
Потенциал действия или покой нервного импульса
Работа органов и тканей нашего организма зависит от многих факторов. Некоторые клетки (кардиомиоциты и нервы) зависят от передачи нервных импульсов, генерируемых в специальных компонентах клеток или узлах. В основе нервного импульса лежит образование специфической волны возбуждения, носящей название потенциала действия.
Что это такое?
Потенциалом действия принято называть волну возбуждения, передвигающуюся от клетки к клетке. За счет ее образования и прохождения через клеточные мембраны происходит кратковременное изменение их заряда (в норме внутренняя сторона мембраны заряжена отрицательно, а наружная – положительно). Образованная волна способствует изменению свойств ионных каналов клетки, что приводит к перезарядке мембраны. В тот момент, когда потенциал действия проходит через мембрану, происходит кратковременное изменение ее заряда, что приводит к изменению свойств клетки.
Образование данной волны лежит в основе функционирования нервного волокна, а также системы путей проведения сердца.
При нарушении его образования развиваются многие заболевания, что делает определение потенциала действия необходимым в комплексе лечебно-диагностических мероприятий.
Как же образуется потенциал действия и что для него характерно?
История исследования
Изучение возникновения возбуждения в клетках и волокнах было начато довольно давно. Первыми его возникновение заметили биологи, изучавшие воздействие различных раздражителей на оголенный берцовый нерв лягушки. Ими было замечено, что при воздействии на него концентрированным раствором пищевой соли наблюдалось сокращение мышц.
В дальнейшем исследования были продолжены неврологами, однако основная наука после физики, изучающая потенциал действия – физиология. Именно физиологами было доказано наличие потенциала действия в клетках сердца и нервах.
По мере углубления в изучение потенциалов было доказано наличие и потенциала покоя.
С начала 19 века начали создаваться методы, позволяющие зафиксировать наличие данных потенциалов и измерить их величину. В настоящее время фиксация и изучение потенциалов действия проводится в двух инструментальных исследованиях – снятии электрокардиограмм и электроэнцефалограмм.
Механизм потенциала действия
Образование возбуждения происходит за счет изменения внутриклеточной концентрации ионов натрия и калия. В норме в клетке содержится больше калия, чем натрия. Внеклеточная концентрация ионов натрия значительно больше, чем в цитоплазме. Изменения, вызываемые потенциалом действия, способствуют изменению заряда на мембране, в результате чего обуславливается ток ионов натрия внутрь клетки. Из-за этого изменяются заряды снаружи и внутри клетки (цитоплазма заряжается положительно, а внешняя среда – отрицательно.

Это делается для облегчения прохождения волны по клетке.
После того как волна была передана через синапс, происходит обратное восстановление заряда за счет тока внутрь клетки отрицательно заряженных ионов хлора. Восстанавливаются исходные уровни заряда снаружи и внутри клетки, что приводит к образованию потенциала покоя.
Периоды покоя и возбуждения чередуются. В патологической клетке все может происходить иначе, и образование ПД там будет подчиняться несколько иным законам.
Фазы ПД
Течение потенциала действия можно разделить на несколько фаз.
Первая фаза протекает до образования критического уровня деполяризации (проходящим потенциалом действия стимулируется медленная разрядка мембраны, которая достигает максимального уровня, обычно он составляет около -90 мЭв). Данная фаза носит название предспайк. Осуществляется за счет входа в клетку ионов натрия.
Следующая фаза - пиковый потенциал (или спайк), образует параболу с острым углом, где восходящая часть потенциала означает деполяризацию мембраны (быстрая), а нисходящая часть – реполяризацию.
Третья фаза – отрицательный следовый потенциал – показывает следовую деполяризацию (переход от пика деполяризации до состояния покоя). Обусловлена входом ионов хлора внутрь клетки.
На четвертом этапе, фазе положительного следового потенциала, происходит возврат уровней заряда мембраны к исходному.
Данные фазы, обусловленные потенциалом действия, строго следуют одна за одной.
Функции потенциала действия
Несомненно, развитие потенциала действия имеет важное значение в функционировании тех или иных клеток. В работе сердца возбуждению принадлежит главная роль. Без него сердце было бы просто неактивным органом, но за счет распространения волны по всем клеткам сердца происходит его сокращение, что способствует проталкиванию крови по сосудистому руслу, обогащению ею всех тканей и органов.
Нервная система также не могла бы нормально выполнять свою функцию без потенциала действия. Органы не могли бы получать сигналы к выполнению той или иной функции, в результате чего были бы просто бесполезными. Кроме того, совершенствование передачи нервного импульса в нервных волокнах (появление миелина и перехватов Ранвье) позволило передавать сигнал за считаные доли секунды, что и обусловило развитие рефлексов и сознательных движений.
Кроме данных систем органов, потенциал действия образуется и во многих других клетках, однако в них он играет роль лишь в выполнении клеткой своих специфических функций.
Возникновение потенциала действия в сердце
Основным органом, работа которого основана на принципе образования потенциала действия, является сердце. За счет существования узлов образования импульсов осуществляется работа данного органа, функция которого заключается в доставке крови к тканям и органам.
Генерация потенциала действия в сердце происходит в синусовом узле. Он находится в месте впадения полых вен в правом предсердии. Оттуда импульс распространяется по волокнам проводящей системы сердца – от узла к атриовентрикулярному соединению. Проходя по пучку Гиса, точнее, по его ножкам, импульс проходит к правому и левому желудочку. В их толще расположены более мелкие пути проведения – волокна Пуркинье, по которым возбуждение доходит до каждой клетки сердца.
Потенциал действия кардиомиоцитов является составным, т.е. зависит от сокращения всех клеток сердечной ткани. При наличии блока (рубец после инфаркта) образование потенциала действия нарушается, что фиксируется на электрокардиограмме.
Нервная система
Как же образуется ПД в нейронах – клетках нервной системы. Тут все осуществляется несколько проще.
Внешний импульс воспринимается отростками нервных клеток – дендритами, связанными с рецепторами, расположенными как в коже, так и во всех других тканях (потенциал покоя и потенциал действия также сменяют друг друга). Раздражение провоцирует образование потенциала действия в них, после чего импульс через тело нервной клетки идет в ее длинный отросток – аксон, а от него через синапсы – к другим клеткам. Таким образом, образованная волна возбуждения доходит до головного мозга.
Особенностью нервной системы является наличие двух типов волокон – покрытых миелином и без него. Возникновение потенциала действия и его передача в тех волокнах, где есть миелин, осуществляется значительно быстрее, чем в демиелинезированных.
Данный феномен наблюдается из-за того, что распространение ПД по миелинизированным волокнам происходит за счет “прыжков” – импульс перескакивает участки миелина, что в результате уменьшает его путь и, соответственно, ускоряет распространение.
Потенциал покоя
Без развития потенциала покоя не было бы и потенциала действия. Под потенциалом покоя понимают нормальное, невозбужденное состояние клетки, при котором заряды внутри и вне ее мембраны значительно отличаются (то есть снаружи мембрана заряжена положительно, а внутри – отрицательно). Потенциал покоя показывает разницу между зарядами внутри и извне клетки. Обычно в норме он составляет от -50 до -110 мЭв. В нервных волокнах данная величина обычно равна -70 мЭв.
Обусловлен он миграцией ионов хлора внутрь клетки и созданием негативного заряда на внутренней стороне мембраны.

При смене концентрации внутриклеточных ионов (как было указано выше) ПП сменяет ПД.
В норме все клетки организма находятся в невозбужденном состоянии, поэтому смену потенциалов можно считать физиологически необходимым процессом, так как без них не могли бы осуществлять свою деятельность сердечно-сосудистая и нервная системы.
Значимость исследования потенциалов покоя и действия
Потенциал покоя и потенциал действия позволяют определить состояние организма, а также отдельных органов.
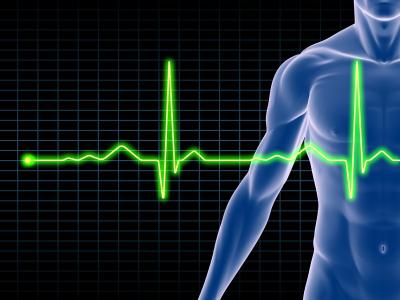
Фиксация потенциала действия с сердца (электрокардиография) позволяет определить его состояние, а также функциональную способность всех его отделов. Если изучать нормальную ЭКГ, то можно заметить, что все зубцы на ней есть проявление потенциала действия и последующего потенциала покоя (соответственно, возникновение данных потенциалов в предсердиях отображает зубец Р, а распространение возбуждения в желудочках – зубец R).
Что касаемо электроэнцефалограммы, то на ней возникновение различных волн и ритмов (в частности, альфа и бета-волн у здорового человека) также обусловлено возникновением потенциалов действия в нейронах головного мозга.
Данные исследования позволяют своевременно выявить развитие того или иного патологического процесса и обуславливают практически до 50 процентов успешного лечения исходного заболевания.
При образовании потенциалов действия (спайное) в химических синапсах между нейронами происходит высвобождение медиаторов из пузырьков пресинаптической мембраны. Медиатор проходит через синаптическую щель и активирует рецепторы, встроенные в постсинаптическую мембрану нейрона-мишени. Рецепторы, в свою очередь, активируют медиаторозависимые ионные каналы, изменяющие поляризацию нейрона-мишени. При смещении показателей мембранного потенциала от уровня потенциала покоя (-70 мВ) в отрицательную сторону (например, — 80 мВ и более) происходит гиперполяризация мембраны, а при смещении в положительную сторону — деполяризация.
а) Электротонические потенциалы. Первоначальный ответ нейрона-мишени на воздействие возбуждающих импульсов заключается в формировании локальных ступенчатых электротонических потенциалов. Положительные злектротонические потенциалы обычно возникают в мультииолярных нейронах в результате деполяризации, обусловленной активацией медиаторозависимых ионных каналов. При низкочастотном стимулирующем воздействии по дендритам распространяются небольшие волны деполяризации, угасающие в течение 2-3 мс.
При увеличении частоты воздействия происходит ступенчатая временная суммация возбуждения, в результате чего образуется прогрессивно увеличивающаяся волна, которая распространяется на поверхность сомы. Одновременное схождение в области сомы волн возбуждения, распространяющихся по разным дендритам, приводит к пространственной суммации возбуждения. При деполяризации приблизительно на 15 мВ (и значении потенциала -55 мВ) происходит открытие потенциалозависимых ионных каналов в наиболее чувствительной области нейрона — триггерной точке (триггерной области), располагающейся в начальном сегменте аксона. При достижении уровня деполяризации (генераторного потенциала) пороговых значений, т.е. потенциала, способного запустить открытие потенциалозависимых ионных каналов, происходит формирование потенциала действия.
В триггерной зоне чувствительных нейронов черешнях и спинномозговых нервов формируется так называемый рецепторный потенциал. В этой области расположено большое количество ионных каналов, активирующихся под воздействием специфических чувствительных стимулов и вызывающих ступенчатое распространение волны деполяризации.
Определение триггерной области миелинизированных нервных волокон не вызывает затруднений: у мультиполярных нейронов эта область граничит с первым миелиновым сегментом с его проксимальной стороны, а у периферических чувствительных нейронов — с последним миелиновым сегментом с его дистальной стороны.
Тормозные (гиперполяризующие) постсинаптические потенциалы возникают при открытии ионных каналов, генерирующих выходящий ток (например, калиевых каналов). Распространение тормозных потенциалов также является угасающим.
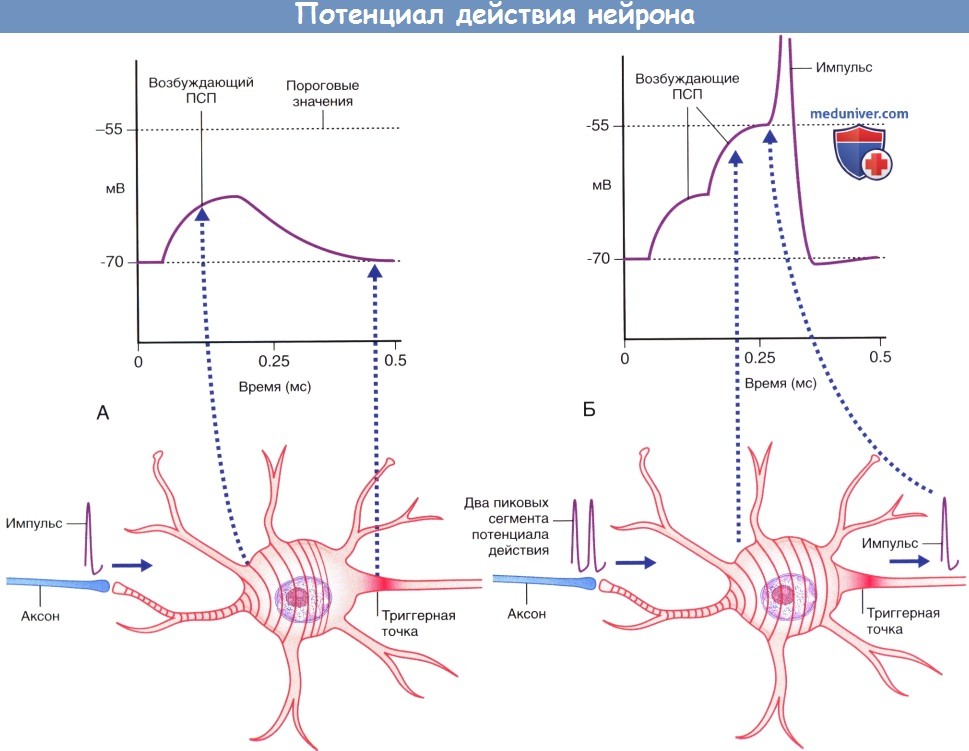
Временная суммация.
(А) Аксон чувствительного нейрона (выделен голубым цветом) передает единичный импульс двигательному нейрону,
вызывающий возбуждающему постсинаптический потенциал (ПСП), значения которого не достигают пороговых величин, вследствие чего этот потенциал затухает.
(Б) Аксон чувствительного нейрона передает два импульса.
Происходит временная суммация импульсов с достижением в начальном сегменте аксона пороговых значений возбуждения,
что приводит к формированию потенциала действия, который в дальнейшем распространяется по аксону двигательного нейрона. 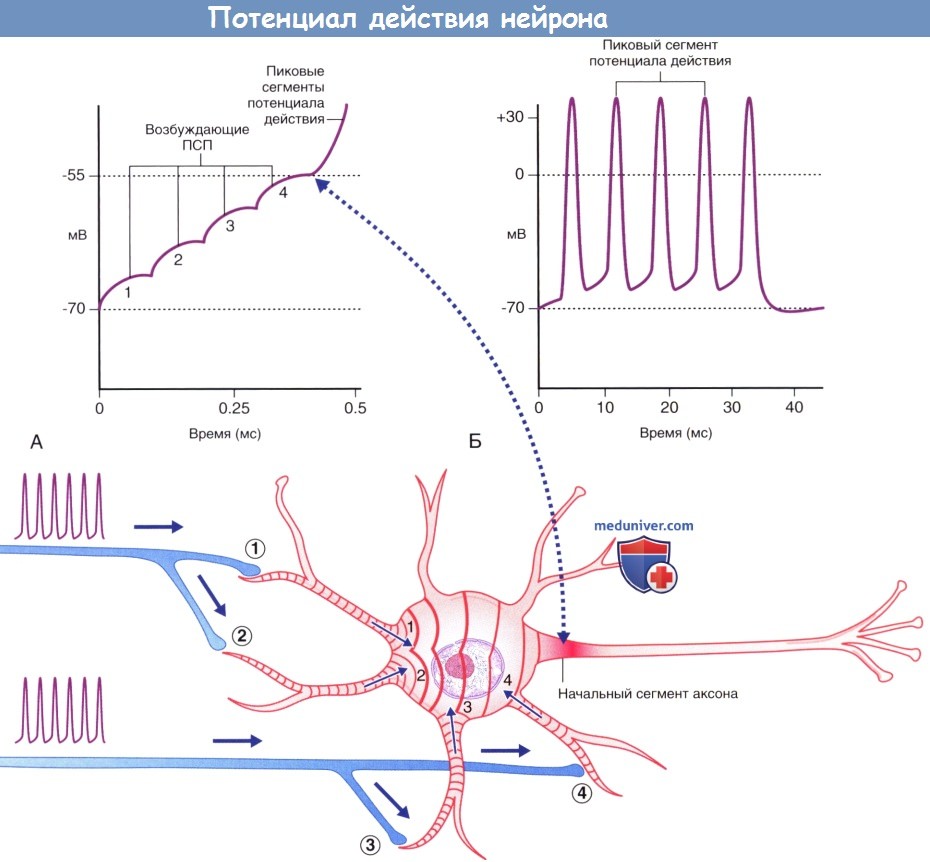
(А) Ступенчатая суммация возбуждающих постсинаптических потенциалов (ПСП), за счет которой происходит формирование пикового потенциала. Пунктирной линией указана область, соответствующая графику А.
(Б) Генераторные потенциалы достаточной силы формируют множественные пиковые потенциалы. 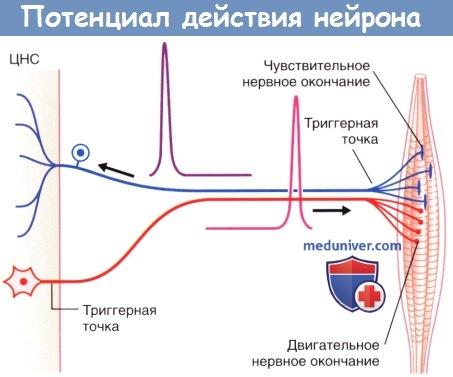
Потенциалы действия двигательных и чувствительных нейронов скелетной мускулатуры. ЦНС — центральная нервная система. 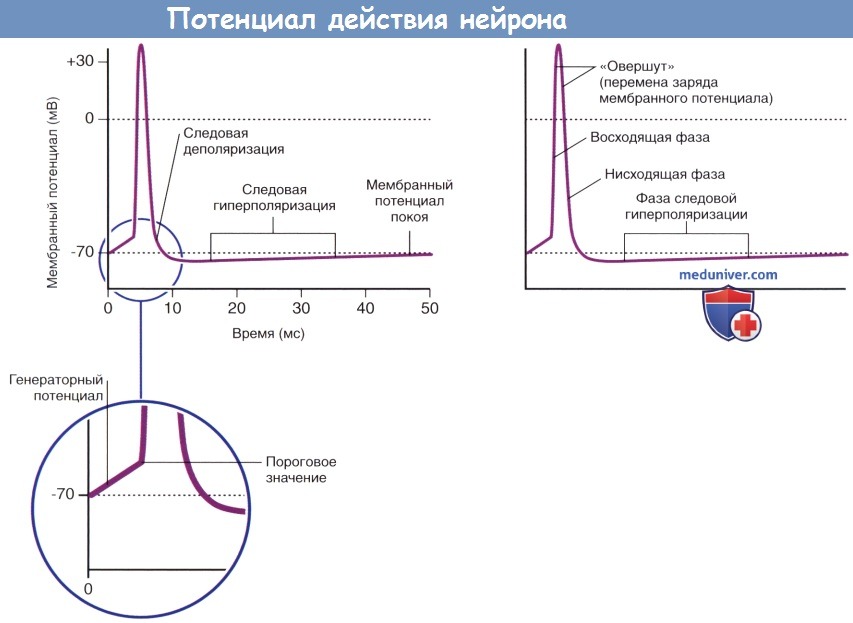
Основные характеристики потенциала действия.
Возникновение фазы следовой гиперполяризации объясняют тем, что после полной инактивации потенциалозависимых натриевых каналов многие калиевые каналы остаются открытыми. Восстановление ионных градиентов обеспечивает натрий-калиевый насос.
В ходе подробного изучения натриевых каналов было обнаружено, что их регуляция осуществляется двумя путями. В состоянии покоя (-70 мВ) активационные ворота, расположенные в средней части натриевых и калиевых каналов, закрыты. Первыми на пороговое раздражение реагируют натриевые каналы: происходят открытие активационных ворот и быстрый выход ионов Na + в клетку по концентрационному и электрическому градиентам. При достижении потенциалом действия пиковых значений (+35 мВ) происходит открытие калиевых каналов, а натриевые каналы закрываются инактивационными воротами, представленными глобулярным белком. При реполяризации и восстановлении мембранного потенциала (-70 мВ) активационные натриевые ворота закрываются, а затем вновь открываются инактивационные натриевые ворота, тем самым приводя натриевые каналы в исходное состояние.
Работа потенциалозависимых калиевых каналов регулирует одноворотный механизм: этими каналами управляют только активационные ворота.
Во время фазы нарастания и в начале фазы реполяризации потенциала действия нейрон находится в абсолютно рефрактерном состоянии и не может воспринимать следующие импульсы в результате инактивации потенциалозависимых натриевых каналов. Вслед за этим периодом наступает интервал относительной рефрактерности, во время которого стимул, превышающий исходный более чем на 15 мВ, способен вызвать формирование повторного потенциала действия. Часто для достижения показателя 35 мВ генераторный потенциал производит от 50 до 100 импульсов в секунду (частота составляет от 50 до 100 Гц).
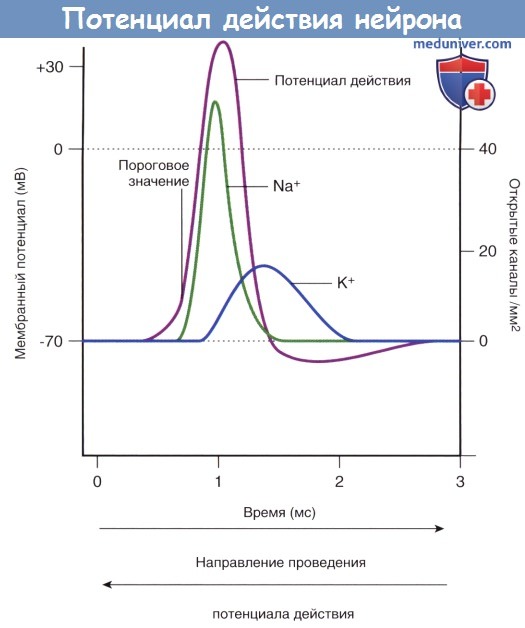
Изменения проницаемости потенциалозависимых натриевых и калиевых каналов, обеспечивающих развитие потенциала действия. 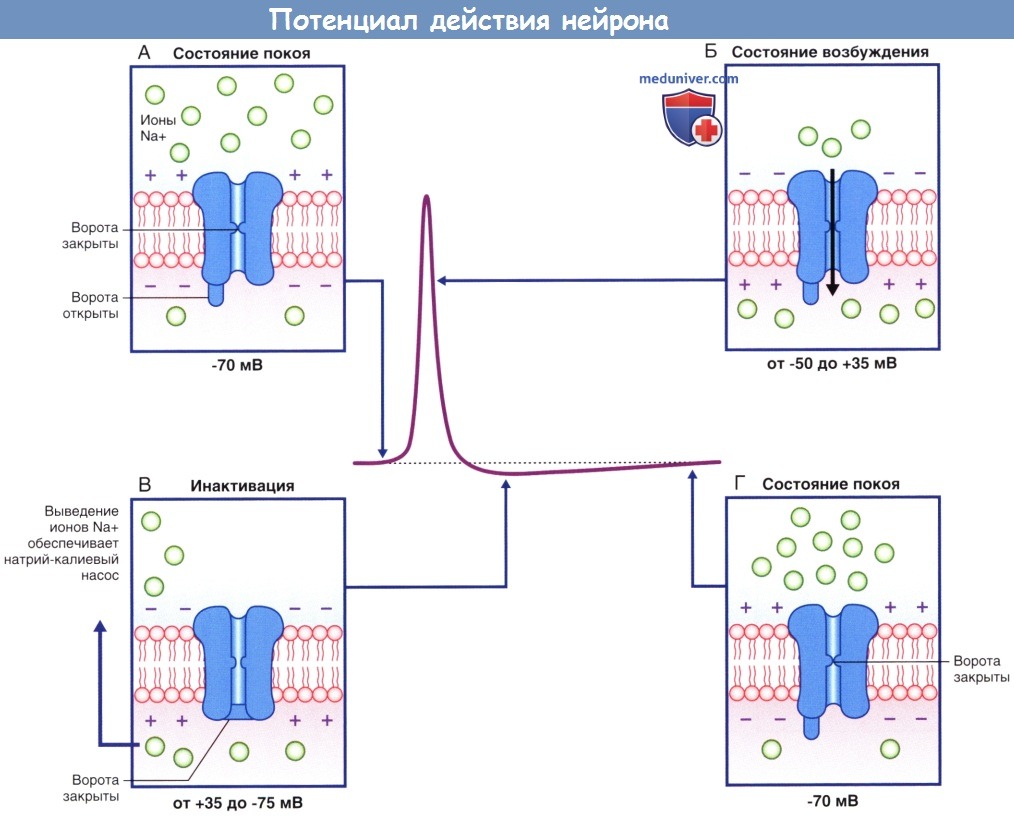
Потенциалозависимые натриевые каналы в разные фазы потенциала действия.
(А) В состоянии покоя, предшествующем возбуждению, активационные ворота находятся в закрытом состоянии, а инактивационные — в открытом.
(Б) При преодолении порога возбуждения происходит открытие активационных ворот, что приводит к полному открытию ионного канала.
(В) Инактивационные ворота закрываются, что приводит к закрытию ионного канала.
(Г) При восстановлении потенциала покоя активационные ворота закрываются, а инактивационные — открываются.
Ионный канал возвращается в исходное состояние. 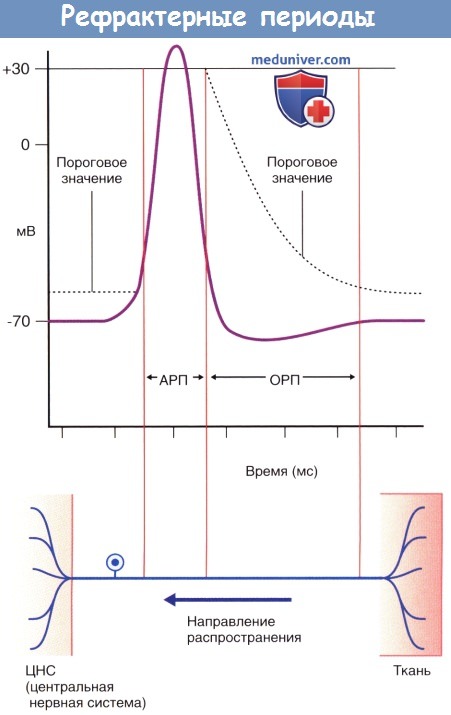
Рефрактерные периоды.
АРП — абсолютный рефрактерный период;
ОРП — относительный рефрактерный период.
в) Передача нервных импульсов. Изменение потенциала триггерной зоны распространяется вдоль аксона в соответствии с электротонической траекторией. Положительные заряды внутренней стороны мембраны распространяются в обоих направлениях через аксоплазму. Положительные заряды внешней стороны мембраны аналогичным образом проходят через внеклеточную жидкость и нейтрализуют отрицательный заряд на внешней стороне мембраны. Прилежащий проксимальный участок мембраны достаточно рефрактерен и способен противостоять деполяризации. Одновременно с этим в прилежащем дистальном сегменте возникает локальный ответ (деполяризация), который достигает пороговых величин и распространяется далее вдоль ствола и ветвей аксона к нервным окончаниям, обеспечивая проведение потенциала действия.
Проведение импульса по немиелинизированному нервному волокну происходит непрерывно, а по миелинизированному — сальтаторно (скачкообразно). Миелиновая оболочка выполняет функцию изолятора и располагается между перехватами Ранвье, где сконцентрированы натриевые каналы. В связи с этим в каждом перехвате Ранвье последовательно возникают потенциалы действия за счет распространения положительного тока вдоль аксоплазмы миелинизированного участка аксона и его выхода в области следующего перехвата Ранвье. При распространении тока в обратном направлении по внеклеточной жидкости для повторного заряда деполяризованного участка мембраны снятие положительного заряда приводит к деполяризации следующего перехвата Ранвье.
г) Скорость проведения нервных импульсов. Скорость проведения нервного импульса по немиелинизированным нервным волокнам прямо пропорциональна диаметру аксона. В первую очередь это связано с тем, что больший объем аксоплазмы обеспечивает более высокую скорость продольного распространения возбуждения. Кроме того, чем больше диаметр аксона, тем больше площадь поверхности его плазматической мембраны, следовательно, на ней расположено большее количество ионных каналов, а деполяризация и восстановление заряда мембраны происходят быстрее. Диаметр аксонов варьирует от 0,2 до 2 мкм, а скорость проведения импульсов — от 2 до 25 м/с.
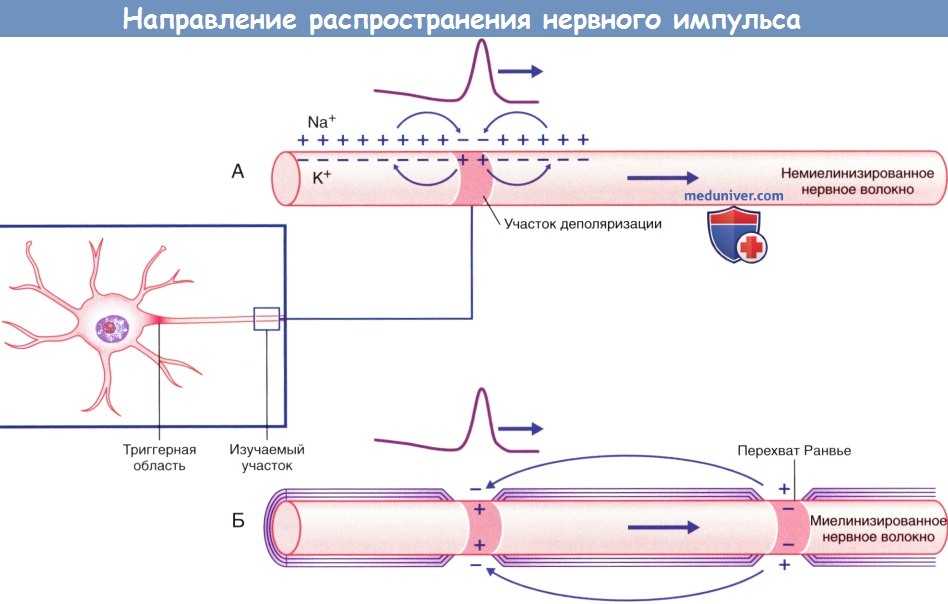
Направление распространения импульса, показанное в виде прохождения положительного заряда.
(А) Непрерывное проведение импульса по немиелинизированному нервному волокну.
(Б) Скачкообразное проведение импульса по миелинизированному нервному волокну.
д) Механизм действия местных анестетиков. При применении местных анестетиков происходит обратимая блокировка проведения нервных импульсов за счет инактивации натриевых каналов, особенно потенциалозависимых, вследствие чего деполяризация мембраны становится невозможной. Молекулы местных анестетиков связывают белок, окружающий ионные каналы. В течение времени действия анестетика происходит связывание белка, окружающего ионные каналы. Для доступа к этим белкам местные анестетики должны быть хорошо растворимы в жирах. При введении анестетика вблизи периферического нерва в первую очередь происходит инактивация немиелинизированных и тонко миелинизированных Аб-волокон. При воздействии на смешанный нерв может возникнуть временный двигательный паралич.
Большинство местных анестетиков — производные амидов (бупивакаин, лидокаин) или сложных эфиров (бензокаин, новокаин). И те, и другие вызывают местное расширение сосудов за счет прямого расслабления гладкой мускулатуры артериол, что приводит к ускорению их выведения. В связи с этим в растворы для местной анестезии часто добавляют адреналин, вызывающий сокращение гладкой мускулатуры и сужение сосудов.
е) Резюме. Электрически заряженные атомы или группы атомов называют ионами. Ионы Na + и К + являются катионами, а ионы Cl + и отрицательно заряженные белки — анионами. Клеточные мембраны представляют собой заряженные конденсаторы с потенциалом покоя величиной -70 мВ.
Неуправляемые натриевые, калиевые и хлорные каналы открыты постоянно, и движение соответствующих ионов через них происходит по градиенту концентрации. Натриевые каналы относительно малочисленны, а калиевые и кальциевые каналы представлены в большом количестве. Ионы К + сконцентрированы в цитозоле за счет притягивающего влияния белков-анионов цитоскелета и отталкивающего действия ионов Na + , расположенных на внешней части плазматической мембраны. Натрий-калиевый насос обеспечивает поддержание мембранного потенциала.
Первоначальная реакция мультиполярного нейрона на возбуждающий стимул заключается в формировании угасающих волн положительного электротонуса. В результате их временной и/или пространственной суммации происходит образование генераторного потенциала в начальном сегменте аксона. При достижении пороговых значений (-55 мВ) потенциалозависимые ионные каналы генерируют потенциал действия, распространяющийся по нервному волокну. Тормозные стимулы вызывают формирование волн отрицательного электротонуса, суммация которых приводит к гиперполяризации мембраны, тем самым отдаляя величины ее потенциала от пороговых значений.
Потенциал действия (спайк) проходит фазу нарастания (деполяризации) от начальных значений до +35 мВ, фазу убывания (реполяризации), в которую возвращается к исходным показателям, и фазу следовой гиперполяризации, в которую понижается до -75 мВ, а затем вновь возвращается к первоначальным значениям. Деполяризация мембраны происходит в результате активации потенциалозависимых натриевых каналов, в ходе которой происходит их стремительное (со скоростью + в клетку и деполяризации мембранного потенциала. При достижении мембранного потенциала +35 мВ происходит закрытие натриевых каналов при помощи инактивационных ворот.
При достижении потенциала действия максимальных значений происходит открытие потенциалозависимых калиевых каналов, и поток ионов Na + в клетку сменяется потоком ионов К + из клетки, что приводит к реполяризации и следовой гиперполяризации мембранного потенциала.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Нервный импульс
Строго говоря, движущееся по нервам возбуждение представляет собой нервные импульсы, а не потенциалы действия.
Можно сказать короче:
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин "потенциал действия". Хотя потенциал действия - это только электрический компонент нервного импульса.
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
Потенциал действия - это электрическая характеристика (электрическая составляющая) нервного импульса.
Нервный импульс - это сложный структурно-электро-химический процесс, распространяющийся по мембране нейрона в виде бегущей волны изменений.
Потенциал действия - это только электрический компонент нервного импульса, характеризующий изменения электрического заряда (потенциала) на локальном участке мембраны во время прохождения через него нервного импульса (от -70 до +30 мВ и обратно). (Кликните на изображение слева, чтобы увидеть анимацию.)
Сравните два приведённых выше рисунка (покликайте по ним) и, как говорится, почувствуйте разницу!
Где рождаются нервные импульсы?
Как ни странно, не все студенты, изучившие физиологию возбуждения, могут ответить на этот вопрос. ((
Хотя ответ не сложен. Нервные импульсы рождаются на нейронах всего в нескольких местах:
1) аксонный холмик (это переход тела нейрона в аксон),
2) рецепторное окончание дендрита,
3) первый перехват Ранвье на дендрите (триггерная зона дендрита),
4) постсинаптическая мембрана возбуждающего синапса.
Места возникновения нервных импульсов:
1. Аксонный холмик - главный породитель нервных импульсов.
Аксонный холмик - это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем (генератором) нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интегрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) - и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим "начинаниям". Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они "перебиваются" потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона.
Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала генераторный потенциал, а затем и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита).
Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия - и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса.
В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Видео: Проведение нервного импульса по нервному волокну
Потенциал действия как нервный импульс
Ниже размещён материал, взятый из учебно-методического пособия автора данного сайта, на который вполне можно ссылаться в своём списке литературы:
Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
Все процессы мембранных изменений, происходящих в ходе распространяющегося возбуждения, достаточно хорошо изучены и описаны в научной и учебной литературе. Но не всегда это описание легко понять, поскольку в данном процессе задействовано слишком много компонентов (с точки зрения обычного студента, а не вундеркинда, конечно).
Для облегчения понимания мы предлагаем рассматривать единый электрохимический процесс распространяющегося динамичного возбуждения с трех сторон, на трех уровнях:
Электрические явления – развитие потенциала действия.
Химические явления – движение ионных потоков.
Структурные явления – поведение ионных каналов.
Три стороны процесса распространяющегося возбуждения
1. Потенциал действия (ПД)
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Обычно мембранный потенциал в нейронах ЦНС изменяется от –70 мВ до +30 мВ, а затем вновь возвращается к исходному состоянию, т.е. к –70 мВ. Как видим, понятие потенциала действия характеризуется через электрические явления на мембране.
На электрическом уровне изменения начинаются как смена поляризованного состояния мембраны на деполяризацию. Сначала деполяризация идет в виде локального возбуждающего потенциала. Вплоть до критического уровня деполяризации (примерно –50 мВ) это относительно простое линейное уменьшение электроотрицательности, пропорциональное силе воздействующего раздражителя. А вот потом начинается более крутая самоусиливающаяся деполяризация, она развивается не с постоянной скоростью, а с ускорением . Говоря образно, деполяризация так разгоняется, что перескакивает через нулевую отметку, не заметив этого, и даже переходит в положительную поляризацию. После достижения пика (обычно +30 мВ) начинается обратный процесс – реполяризация , т.е. восстановление отрицательной поляризации мембраны.
Кратко опишем электрические явления во время течения потенциала действия:
Восходящая ветвь графика:
потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ);
нарастающий локальный потенциал – пропорциональная раздражителю деполяризация;
критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации (за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия;
самоусиливающаяся круто нарастающая деполяризация;
переход нулевой отметки (0 мВ) – смена полярности мембраны;
пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия.
Нисходящая ветвь графика:
реполяризация – восстановление прежней электроотрицательности мембраны;
переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную;
переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости;
следовые процессы (следовая деполяризация или следовая гиперполяризация);
восстановление потенциала покоя – норма (–70 мВ).
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
2. Ионные потоки
Итак, важно осознать, что все электрические токи, которые идут через мембрану, являются ионными потоками . Привычного нам из физики тока в виде потока электронов в клетках, как в водных системах, просто нет. Ссылки на потоки электронов будут ошибкой.
На химическом уровне мы, описывая распространяющееся возбуждение, должны рассмотреть, как изменяются характеристики ионных потоков, идущих через мембрану. Главное в этом процессе то, что при деполяризации резко усиливается поток ионов натрия внутрь клетки, а затем он внезапно прекращается на спайке потенциала действия. Входящий поток натрия как раз и вызывает деполяризацию, так как ионы натрия приносят с собой положительные заряды в клетку (чем и снижают электроотрицательность). Затем, после спайка, значительно нарастает выходящий наружу поток ионов калия, что вызывает реполяризацию. Ведь калий, как мы неоднократно говорили, выносит с собой из клетки положительные заряды. Отрицательные заряды остаются внутри клетки в большинстве, и за счет этого усиливается электроотрицательность. Это и есть восстановление поляризации за счет выходящего потока ионов калия. Заметим, что выходящий поток ионов калия возникает практически одновременно с появлением натриевого потока, но нарастает медленно и длится в 10 раз дольше. Несмотря на продолжительность калиевого потока самих ионов расходуется немного – всего одна миллионная доля от запаса калия в клетке (0,000001 часть).
Подведем итоги. Восходящая ветвь графика потенциала действия образуется за счет входа в клетку ионов натрия, а нисходящая – за счет выхода из клетки ионов калия.
3. Ионные каналы
Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы .
Пожар как метафора распространяющегося возбуждения
Когда мы искали образный пример, аналогию или метафору, которая может наглядно передать смысл распространяющегося возбуждения, то остановились на образе пожара. Действительно, распространяющееся возбуждение похоже на лесной пожар, когда горящие деревья остаются на месте, а фронт огня распространяется и уходит все дальше и дальше во все стороны от очага возгорания.
Как же в этой метафоре будет выглядеть явление торможения?
Ответ очевиден – торможение будет выглядеть как тушение пожара, как уменьшение горения и затухание огня. Но если огонь распространяется сам по себе, то тушение требует усилий. Из потушенного участка процесс тушения сам по себе не пойдет во все стороны.
Существует три варианта борьбы с пожаром: (1) либо надо ждать, когда все сгорит и огонь истощит все горючие запасы, (2) либо надо поливать водой горящие участки, чтобы они погасли, (3) либо надо поливать заранее ближайшие нетронутые огнем участки, чтобы они не загорелись.
АВТОВОЛНЫ В АКТИВНО-ВОЗБУДИМЫХ СРЕДАХ (АВС)
При распространении волны в активно-возбудимых средах не происходит переноса энергии. Энергия не переносится, а освобождается, когда до участка АВС доходит возбуждение. Можно провести аналогию с серией взрывов зарядов, заложенных на некотором расстоянии друг от друга (например, при тушении лесных пожаров, строительстве, мелиоративных работах), когда взрыв одного заряда вызывает взрыв рядом расположенного и так далее. Лесной пожар также является примером распространения волны в активно- возбудимой среде. Пламя распространяется по области с распределенными запасами энергии - деревья, валежник, сухой мох.
Основные свойства волн, распространяющихся в активно-возбудимых средах (АВС)
Волна возбуждения распространяется в АВС без затухания; прохождение волны возбуждения связано с рефрактерностью - невозбудимостью среды в течение некоторого промежутка времени (периода рефрактерности).
Видео: Потенциал действия (Action potential)
Читайте также:
