
Проведення збудження по нервам
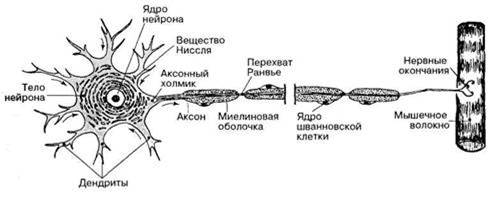
Функцию быстрой передачи возбуждения к нервной клетке и от нее выполняют ее отростки - дендриты и аксоны, т.е. нервные волокна. В зависимости от структуры их делят на мякотные, имеющие миелиновую оболочку, и безмякотные. Эта оболочка формируется шванновскими клетками, являющиеся видоизмененными глиальными клетками. Они содержат миелин, который в основном состоит из липидов. Он выполняет изолирующую и трофическую функции. Одна шванновская клетка образует оболочку на 1 мм нервного волокна. Участки, где оболочка прерывается, т.е. не покрыты миелином называют перехватами Ранвье. Ширина перехвата 1 мкм (рис.).
Функционально все нервные волокна делят на три группы:
1.Волокна типа А - это толстые волокна, имеющие миелиновую оболочку. В эту группу входят 4 подтипа:
1.1.Аa - к ним относятся двигательные волокна скелетных мышц и афферентные нервы, идущие от мышечных веретен (рецепторов растяжения). Скорость проведения по ним максимальна - 70-120 м/сек
1.2.Аb - афферентные волокна, идущие от рецепторов давления и прикосновения кожи. 30 - 70 м/сек
1.3.Аg - эфферентные волокна, идущие к мышечным веретенам (15 - 30 м/сек).
1.4.Аd - афферентные волокна от температурных и болевых рецепторов кожи (12 - 30 м/сек).
2. Волокна группы В - тонкие миелинизированные волокна, являющиеся преганглионарными волокнами вегетативных эфферентных путей. Скорость проведения - 3-18 м/сек
3.Волокна группы С, безмиелиновые постганглионарные волокна вегетативной нервной системы. Скорость 0,5 -3 м/сек.
Проведение возбуждения по нервам подчиняется следующим законам:
1.Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезке, вторая - действии веществ блокирующих проведение, например новокаина.
2.Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаще всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения.
3.Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола.
4.Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются проходя по ним.
5.Скорость проведения прямо пропорциональна диаметру нерва.
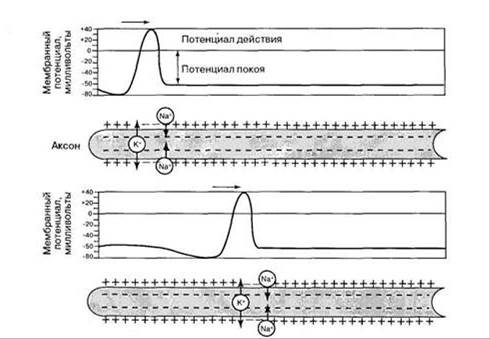
Нервные волокна обладают свойствами электрического кабеля, у которого не очень хорошая изоляция. В основе механизма проведения возбуждения лежит возникновение местных токов. В результате генерации ПД в аксонном холмике и реверсии мембранного потенциала, мембрана аксона приобретает противоположный заряд. Снаружи она становится отрицательной, внутри положительной. Мембрана нижележащего, невозбужденного участка аксона заряжена противоположным образом. Поэтому между этими участками, по наружной и внутренней поверхностям мембраны начинают проходить местные токи. Эти токи деполяризуют мембрану нижележащего невозбужденного участка нерва до критического уровня и в нем также генерируется ПД. Затем процесс повторяется и возбуждается более отдаленный участок нерва и т.д. (рис.). Т.к. по мембране безмякотного волокна местные токи текут не прерываясь, поэтому такое проведение называется непрерывным. При непрерывном проведении местные токи захватывают большую поверхность волокна, поэтому им требуется. длительное время для прохождения по участку волокна. В результате дальность и скорость проведения возбуждения по безмякотным волокнам небольшая.
В мякотных волокнах, участки покрытые миелином обладают большим электрическим сопротивлением. Поэтому непрерывное проведение ПД невозможно. При генерации ПД местные токи текут лишь между соседними перехватами. По закону "все или ничего" возбуждается ближайший к аксонному холмику перехват Ранвье, затем соседний нижележащий перехват и т.д. (рис.). Такое проведение называется сальтаторным (прыжком). При этом механизме ослабления местных токов не происходит и нервные импульсы распространяются на большое расстояние и с большой скоростью.
| | | следующая лекция ==> | |
| Физиология гладких мышц | | | Синаптическая передача |
Дата добавления: 2017-11-04 ; просмотров: 541 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью.
Проведение возбуждения по нервным волокнам осуществляется по определенным законам.
Закон двустороннего проведения возбуждения по нервному волокну.Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу.
Закон анатомической ифизиологической целостности нервного волокна.Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности). Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированногопроведения возбуждения по нервномуволокну.В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно.
Возбуждение (потенциал действия) распространяется по нервному волокну без затухания.
Периферический нерв практически неутомляем.
Механизм проведения возбуждения по нерву.
Возбуждение (потенциал действия - ПД) распространяется в аксонах, телах нервных клеток, а также иногда в дендритах без снижения амплитуды и без снижения скорости (бездекрементно). Механизм распространения возбуждения у различных нервных волокон неодинаков. При распространении возбуждения по безмиелиновому нервному волокну механизм проведения включает два компонента: раздражающее действие катэлектротона, порождаемое локальным ПД, на соседний участок электровозбудимой мембраны и возникновение ПД в этом участке мембраны. Локальная деполяризация мембраны нарушает электрическую стабильность мембраны, различная величина поляризации мембраны в смежных ее участках порождает электродвижущую силу и местный электрический ток, силовые линии которого замыкаются через ионные каналы. Активация ионного канала повышает натриевую проводимость, после электротонического достижения критического уровня деполяризации (КУД) в новом участке мембраны генерируется ПД. В свою очередь этот потенциал действия вызывает местные токи, а они в новом участке мембраны генерируют потенциал действия. На всем протяжении нервного волокна происходит процесс новой генерации потенциала действия мембраны волокна. Данный тип передачи возбуждения называется непрерывным.
Функцію швидкої передачі збудження в нервовій клітині і від неї виконують її відростки – дендрити і аксони, тобто нервові волокна. В залежності від структури їх ділять на м’якушеві, що мають мієлінових оболонку, і безмякотние. Ця оболонка формується шванновскими клітинами, які є видозміненими гліальними клітинами. Вони містять мієлін, який в основному складається з ліпідів. Він виконує ізолюючу і трофічну функції. Одна шванівську клітка утворить оболонку на 1 мм нервового волокна. Ділянки, де оболонка переривається, тобто Не покрита мієліном, називають перехопленнями Ранвье. Ширина перехоплення 1 мкм.
Функціонально всі нервові волокна ділять на три групи:
1.Волокна типу А – це товсті волокна, що мають мієлінових оболонку. У цю групу входять 4 підтипи:
– 1.1.Аa – до них відносяться рухові волокна скелетних м’язів і аферентні нерви, які від м’язових веретен (рецепторів розтягування). Швидкість проведення по них максимальна – 70-120 м / сек
– 1.2.Аb – аферентні волокна, що йдуть від рецепторів тиску і дотику шкіри. 30 – 70 м / сек
– 1.3.Аg – еферентні волокна, що йдуть до м’язових веретенам (15 – 30 м / сек).
– 1.4.Аd – аферентні волокна від температурних і больових рецепторів шкіри (12 – 30 м / сек).
2.Волокна групи В – тонкі міелінізірованние волокна, які є прегангліонарними волокнами вегетативних еферентних шляхів. Швидкість проведення – 3-18 м / сек
3.Волокна групи С, безміеліновие постгангліонарні волокна вегетативної нервової системи. Швидкість 0,5 – 3 м / сек.
Проведення збудження по нервах підпорядковується наступним законам:
1.Закон анатомічної і фізіологічної цілісності нерва. Перша порушується при перерезке, друга – дії речовин, блокуючих проведення, наприклад новокаїну.
2.Закон двостороннього проведення збудження. Воно поширюється в обидва боки від місця роздратування. В організмі найчастіше збудження по аферентні шляхах воно йде до нейрона, а по еферентних – від нейрона. Таке поширення називається ортодромная. Дуже рідко виникає зворотне, або антидромное, поширення збудження.
3.Закон ізольованого проведення. Збудження не передається з одного нервового волокна на інше, що входить до складу цього ж нервового стовбура.
4.Закон бездекрементного проведення. Збудження проводиться по нервах без декремента, тобто загасання. Отже, нервові імпульси послаблюються проходячи по ним.
5.Скорость проведення прямо пропорційна діаметру нерва.
Нервові волокна володіють властивостями електричного кабелю, у якого не дуже хороша ізоляція. В основі механізму проведення збудження лежить виникнення місцевих струмів. У результаті генерації ПД в Аксоплазматичний горбик і реверсії мембранного потенціалу, мембрана аксона набуває протилежний заряд. Зовні вона стає негативною, всередині позитивною. Мембрана нижележащего, невозбужденного ділянки аксона заряджена протилежним чином. Тому між цими ділянками по зовнішній і внутрішній поверхнях мембрани починають проходити місцеві струми. Ці струми деполярізуют мембрану нижележащего невозбужденного ділянки нерва до критичного рівня і в ньому також генерується ПД. Потім процес повторюється і збуджується більш віддалений ділянку нерва і т.д. Т.к. по мембрані безмякотного волокна місцеві струми течуть не перериваючись, тому таке проведення називається безперервним. При безперервному проведенні місцеві струми захоплюють велику поверхню волокна, тому їм потрібен тривалий час для проходження по ділянці волокна. У результаті дальність і швидкість проведення збудження по безмякотним волокнам невелика.
У м’якушевих волокнах ділянки, покриті мієліном, володіють великим електричним опором. Тому безперервне проведення ПД неможливо. При генерації ПД місцеві струми течуть лише між сусідніми перехопленнями. За законом “все або нічого” збуджується найближчий до Аксоплазматичний горбик перехоплення Ранвье, потім сусідній нижележащий перехоплення і т.д. Таке проведення називається сальтаторного (стрибком). При цьому механізмі ослаблення місцевих струмів не відбувається, і нервові імпульси поширюються на велику відстань і з великою швидкістю.
Аксони та дендрити разом з оболонками, що входять до складу периферичних нервів, є нервовими волокнами. Нервові волокна, що мають мієлінову оболонку, називають мієліновими волокнами, а ті, що не мають її, – безмієліновими. Всередині волокна міститься осьовий циліндр з нейрофібрилами. Нейрофібрили складаються з мікротрубочок (діаметр до 30 нм) і нейрофіламентів (до 10 нм). Вони забезпечують транспортування різних речовин і деяких органел по нервових волокнах від тіла нейрона до нервових закінчень і у зворотному напрямку. На периферію транспортуються білки, які формують іонні канали і насоси, медіатори, мітохондрії.
Нервові волокна входять до складу нервів, які іннервують органи чуття і скелетні м'язи, внутрішні органи та судини. Мієлінізоване нервове волокно складається з осьового циліндра і мієлінової оболонки, яка його покриває (рис. 2.1). Мієлінова оболонка створюється внаслідок того, що мієлоцит (шваннівська клітина) багаторазово обгортає осьовий циліндр, шари її зливаються і створюють щільний жировий футляр. Мієлінова оболонка через проміжки рівної довжини розривається і залишає таким чином відкритими ділянки мембрани шириною близько 1 мкм. Ці ділянки одержали назву перехватів Ранв'є. Довжина міжперехватних ділянок пропорційна діаметру волокна. Так, при діаметрі 10-20 мкм довжина проміжку між перехватами становить 1-2 мм. У найтонших волокнах (діаметром 1-2 мкм) ці ділянки мають довжину близько 0,2 мм. Безмієлінові волокна відокремлюються одне від одного тільки шваннівськими клітинами.

Рис. 2.1. Будова мієлінових волокон (схема та електронна мікроскопія):
1 – аксон, 2 – мієлінова оболонка, 3 – шваннівська клітина, 4 – перехват Ранв'є, 5 – ядро шваннівської клітини
У процесах виникнення і проведення нервових імпульсів головну роль відіграє поверхнева мембрана осьового циліндра. Мієлінова оболонка виконує трофічну функцію, а також є електричним ізолятором. Завдяки існуванню мієлінової оболонки потенціал дії, тобто збудження, виникає тільки в перехватах Ранв'є, що має важливе значення для розповсюдження нервового імпульсу вздовж волокна.
Нервові волокна не можуть існувати без зв'язку з тілом нервової клітини: перерізування нерва призводить до загибелі волокон. Регенерація нервів проходить повільно – зі швидкістю 0,5-4,5 мм за Добу.
Для забезпечення росту нейронів та підтримання їх життєдіяльності існує група речовин, яку називають нейротрофінами. До них належать фактор росту нервів (NGF), фактор росту, виділений із головного мозку (BDNF), нейротрофін-3 (NT-З), нейротрофін-4/5 (NT-4/5), нейротрофін-6 (NT-6) та нейротрофін-7 (NT-7).
Нейротрофіни володіють нейропротективними, репаративними та проліферативними властивостями. Вони запускають процеси білкового синтезу, росту, розвитку, відновлення функцій, регулюють синаптичну передачу, забезпечують пластичність нервової системи. Сьогодні встановленим є факт, що не тільки без кисню та глюкози, а й без нейротрофінів нейрони існувати не можуть.
Нейротрофіни переважно синтезуються астроцитами і транспортуються до місць їх контакту з нейронами, де вони зв'язуються зі своїми рецепторами, яких ідентифіковано два типи – тирозинкіназний (Trk) та нейротрофіновий р75. Вони можуть також абсорбуватися нейролемою і ретроградним транспортом переміщатися до тіла нейрона. Є нейрони, які самі виробляють нейротрофіни. У цьому випадку вони транспортуються антиретроградно, досягають синапсів і переходять до наступного нейрона.
Під час ембріонального розвитку нейротрофіни виділяються клітинами-мішенями, наприклад, м'язовим волокном при формуванні нервово-м'язового синапсу. Дифундуючи у напрямку аксона, вони зв'язуються з рецепторами і стимулюють його ріст та формування синапсу. Фактор росту нервів необхідний для розвитку нейронів симпатичної нервової системи, деяких сенсорних нейронів та формування симпатичних гангліїв. У дорослих він зменшує явища апоптозу у нервовій тканині. Виключення гена, відповідального за нейротрофін-3, характеризується втратою механорецепторів шкіри, а відповідального за BDNF – викликає зменшення кількості периферичних чутливих нейронів і дегенеративні зміни у вестибулярних гангліях. У дорослої людини рецепторів до BDNF багато у ділянках центральної нервової системи з високою синаптичною пластичністю.
Завдяки нейротрофінам ушкодження периферичних нервів часто є оборотнім явищем через їх здатність індукувати розгалуження дендритного дерева та ріст аксона. Якщо відбулося зруйнування цілісності нерва, до дистальної частини аксона від проксимальної починають проростати шипики. Цьому сприяють фактори росту, які секретуються шванівськими клітинами, а інгібіторні молекули з периневрію забезпечують його правильний напрямок. За умови досягнення аксона до від'єднаного кінця формується новий синапс. Гальмують регенерацію аксона мієлін, проліферація астроцитів, активація мікроглії, міграція лімфоцитів.
При вивченні проведення збудження по нервових волокнах було встановлено декілька необхідних умов і правил (ЗАКОНІВ) перебігу цього процесу.
1. Анатомічна і фізіологічна цілісність волокна Проведення імпульсів порушується не тільки при механічному руйнуванні волокна, але й при блокуванні натрієвих каналів збудливої мембрани тетродотоксином чи місцевими анестетиками, різкому охолодженні, стійкій деполяризації іонами калію, які можуть накопичуватись при ішемії в міжклітинних щілинах.
Першою речовиною, в якої були виявлені анестезуючі властивості, став кокаїн. У 1860 р. Альберт Німанн, який досліджував його хімічний склад, відзначив, що після жування листя коки зникає чутливість язика. У клініці для місцевої анестезії кокаїн уперше використав Карл Коллер у 1886 р. під час операції на оці. Токсичні ефекти та залежність, яка розвивалася при застосуванні кокаїну, стали поштовхом для пошуку нових синтетичних анестетиків. Механізм дії знеболюючих засобів полягає у блокуванні потенціалзалежних натрієвих каналів, що унеможливлює розвиток деполяризації та поширення збудження по нервовому волокну. Найбільш чутливими до дії анестетиків є немієлінізовані волокна типу С, а найбільш резистентними волокнами – моторні.
- 2. Закон двобічного проведення збудження При подразненні нервового волокна збудження розповсюджується по ньому як у відцентровому, так і в доцентровому напрямках. Двобічне проведення не є тільки лабораторним феноменом. У природних умовах потенціал дії нервової клітини виникає в тій її частині, де тіло переходить в аксон (початковий сегмент, аксонний горбик). Із початкового сегмента потенціал дії розповсюджується у два боки: по аксону в напрямку нервових закінчень і в тілі клітини в напрямку її дендритів.
- 3. Закон ізольованого проведення збудження. У нерві імпульси розповсюджуються вздовж кожного волокна ізольовано, тобто не переходять з одного волокна на інше і впливають тільки на ті

Рис. 2.2. Сальтаторне проведення збудження по мієліновому волокну
клітини ефектора, з якими контактують закінчення цього нервового волокна. Це має важливе значення у зв'язку з тим, що рухові, чутливі та вегетативні волокна периферичного нервового стовбура іннервують різні, розташовані далеко одна від одної, клітини, тканини та органи.
4. Проведення збудження по немієлінізованих та мієлінізованих нервових волокнах. Збудження (ПД) розповсюджується по нервових волокнах без зниження амплітуди ПД і без зниження швидкості, тобто бездекрементно.
Механізм проведення збудження має два компоненти: виникнення ПД в ділянці мембрани, що подразнюється, та подразлива дія на сусідню ділянку кателектротонічного сигналу, який викликається ПД. Проведення ПД – це щось подібне до естафети, у якій кожна ділянка вздовж волокна виступає спочатку як подразнювана, а потім як подразнююча.
Швидкість проведення ПД по безмієлінових волокнах тим більша, чим товстіше волокно і чим нижчий опір зовнішнього середовища.
У мієлінових волокнах проводиться електричний струм і генеруються ПД лише в перехватах Ранв'є. Розповсюдження ПД тут здійснюється стрибкоподібно – сальтаторно (від лат. salto – стрибок) – від перехвату до перехвату (рис 2.2).
Оскільки мієлінові сегменти значно довші за перехвати (1000-2000 мкм проти 1 мкм), то такий спосіб функціонування провідника значно економніший у плані використання іонів (навантаження на іонний насос зменшується) і забезпечує значно більші швидкості проведення збудження. Стрибки ПД через міжперехватну ділянку здійснюються завдяки тому, що амплітуда ПД в 5-6 разів перевищує порогову величину, яка необхідна для збудження сусіднього перехвату.
Час, необхідний для передачі збудження від одного перехвату до іншого, приблизно однаковий у волокон різного діаметра (0,07 мс). Оскільки довжина міжперехватних ділянок пропорційна діаметру нервового волокна, у мієлінізованих волокнах швидкість проведення імпульсів пропорційна їх діаметру.
Нервові волокна класифікують залежно від швидкості проведення збудження, тривалості фаз потенціалу дії, будови волокон (табл. 2.1).
- 5. Відносна невтомлюваність нервового волокна. Нервове волокно проводить ПД значно довший проміжок часу, ніж може відповідати на них орган, який іннервується цим волокном.
- 6. Закон функціональної неспецифічності нервових волокон. Результат збудження залежить не від того, по якому волокну прийшли ПД а від того, який ефектор збуджується або до якого центру вони прямують. Це є підставою для проведення нейропластики в нейрохірургічній практиці.
Таблиця 2.1. Класифікація нервових волокон за Ерлангером – Гассером
ФУНКЦИЮ быстрой передачи возбуждения к нервной клетке и от нее выполняют ее отростки - дендриты и аксоны, т.е нервные волокна. В зависимости от структуры их делят на мякотные, имеющие миелиновую оболочку, и безмякотные. Эта оболочка формируется шванновскими клетками, являющиеся видоизмененными глиальными клетками. Они содержат миелин, который в основном состоит из липидов. Он выполняет изолирующую и трофическую функции. Одна шванновская клетка образует оболочку на 1 мм нервного волокна. Участки, где оболочка прерывается, т.е. 1:е покрыты миелином называют перехватами Ранвье. Ширина перехвата 1 мкм (.рис.). Функционально все нервные волокна делят на три группы: Скорость передачи нервных импульсов по миелинизированным волокнам значительно превышает таковую по волокнам лишенным миелиновой оболочки.
1 Волокна типа А - это толстые волокна, имеющие миелиновую оболочку. В эту группу входят 4 подтипа: двигательные волокна скелетных мышц и афферентные нервы (рецепторов растяжения). Скорость проведения по ним максимальна - 70-120 м,'сек
2 ß (бета) - афферентные волокна, идущие от рецепторов давления и прикосновения кожи. 30-70 м/сек
3 (гамма)- эфферентные волокна, идущие к мышечным веретенам (15-30 м/сек).
4 (сигма) - афферентные волокна от температурных и болевых рецепторов кожи (12-30 м/сек).
Волокна группы В - тонкие миелинизированные волокна, являющиеся преганглионарными волокнами вегетативных эфферентных путей. Скорость проведения - 3-18 м/сек
Волокна группы С, безмиелиновые постганглионарные волокна вегетативной нервной системы. Скорость 0,5 –3 м сек. проведение возбуждения по нервам подчиняется следующим законам:
1. Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезе, вторая - действии веществ блокирующих проведение, например новокаина.
2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаше всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения.
3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола.
4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания.
Следовательно, нервные импульсы не ослабляются, проходя по ним.
5. Скорость проведения прямо-пропорциональна диаметру нерва. (Нервные волокна обладают свойствами центрического кабеля, у которого не очень хорошая изоляция. В основе механизма проведения возбуждения лежит возникновение местных токов: В результате генерации ПД в аксоном холмике и реверсии мембранного потенциала, мембрана аксона приобретает противоположный заряд. Снаружи она становится отрицательной, ^внутри Положительной. Мембрана нижележащего, невозбужденного участка аксона заряжена противоположным образом. ''Поэтому между этими участками, по наружной и внутренней поверхностям мембраны начинают проходить местные, токи. Эти токи деполяризуют мембрану нижележащего невозбужденного участка нерва до критического уровня к в нем также генерируется ПД. Затем процесс повторяется и возбуждается более отдаленный участок нерва и т.д. (рис.). Т.к. по мембране безмякотного волокна местные токи текут не прерываясь, поэтому такое проведение называется непрерывным. При непрерывном проведении местные токи захватывают большую поверхность волокна, поэтому им " требуется, длительное время для прохождения по участку волокна. В результате дальность, и. скорость проведения возбуждения по безмякотным волокнам небольшая. В мякотных волокнах участки, покрытые миелином обладают большим электрическим сопротивлением. Поэтому непрерывное проведение ПД Невозможно. При генерации ПД местные токи текут лишь между соседними, перехватами По закону "все или ничего" возбуждается ближайший к аксонному холмику перехват Ранвье, затем соседний нижележащий перехват и т.д. (рис.). Такое проведение называется сальтаторным (прыжком). При этом
механизме ослабления местных токов не происходит, и нервные импульсы распространяются на большое расстояние и с большой скоростью.
Нервные центры

Аванесян
Свойства нервных центров
Нервным центром (НЦ) называется совокупность нейронов в различных отделах ЦНС, обеспечивающих регуляцию какой-либо функции организма. Например, бульбарный дыхательный центр.
Для проведения возбуждения через нервные центры характерны следующие, особенности:
1. однострочное проведение, оно идет от афферентного, через вставочный к эфферентному нейрону. Это обусловлено наличием межнейронных синапсов.
2. Центральная задержка проведения возбуждения т.е по НЦ возбуждения идет значительно медленнее, чем по нервному волокну. Это объясняется синаптической задержкой т.к больше всего синапсов в центральном звене рефлекторной дуги, там скорость проведения наименьшая. Исходя из этого, время рефлекса - это время от начала воздействия раздражителя до появления ответной реакции. Чем длительнее центральная задержка, тем больше время рефлекса. Вместе с тем оно зависит от силы раздражителя. Чем она больше, тем время рефлекса короче и наоборот. Эго объясняется явлением суммации возбуждений в синапсах. Кроме того, оно определяется и функциональным состоянием ЦНС. Например, при утомлении НЦ длительность рефлекторной реакции увеличивается.
3. Пространственная и временная суммация. Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда ВПСП. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторов нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются 11 и мембране нейрона генерируется распространяющийся ПД.
4. Трансформация ритма возбуждения - изменение частоты нервных импульсов при прохождении через нервный центр. Частота может понижаться или повышаться. Например, повышающая трансформация (увеличение частоты) обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне. Второе, генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких ВПСП и возникновением одного ПД в нейроне.
5. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения
нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы, выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
6. Последействие- это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
7. Тонус нервных центров - состояние постоянной повышенной активности. Он обусловлен постоянным поступлением к НЦ нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например, проявлением тонуса соответствующих центров является тонус определенной группы мышц.
8. автоматия или спонтанная активность нервных центров. Периодическая или постоянная генерация нейронами нервных ИМПУЛЬСОВ, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессор метаболизма в нейронах и действием на них гуморальных факторов.
9. Пластичность нервных центров. Это их способность изменять функциональные свойства. При этом центр приобретает возможность выполнять новые функции или восстанавливать старые после повреждения. В основе пластичности Н.Ц. лежит пластичность синапсов и мембран нейронов, которые могут изменять свою молекулярную структуру.
10. Низкая физиологическая лабильность и быстрая утомляемость.
Н.Ц. могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов. Утомляемость нервных центров - постепенное снижение и полное прекращение ответа при продолжительном раздражении афферентных нервных волокон. Утомление нервных центров вызывается прежде всего нарушением проведения возбуждения в межнейронных синапсах. Тот факт, что утомление сначала возникает в синапсе доказывается простым опытом. В то время, как раздражение афферентного нервного волокна спинальной лягушки не вызывает сокращения мышцы, стимуляция эфферентного волокна приводит к мышечной реакции.
В настоящее время считают, что утомление синапса обусловлено резким снижением запаса медиатора в пресинаптической мембране (истощение), уменьшением чувствительности постсинаптической мембраны (десенсетизация) и уменьшением энергетических ресурсов нейрона. Не все рефлекторные реакции одинаково быстро приводят к развитию утомления. Некоторые рефлексы могут в течение длительного времени протекать без развития утомления. К таким рефлексам относятся проприорецептивные тонические рефлексы
Простейшим нервным центром является нервная цепь, состоящая из трех последовательно соединенных нейронов (рис). Нейроны сложных нервных центров имеют многочисленные связи между собой, образуя нервные сети трех типов: иерархические локальные и дивергентные с одним входом
ИВАНЬКО
Иерархические сети.
Наиболее распространенный тип межнейронных связей можно увидеть в главных сенсорных и двигательных путях.
В сенсорных системах иерархическая организация носит восходящий характер; в нее включаются различные клеточные уровни, по которым информация поступает в высшие центры – от первичных рецепторов к вторичным вставочным нейронам, затем к третичным и т.д.
Двигательные системы организованы по принципу нисходящей иерархии, где команды “спускаются” от нервной системы к мышцам: клетки, расположенные, фигурально говоря, “наверху”, передают информацию специфическим моторным клеткам спинного мозга, а те в свою очередь – определенным группам мышечных клеток.
Ниже лежащий текст прочитать и рассказать о том, как организуются нервные сплетения и что они снабжают на примере шейного узла.




Галетина
Последнее изменение этой страницы: 2016-09-19; Нарушение авторского права страницы
Читайте также:
