвольному контролю.
Гладкие мышцы внутренних органов желудка, кишечника, сосудов, обладают автоматизмом в работе, иннервированы автономной нервной системой и не могут управляться произвольно.
Поперечнополосатые мышцы скелета позвоночных животных состоят из множества отдельных волокон, которые расположены в общей соединительнотканной оболочке и крепятся к сухожилиям, связанным со скелетом. Имеются мышцы параллельно-волокнистого и перистого типа строения. Сила мышцы пропорциональна площади физиологического поперечного сечения.
Каждое волокно скелетной мышцы – это многоядерное клеточное образование, симпласт, сложенный объединением нескольких миобластов. Диаметр симпластного мышечного волокна не превышает 100 мкм, длина составляет 2-3 см.
Основной структурный элемент мышечного волокна – миофибриллы. Миофибриллы образуют структуру с чередованием светлых и темных дисков, пространственно совпадающих у многих волокон в мышце, что и дало основание для термина исчерченные.
В соответствии со схемой Хаксли, а также биохимическими исследованиями, миофибриллы содержат сократительные белки. Толстые и тонкие нити миозина и актина вдвигаются в промежутки между собой за счет скольжения в присутствии АТФ и ионов Са ++ .
Миозин - сократительный белок, массой 490000, имеет две фракции, легкого и тяжелого миозина, 2 альфа-спирали. Длина молекулы до 1400 ангстрем. По массе составляет 54% от массы мышцы.
Актин, белок с массой 46000, образует одну закрученную цепь, состоящую из двух спиралей. Различают G-актин, глобулярный, и F-актин, составленный из отдельных глобул. По массе занимает 21% мышцы.
Тропомиозин (нить) и тропонин (глобула), выполняют регуляторную роль, без кальция препятствуют сокращению, составляют 15% массы мышцы.
Миозиновые и актиновые нити составляют основу сократительного механизма. Кроме нитей, в мышечном волокне имеется тубулярная система, или система канальцев. Образована впячиваниями наружной мембраны (Т-канальцы) и саркоплазматическим ретикулумом – сетью продольно ориентированных каналов и цистерн между миофибриллами.
Поперечнополосатые мышцы способны сокращаться в двух режимах – изотоническом (постоянная тяга) и изометрическом (постоянная длина), хотя эти понятия по механизму перекрываются. Ауксотонический режим более часто встречается, при сокращении меняется и тяга, и геометрия мышцы.
Исчерченные мышцы обладают широким диапазоном скоростей сокращения, поэтому их можно делить на быстрые (фазические) и медленные (тонические).
Крикотиреоидная мышца летучей мыши, 200 сокр/с, сгибатели крыльев колибри 45 сокр/с, у зяблика 25 сокр/с. Глазодвигательные мышцы человека при саккадах сокращаются за 5-6 мс. Мышцы кузнечиков способны давать 500 сокр/с.
Скорость сокращения мышц зависит от их биохимических свойств и иннервации. У позвоночных мышцы бывают богатыми миоглобином (депо кислорода) и белыми. Красные миоглобиновые мышцы сокращаются медленно и не так скоро утомляются. По набору ферментов белые мышцы приспособлены к анаэробному гликолизу, а красные к окислительному фосфорилированию. Для красных мышц характерен запас липидов, много крупных митохондрий, высокая активность фосфорилазы, цитохромоксидазы, сукцинатдегидрогеназы. Для белых быстрых мышц характерно малое число митохондрий, много гликогена, высокая активность мышечной лактатдегидрогеназы. Сердечная мышца относится к красному типу, содержит мало гликогена, но много ферментов аэробного обмена.
В любых мышцах имеются быстрые и медленные волокна.
Как правило, быстрые мышечные волокна иннервированы более толстыми нервными волокнами, они имеют более высокий потенциал покоя. У медленных мышечных волокон обнаруживается большая емкость, и высокое удельное сопротивление мембраны, что дает большую постоянную времени. Тонические медленные мышцы получают много двигательных нервных пресинаптических окончаний, поэтому возбуждение обеспечивается нервным механизмом.
Медленные волокна сокращаются градуально, продолжительно м с меньшим утомлением. Нервный импульс и последующее высвобождение АХ в синапсе запускает процесс сокращения, ПКП совсем не обязательно приводит к ПД.
Быстрые мышцы сокращаются чаще после генерации ПД в мышечной мембране.
Одиночный тип иннервации мышечных волокон бывает в быстрых мышцах, аксон следует от крупного мотонейрона. Множественный тип иннервации мышечных волокон характерен для медленных, тонических мышц. На одном мышечном волокне расположены синапсы, образованные несколькими малыми мотонейронами. ПКП от разных синапсов суммируются и обеспечивают градуальную деполяризацию.
Мембранный потенциал покоя мышечного волокна определяется двумя ионами, находящимися в определенных соотношениях, ионами K + и Cl - . Мембрана мышечной клетки проницаема для этих двух ионов в состоянии покоя. Мембранный потенциал определяется суммой концентраций электрогенных катионов и анионов, но зависит от K + и Cl - в большей степени, чем от других. Градиенты калия и хлора (калия больше в саркоплазме, хлора в интерстиции) выравнены таким образом, что пока не произойдет изменение концентрации одного из них относительно другого, мембранный потенциал сдвинуться не может (правило Доннана). Например, рост концентрации К + во внеклеточной среде не снижает мембранный потенциал немедленно, сначала в волокно должен поступить хлор, чтобы выровнять градиент концентраций анионов и катионов по обе стороны клеточной мембраны.
Но механизм генерации потенциала концевой пластинки и потенциала действия сходен с тем, который реализуется и в других возбудимых клетках. Большая емкость мембраны обусловливает большую амплитуду ПД, большую его длительность и большой рефрактерный период. Большие токи, генерируемые мышцами, дают возможность регистрировать миограмму с поверхности тела.
Электромеханическое сопряжение. Для чего нужен потенциал концевой пластинки или ПД в мышце? Для осуществления сокращения. Необходимым условием сокращения мышцы, то есть условием перемещения актиновых и миозиновых нитей относительно друг друга, является повышение в саркоплазме концентрации ионов Са ++ .
Считается, что на 1 шаг перемещения сократительных белков тратится энергия 1 молекулы АТФ, роль АТФ-азы играет актомиозин, т.е. комплекс актина и миозина. Необходимо присутствие Са ++ и Mg ++ . АТФ тратится и на расслабление мышцы, для работы кальциевых ионных насосов, перекачивающих кальций во внутриклеточные депо. Механизм расслабления мышц изучен слабее, чем механизм сокращения. Есть мнение, что мышцы расслабляются из-за действия внешних сил (активность мышц-антагонистов) и пластификации.
Мышцы способны сокращаться в нескольких режимах, в зависимости от частоты прихода управляющих стимулов. При 15 Гц регистрируется зубчатый тетанус, при частоте, превышающей 25 Гц – гладкий. Одиночные сокращения происходят при частотах, близких к 1 Гц. Для каждой из мышц имеется оптимум и пессимум частоты, при оптимальной частоте стимуляции она укорачивается на 20% первоначальной длины и развивает максимальное усилие. При этом часть волокон может сокращаться по гладкому тетанусу, часть - по зубчатому.
Сила и работа мышц.
Работа мышцы, как и всякого физического тела, определяется способностью поднять определенный груз (Р) на известную высоту h:
Одиночное волокно может развивать тягу 100-200 мг, вся мышца- несколько кг.
Из курса биохимии известно, что 1 моль АТФ дает 48 кДж энергии, для его ресинтеза нужно 3 моля кислорода. При тяжелой физической работе запасов О2 в мышцах мало для немедленного и достаточного ресинтеза АТФ. Поэтому начинается мобилизация анаэробного распада креатинфосфата и гликолиза, недоокисленные продукты (прежде всего молочная кислота) накапливаются. Создается кислородная задолженность, которая погашается системными реакциями – увеличением кровотока, учащением сердцебиений и ростом частоты дыхания. При тренировке возможна компенсация за счет правильного дыхания и кровообращения. Но чаще развивается утомление, прежде всего нервных центров, а затем и мышц. Утомление – в значительной степени результат кислородного дефицита.
Особенности гладких мышц.
Сила сокращений гладких мышц в пересчете на площадь физиологического поперечного сечения сопоставима с силой скелетных. Скорость сокращения и расслабления может быть на 2 порядка меньше.
| | | следующая лекция ==> | |
| ГАМК, гамма-аминомасляная кислота, наиболее характерна для тормозных синапсов головного мозга, в спинном мозге чаще используется глицин | | | Лекция 7 Общая физиология центральной нервной системы |
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также:
