
Особенности проведения нервного импульса по нервному волокну
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
Потенциалы действия (импульсы возбуждения) обладают способностью распространяться вдоль по нервным и мышечным волокнам. Потенциалы могут быть локальными, они распространяются на небольшие расстояния 1-2 мм с затуханием (декрементом) и импульсными. Импульсные потенциалы распространяются без декремента на значительные расстояния – до нескольких десятков сантиметров. Локальные потенциалы возникают в ответ на действие подпорогового раздражителя, например, на мембране рецепторной клетки. Если локальное возбуждение попадает в участок мембраны, способной генерировать ПД, и амплитуда локального тока достигает критического уровня деполяризации, формируется ПД, который распространяется по всей длине нервного волокна.
Передача информации на большие расстояния в пределах нервной системы осуществляется с помощью нервных импульсов по аксонам нейронов. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциал чувствительных ионных каналов. В зависимости от расположения и концентрации ионных каналов в мембране волокна выделяют два способа проведения нервного импульса.
1. Непрерывное проведение нервного импульсаосуществляется в безмиелиновых волокнах, объясняется равномерным распределением потенциал чувствительных ионных каналов, участвующих в генерации ПД.
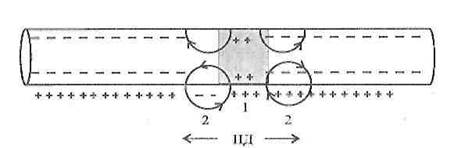
Рисунок 3 - Непрерывное распространение ПД в нервном волокне
Возникший ПД обеспечивает открытие потенциал зависимых Na-каналов на соседнем участке мембраны нервного волокна и движение ионов Na + внутрь волокна, что обеспечивает развитие критического уровня деполяризации на соседнем участке нервного волокна и возникновение нового ПД. Непрерывное распространение нервного импульса идет через генерацию новых импульсов по эстафете, когда каждый возникший импульс является раздражителем для соседнего участка нервного волокна и обеспечивает возникновение нового ПД.
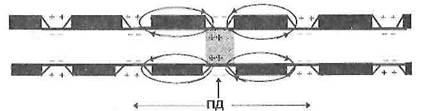
Рисунок 4 – Сальтаторное распространение ПД в нервных волокнах
2. Сальтаторное проведение нервного импульса (ПД) осуществляется в миелиновых волокнах, так как у них потенциал чувствительные ионные каналы локализованы только в участках мембраны перехватов Ранвье, где их плотность достигает 12 000 на 1 мкм 2 . В области межузловых сегментов, обладающих высокими изолирующими свойствами, потенциал чувствительных каналов нет, вследствие чего мембрана осевого цилиндра там практически невозбудима. Поэтому ПД, возникший в одном перехвате Ранвье распространяется через межузловой сегмент до соседнего перехвата, деполяризует мембрану до критического уровня и вызывает возникновение потенциала действия.
Сальтаторное проведение нервных импульсов является эволюционно более поздним механизмом, возникшим впервые у позвоночных в связи с миелинизацией нервных волокон. Оно имеет два важных преимущества по сравнению с непрерывным механизмом проведения возбуждения:
-более экономично по затрате энергии, так как возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% от площади мембраны волокна, следовательно, требуется меньше энергии для восстановления трансмембранных градиентов ионов натрия и калия;
-возбуждение проводится с большей скоростью (до 120 м/с), чем в безмиелиновых волокнах (0,5-2,0 м/с).
В связи с этими приемуществами миелиновые волокна в нервной системе сформировались там, где необходима наиболее быстрая регуляция функций. Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей.
Проведение нервных импульсов по волокнам нейронов починяется определенным законам:
Закона 1: нервный импульс распространяется в обе стороны от места раздражения.
Закон 2: проведение нервного импульса по волокну происходит изолировано и не распространяется на параллельные волокна. Объяснение этого закона заключается в том, что аксолемма имеет очень высокое сопротивление и не пропускает петли тока на невозбужденные волокна, расположенные рядом. Изолированное проведение обеспечивает высокую точность регуляторной деятельности ЦНС.
Закон 3: скорость проведения возбуждения по нервному волокну определяется его диаметром. Отсюда следствие: чем толще нервное волокно, тем больше скорость проведения нервного импульса по этому волокну.
Закон 4: нерв сохраняет способность к проведению возбуждения в течение 6-8 часов непрерывного раздражения (закон Н.Е. Введенского, 1883).
Закон 5: действие веществ, блокирующих работу ионных каналов, без нарушения целостности нервного волокна вызывает состояние обратимого парабиоза, (закон парабиоза, Введенский, 1901).
Кроме того, выделяют несколько видов распространения возбуждения:
1. Иррадиация (дивергенция) возбуждения в ЦНС, которая объясняется ветвлением аксонов и наличием вставочных нейронов, аксоны которых также ветвятся. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов.
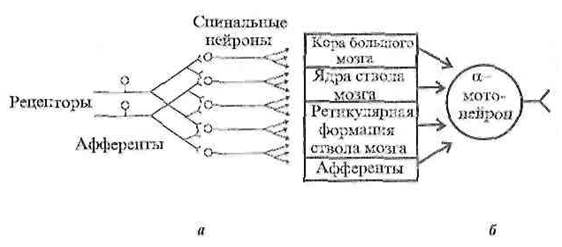
Рисунок 4 – Дивергенция афферентных дорсальных корешков на нейроны головного мозга (а – дивергенция, б – конвергенция)
2. Конвергенция возбуждения представляет собой схождение нескольких нервных импульсов, идущих по разным путям к одному и тому же нейрону. Явление конвергенции распространения возбуждения описал Э. Шеррингтон, поэтому явление было названо принцип шеррингтоновской воронки или принцип общего конечного пути. Примером может служить конвергенция возбуждения на спинальном мотонейроне. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг.
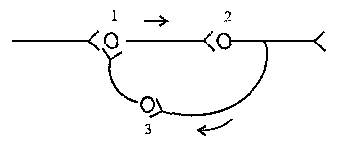
Рисунок 5 – Циркуляция возбуждения в замкнутых нейронных цепях
3. Циркуляция возбуждения по замкнутым нейронным цепям, возникает в результате замыкания группы нейронов в кольцевую структуру. Циркуляция возбуждения – одна из причин явления последействия. Считают, что циркуляция возбуждения в замкнутых нейронных цепях наиболее вероятный механизм феномена кратковременной памяти. Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом.
Литература
1. Смирнов, В.М. Физиология сенсорных систем и высшая нервная деятельность / В.М. Смирнов, С.М. Будылина. - М.: Медицина, 2003. - 304 с.
2. Шульговский, В. В. Основы нейрофизиологии: Учебное пособие для студентов вузов. - М.: Аспект Пресс, 2000. - с. 277.
3. Батуев, А.С. Физиология поведения. Нейрофизиологические закономерности / А.С. Батуев. - Л.: Наука, 1986. - 340 с.
4. Александров, Ю.И. Психофизиология / Ю.И.Александров. - М.: Медицина, 2001. - 230 с.
Данилова, Н.Н. Физиология высшей нервной деятельности / Н. Н. Данилова, А.Л. Крылова. - Ростов н/Д: Феникс, 1999. – 480 с.
[1] ионоселективные каналы и ионные насосы
[2] правило действует и в обратном направлении
[3] 1780 год, профессор анатомии Болонского университета
Синапс, его строение. Виды синапсов. Механизм проведения возбуждения через синапсы. Постсинаптическое и пресинаптическое торможение.
Синапсы - это специализированная структура, которая обеспечивает передачу нервного импульса из нервного волокна на эффекторную клетку - мышечное волокно, нейрон или секреторную клетку.
Синапсы - это места соединения нервного отростка (аксона) одного нейрона с телом или отростком (дендритом, аксоном) другой нервной клетки (прерывистый контакт между нервными клетками).
Все структуры, обеспечивающие передачу сигнала с одной нервной структуры на другую - синапсы.
Значение - передает нервные импульсы с одного нейрона на другой => обеспечивает передачу возбуждения по нервному волокну (распространение сигнала).
Большое количество синапсов обеспечивает большую площадь для передачи информации.
- 1. По локализации: центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный синапс вегетативной нервной системы). Центральные синапсы можно в свою очередь разделить на аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и т.п. Согласно Г. Шенсрду, различают реципрокные синапсы, последовательные синапсы и синаптические гломерулы (различным способом соединенные через синапсы клетки).
- 2. По развитию в онтогенезе: стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
- 3. По конечному эффекту: тормозные и возбуждающие.
- 4. По механизму передачи сигнала: электрические, химические, смешанные.
- 5. Химические синапсы можно классифицировать:
- а) по форме контакта - терминальные (колбообразное соединение) и преходящие (варикозное расширение аксона);
- б) по природе медиатора - холинергические (медиатор - ацетилхолин, АХ), адренергическис (медиатор - норадреналин, НА), дофаминергические (дофамин), ГАМК-ергические (медиатор - гаммааминомасляная кислота), глицинергические, глутаматергические, аспартатсргические, пептидергические (медиатор - пептиды, например, вещество Р), пуринергические (медиатор - АТФ).
Строение синапса:
- 1. Пресинаптическая мембрана - принадлежит нейрону, ОТ которого передается сигнал.
- 2. Синаптическая щель, заполненная жидкостью с высоким содержанием ионов Са.
- 3. Постсинаптическая мембрана - принадлежит клеткам, НА которые передается сигнал.
Между нейронами всегда существует перерыв, заполненный межтканевой жидкостью.
В зависимости от плотности мембран, выделяют:
- - симметричные (с одинаковой плотностью мембран)
- - асимметричные (плотность одной из мембран выше)
Пресинаптическая мембрана покрывает расширение аксона передающего нейрона.
Расширение - синаптическая пуговка/синаптическая бляшка.
На бляшке - синаптические пузырьки (везикуль).
С внутренней стороны пресинаптической мембраны - белковая/гексогональная решетка (необходима для высвобождения медиатора), в которой находится белок - нейрин. Заполнена синаптическими пузырьками, которые содержат медиатор - специальное вещество, участвующее в передаче сигналов.
В состав мембраны пузырьков входит - стенин (белок).
Пузырьки содержат молекулы медиатора (внутри) - вещество, необходимое для передачи сигнала.
Механизм проведения возбуждения через синапсы.
В состоянии покоя в пресинаптической области везикулы постоянно движутся. Они подходят к пресинаптической мембране, выделяя в синаптическую щель медиатор, т. е. в состоянии покоя выделяются несколько квантов медиатора. Он дефилирует через щель, взаимодействует с холинорецепторами. Но т. к. медиатора мало, то потенциал на постсинаптической мембране тоже мал. Это миниатюрные потенциалы с амплитудой в несколько микровольт. Эти потенциалы поддерживают фоновую активность постсинаптической мембраны. Под действием раздражителя возникает потенциал действия нервного волокна. Под действием круговых токов повышается проницаемость мембраны нервного волокна для Са2+, в результате Са2+ поступает внутрь нервного волокна, вызывая упорядоченное движение везикул и, как следствие, увеличение количества медиатора, выделяющегося в синаптическую щель.
На постсинаптической мембране взаимодействие медиатора с рецепторами открывает Nа-каналы, Nа поступает внутрь клетки. Возникает возбуждающий постсинаптический потенциал. Этот потенциал - особая форма возбуждения, которая не распространяется (свойство местного возбуждения). Затем холинэстераза расщепляет медиатор, освобождая рецептор - в результате восстанавливается заряд постсинаптической мембраны.
Постсинаптическое торможение
Торможение - это активный нервный процесс, результатом которого является прекращение или ослабление возбуждения. Торможение вторично относительно процесса возбуждения, так как всегда возникает как следствие возбуждения.
Торможение в ЦНС открыл И.М. Сеченов (1863). В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Было показано, что латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Открытие И.М. Сеченова послужило толчком для дальнейших исследований торможения в ЦНС. В частности, Ф. Гольц (1870) обнаружил проявления торможения у спинальной лягушки. Он также исследовал латентное время рефлекса. Оказалось, что механическое раздражение кончиков пальцев одной конечности лягушки существенно удлиняет латентный период сгибательного рефлекса другой конечности при погружении ее в раствор кислоты.
Наличие специальных тормозных структур в продолговатом мозге доказал Х. Мегун (1944).
В опытах на кошках при изучении разгибательного рефлекса он установил, что раздражение медиальной части ретикулярной формации продолговатого мозга тормозит рефлекторную активность спинного мозга.
Тонкий анализ тормозных явлений в ЦНС позволил выделить две разновидности торможения:
- 1) постсинаптическое торможение и
- 2) пресинаптическое торможение.
Этот вид торможения открыл Д. Экклс (1952) при регистраци потенциалов мотонейронов спинного мозга у кошки во время сокращения и расслабления мышцы в ходе реализации соответствующих рефлекторных актов. Оказалось, что при рефлекторном расслаблении мышц на мотонейронах регистрируется гиперполяризационный постсинаптический потенциал, уменьшающий возбудимость мотонейрона, угнетающий его способность реагировать на возбуждающие влияния. Поэтому вызванный гиперполяризационный потенциал был назван тормозным постсинаптическим потенциалом (ТПСП).
Пресинаптическое торможение
Пресинаптическое торможение первоначально выявлено в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов.
В очаге деполяризации нарушается процесс распространения возбуждения, следовательно, поступающие импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуде, не обеспечивают выделение медиатора в синаптическую щель в достаточном количестве: нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным.
Разновидности пресинаптического торможения. Обычно выделяют параллельное и латеральное пресинаптическое торможение. Структурная организация этого торможения внешне похожа на такие же варианты постсинаптического торможения. Однако механизм пресинаптического торможения отличается принципиально - возбуждение задерживается не на теле нейрона, а на подступах к нему. В случае постсинаптического торможения заторможен нейрон - снижена возбудимость тела нейрона (обычно вследствие его гиперполяризации).
Локализация в ЦНС пре- и постсинаптического торможения. Оба механизма торможения широко представлены в различных отделах ЦНС: в частности, глицин - медиатор постсинаптического торможения, кроме клеток Реншоу, имеется в ретикулярной формации ствола мозга; ГАМК - в спинном мозге, мозжечке, коре большого мозга. Оба вида торможения могут быть заблокированы: постсинаптическое - стрихнином; пресинаптическое - бику-кулином. Пост- и пресинаптическое торможение блокируется также столбнячным токсином, который нарушает высвобождение тормозных медиаторов.
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности(непрерыаности волокна)
Проведение импульсов по нервномуволокну возможно лишь в том случае, если не нарушена его целостность.( К-е каналы блокируются)
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждениепередается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняетмиелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждениепередается изолированно.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.(осуществляется через синопс)
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
Нервные волокна представляют собой отростки нервных клеток (дендриты, аксоны), покрытые оболочками. При этом отросток в каждом нервном волокне является осевым цилиндром, а окружающие его нейролеммоциты (шванновские клетки), относящиеся к нейроглии, образуют оболочку волокна - нейролемму. С учетом строения оболочек нервные волокна подразделяют на безмякотные (безмиелиновые) и мякотные (миелиновые). Безмиелиновые нервные волокна имеются, главным образом, у вегетативных нейронов. Осевой цилиндр как бы прогибает плазматическую мембрану (оболочку) нейролеммоцита, которая смыкается над ним. Сдвоенная над осевым цилиндром мембрана нейролеммоцита получила название мезаксон. Под шванновской клеткой остается узкое пространство (10-15 нм), содержащее тканевую жидкость, участвующую в проведении нервных импульсов. Один нейролеммоцит окутывает несколько (до 5-20) аксонов нервных клеток. Оболочку отростка нервной клетки образуют многие шванновские клетки, располагающиеся последовательно одна за другой. Миелиновые нервные волокна толстые, они имеют толщину до 20 мкм. Эти волокна образованы довольно толстым аксоном клетки - осевым цилиндром. Вокруг аксона имеется оболочка, состоящая из двух слоев.
17. Рецепторы: понятия, классификация, основные свойства и особенности, механизм возбуждения, функциональная мобильность.
Рецептор – это специализированная структура (клетка или окончание нейрона), которая в процессе эволюции приспособилась к восприятию соответствующего раздражителя внешнего или внутренного мира.
Рецепторами называются специальные образования, трансформирующие энергию внешнего раздражения в специфическую энергию нервного импульса.
По положению в организме:
· Экстерорецепторы (экстероцепторы) — расположены на поверхности или вблизи поверхности тела и воспринимают внешние стимулы (сигналы из окружающей среды)
· Интерорецепторы (интероцепторы) — расположены во внутренних органах и воспринимают внутренние стимулы (например, информацию о состоянии внутренней среды организма)
· Проприорецепторы (проприоцепторы) — рецепторы опорно-двигательного аппарата, позволяющие определить, например, напряжение и степень растяжения мышц и сухожилий. Являются разновидностью интерорецепторов.
По способности воспринимать разные стимулы:
· Мономодальные — реагирующие только на один тип раздражителей (например, фоторецепторы — на свет)
· Полимодальные — реагирующие на несколько типов раздражителей (например. многие болевые рецепторы, а также некоторые рецепторы беспозвоночных, реагирующие одновременно на механические и химические стимулы).
По адекватному раздражителю:
· Хеморецепторы — воспринимают воздействие растворенных или летучих химических веществ.
· Механорецепторы — воспринимают механические стимулы (прикосновение, давление, растяжение, колебания воды или воздуха и т. п.)
· Фоторецепторы — воспринимают видимый и ультрафиолетовый свет
· Терморецепторы — воспринимают понижение (холодовые) или повышение (тепловые) температуры
По структурным особенностям различают первичные и вторичные рецепторы.
Первичные рецепторы — это окончания чувствительных биполярных клеток, тело которых находится вне ЦНС, один отросток подходит к воспринимающей раздражение поверхности, а другой направляется в ЦНС.
Вторичные рецепторы представлены специализированными рецепторными клетками, которые расположены между чувствительным нейроном и точкой приложения раздражителя.
· Избирательность - чувствительность к адекватным раздражителям
· Возбудимость - минимальной величиной энергии адекватного раздражителя, которая необходима для возникновения возбуждения, т.е. порогом возбуждения.
· Низкая величина порогов для адекватных раздражителей
· Адаптация (может сопровождаться как понижением, так и повышением возбудимости рецепторов. Так, при переходе из светлого помещения в темное происходит постепенное повышение возбудимости фоторецепторов глаза, и человек начинает различать слабо освещенные предметы— это так называемая темновая адаптация.)
Первично-чувствующие рецепторы: раздражитель действует на дендрит сенсорного нейрона, изменяется проницаемость клеточной мембраны к ионам (в основном к Na+), образуется локальный электрический потенциал (рецепторный потенциал), который электротонически распространяется вдоль мембраны к аксону. На мембране аксона образуется потенциал действия, передаваемый далее в ЦНС.
Сенсорный нейрон с первично-чувствующим рецептором представляет собой биполярный нейрон, на одном полюсе которого располагается дендрит с ресничкой, а на другом – аксон, передающий возбуждение в ЦНС. Примеры: проприорецепторы, терморецепторы, обонятельные клетки.
Вторично-чувствующие рецепторы: в них раздражитель действует на рецепторную клетку, в ней возникает возбуждение (рецепторный потенциал). На мембране аксона рецепторный потенциал активирует выделение нейромедиатора в синапс, в результате чего на постсинаптической мембране второго нейрона (чаще всего биполярного) образуется генераторный потенциал, который и приводит к образованию потенциала действия на соседних участках постсинаптической мембраны. Далее этот потенциал действия передается в ЦНС. Примеры: волосковые клетки уха, вкусовые рецепторы, фоторецепторы глаза.
18. Основные принципы распространения возбуждения в ЦНС: конвергенция, дивергенция, мультипликация, иррадиация, реверберация, одностороннее проведение.
Конвергенция -схождение к одному нейрону двух или нескольких возбуждений от сенсорных раздражителей (например звук, свет).
Дивергенция - направляется в разные стороны - способность одиночного нейрона устанавливать многочисленные синаптические связи с различными нервными клетками. Иррадиация в физиологии, распространение процесса возбуждения или торможения в центральной нервной системе. Важную роль И. играет в деятельности коры больших полушарий головного мозга.
Реверберация -длительная циркуляция импульсов возбуждения в сетях нейронов или между различными структурами ц.н.с.
Одностороннее проведение возбуждения - возбуждение передается с афферентного на эфферентный нейрон. Причина: клапанное свойство синапса.
19. Торможение в ЦНС (И.М.Сеченов, Гольц, Мегун). Современные представления об основных видах центрального торможения : постсинаптического, пресинаптического и их механизмах.
Интегративная и координационная деятельность центральных нервных образований осуществляется при обязательном участии тормозных процессов. Торможение в ЦНС открыл И.М.Сеченов (1863). В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Было показано, что латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Открытие И.М.Сеченова послужило толчком для дальнейших исследований торможения в ЦНС. В частности, обнаружил проявления торможения у спинальной лягушки Ф.Гольц (1870). Он также исследовал латентное время рефлекса. При этом оказалось, что механическое раздражение кончиков пальцев одной конечности лягушки существенно удлиняет латентный период сгибательного рефлекса другой конечности при погружении ее в раствор кислоты.Наличие специальных тормозных структур в продолговатом мозге доказал Х.Мегун (1944). В опытах на кошках при изучении разгиба- тельного рефлекса Х.Мегун установил, что раздражение медиальной части ретикулярной формации продолговатого мозга тормозит рефлекторную активность спинного мозга.
Тонкий анализ тормозных явлений в ЦНС позволил выделить две разновидности торможения: 1) постсинаптическое торможение и 2) пресинаптическое торможение.
А.Этот вид торможения открыл Д.Экклс(1952) при регистрации потенциалов мотонейронов спинного мозга у кошки во время раздражения мышечных афферентов группы1а. При этом оказалось, что в мотонейронах мышцы-актагониста регистрируются не деполяризацияи возбуждение, а гиперполяризационный постсинаптический потенциал, уменьшающий возбудимость мотонейрона, угнетающий его способность реагировать на возбуждающие влияния. По этой причине вызванный гиперполяризационный потенциал был назван тормозным постсинаптическим потенциалом, ТПСП.У кошки ТПСП регистрируется на 0,5 мс позже, чем ВПСП, что объясняется наличием на пути проведения возбуждения, запускающего ТПСП, одного дополнительного синапса.
Б. Механизм постсинаптического торможения. Возбудимость клетки от ТПСП (гиперполяризационного постсинаптического потенциала)уменьшается, потому что увеличивается пороговый потенциал (AV), так как Екр.(критический уровень деполяризации, КУД)остается на прежнем уровне, а мембранный потенциал (Ео) возрастает. ТПСП возникает под влиянием и аминокислоты глицина,и ГАМК — гамма-аминомасляной кислоты. В спинном мозге глицин выделяется особыми тормозными клетками(клетки Реншоу) в синапсах, образуемых этими клетками на мембраненейрона-мишени. Действуя на ионотропный рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для СГ, при этом СП поступает в клетку согласно концентрационному градиенту вопреки электрическому градиенту, в результате чего развивается гиперполяризация. В бесхлорной среде тормозная роль глицина не реализуется.
Разновидности постсинаптического торможения. Возвратное постсинаптическое торможение— торможение, при котором тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют. Параллельное торможение может выполнять подобную же роль, когда возбуждение блокирует само себя, за счет дивергенции по коллатерали с включением тормозной клетки на своем пути и возвратом импульсов к нейрону, который активировался этим же возбуждением. Латеральное постсинаптическое торможение. Тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями.
А.Открытие. Пресинаптическое торможение первоначально выявлено также в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов. Так, известно, что раздражение первичных афферентов мышечных веретен сопровождается возбуждением гомонимных а-мотонейронов (а-мотонейронов этой же мышцы). Однако опережающее раздражение афферентов сухожильных рецепторов мышц-антагонистов предотвращает возбуждение активируемых а-мотонейронов. Мембранный потенциал и возбудимость исследуемых а-мотонейронов не изменялись либо регистрировался низкоамплитудный ВПСП, недостаточный для возникновения ПД. Мотонейроны не возбуждаются вследствие процессов, происходящих в пресинаптическом окончании, что определяет название этого вида торможения.
Б. Механизм пресинаптического торможения.Электрофизиологическое изучение процессов на уровне пресинаптических окончаний регистрируется выраженная и продолжительная деполяризация, что и ведет к развитию торможения. В очаге деполяризации нарушается процесс распространения возбуждения — следовательно, поступающие импульсы, не имея возможности пройти зонудеполяризации в обычном количестве иобычной амплитуде, не обеспечивают выделения медиатора в синаптическую щель в достаточном количестве, поскольку мало ионов Са2+ входит в нервное окончание — нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным. Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых образуют синапсы на пресинаптических окончаниях аксона- мишени. Торможение (деполяризация) после одного афферентного залпа продолжается 300—400 мс, медиатором является гамма-аминомасляная кислота (ГАМК), которая действует на ГАМК,-рецепторы. Деполяризация является следствием повышения проницаемости для СГ, в результате чего он выходит из клетки.
Читайте также:
