
Проводимость нервных импульсов по нервным волокнам
Специалисты, работающие в клинических нейрофизиологических лабораториях, преимущественно имеют дело с двумя группами вопросов: оценкой функционального состояния периферической нервной системы (ПНС) и оценкой состояния коры головного мозга. Для оценки состояния ПНС применяют методы исследования нервной проводимости (ИНП). При этом исследуемый нерв стимулируют электрическим током с одновременной регистрацией электрических волн, которые нерв генерирует в ответ на раздражение. Второй метод оценки состояния ПНС — электромиография (ЭМГ), при которой в исследуемых мышцах регистрируют волны, возникающие при их произвольном сокращении. Комплекс методов ИНП и ЭМГ называют электродиагностическим обследованием.
При подозрении на поражение ПНС после клинического осмотра проводят исследование нервной проводимости (ИНП). С помощью электрической стимуляции нерва и последующей регистрации деполяризации связанного с этим нервом мышечного волокна можно определить, поражает ли заболевание нерв, нервно-мышечный синапс или саму мышцу. ИНП также позволяет установить, является ли заболевание фокальным или диффузным, повреждает чувствительные и/или двигательные аксоны, поражает миелиновую оболочку или сам аксон.
а) Исследование проведения по нервам верхней конечности. Основной нерв верхней конечности, исследование проводимости которого позволяет выявить наличие фокального (в отличие от генерализованного) поражения периферической нервно-мышечной системы,— срединный нерв. Срединный нерв — одновременно двигательный и чувствительный — обладает тремя основными преимуществами в плане проведения электрофизиологической диагностики.
1. В области локтевого сустава и запястья он расположен близко к поверхности, поэтому технически просто проводить стимуляцию нерва и выполнять регистрацию его реакции.
2. Срединный нерв иннервирует короткую мышцу, отводящую большой палец кисти, которая хорошо подходит для проведения поверхностной и игольчатой ЭМГ (двигательное ИНП).
1. Проведение по двигательным нервам:
- Стимуляция. Обычный стимулирующий электрод имеет катод и анод в форме двух тупых штырей, которые прикладывают к поверхности кожи над нервом. На рисунке ниже изображен электрод, размещенный над срединным нервом у запястья (сразу латеральнее сухожилия длинной ладонной мышцы). Для того чтобы анод не заблокировал проведение нервного импульса, катод должен быть расположен к месту регистрации ближе, чем анод. Когда между катодом и анодом возникает электрический ток достаточной силы, изменение трансмембранного тока ионов вызывает появление нервного импульса, который распространяется по нерву в обоих направлениях. Сначала деполяризация возникает на крупных миелинизированных нервных волокнах, расположенных вблизи катода; к ним относят Аα-аксоны мотонейронов переднего рога спинного мозга. Обычно стимул в 20-40 mА продолжительностью 0,1 мс достаточен для возбуждения всех двигательных единиц короткой мышцы, отводящей большой палец.
- Регистрация. Активный поверхностный датчик, имеющий в нашем случае форму диска, устанавливают к двигательной точке. Это место, расположенное в центре исследуемой мышцы, имеет наибольшую концентрацию двигательных концевых пластинок. Второй, референтный электрод устанавливают на нейтральную область на некотором удалении от первого электрода. Для записи разницы потенциалов между этими двумя электродами используют усилитель, который также увеличивает выраженность вызванной двигательной реакции мышцы. Система настроена таким образом, что при регистрации активным электродом отрицательного значения кривая на мониторе отклоняется вверх.
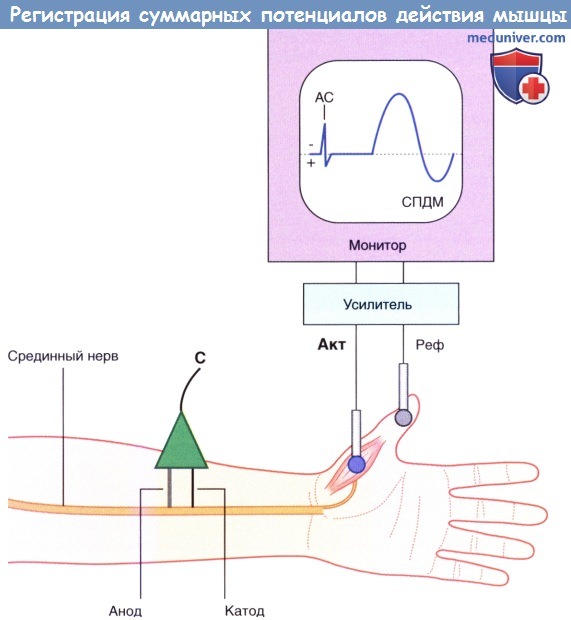
Базовая установка для регистрации СПДМ. АС — артефакт стимуляции.
Стимулирующий электрод (С) расположен над срединным нервом.
Активный электрод (Акт) расположен над короткой мышцей, отводящей большой палец.
Референтный электрод (Реф) установлен дистальнее.
При подаче слабого стимула на мониторе будет отображаться только плоская линия, которая иногда прерывается стимуляционными артефактами. При увеличении интенсивности стимула появляются небольшие суммарные потенциалы действия мышцы (СПДМ). Они возникают за счет активации крупных миелинизированных аксонов, расположенных вблизи стимулирующего электрода. Волна деполяризации, передающаяся по аксону, приводит к деполяризации всех мышц, которые иннервирует этот аксон. В двигательных единицах внутренних мышц руки, в том числе в короткой мышце, отводящей большой палец, один мотонейрон иннервирует 200-300 мышечных волокон. Для крупных мышц, которые не совершают точных мелких движений (например, дельтовидная, икроножная), минимальное отклонение кривой на мониторе будет в несколько раз больше. Это связано с двумя факторами: соотношение мотонейронов к двигательным волокнам составляет примерно 1:1000, а их крупные мышечные волокна генерируют потенциалы действия большей амплитуды.
Следует еще раз подчеркнуть, что кривая, возникающая на мониторе, не отображает сокращения мышц, а представляет результат внеклеточного потенциала, который возникает за счет деполяризации мембран мышечных клеток и передается через подкожные ткани и кожу. Несмотря на это, при большинстве мышечных заболеваний патологический процесс также будет нарушать процесс деполяризации клеточной мембраны, что приведет к появлению аномалий на кривой.
При повышении вольтажа происходит возбуждение дополнительных двигательных единиц. Это происходит до тех пор, пока все двигательные единицы не будут активироваться одним импульсом. Такой стимул называют максимальным. Для точности окончательный стимул обычно делают супрамаксималъным, т.е. на 5-10 % превышающим максимальный. Окончательная наблюдаемая волна и представляет собой СПДМ — сумму потенциалов действия отдельных мышечных волокон.
Результат измерения конечного СПДМ изображен на рисунке ниже. К измеряемым параметрам относят латентность (временной интервал) между временем подачи стимула и началом деполяризации, амплитуду и длительность отрицательной фазы волны. (Появление последней, положительной фазы, обусловлено движением ионов внутрь клетки во время общей реполяризации мышечных волокон.)
- Повторное измерение. Целесообразно повторно измерить СПДН, но уже с другого нерва. Обычно для повторного измерения используют локтевой нерв. Точка С1 расположена на запястье сразу латеральнее локтевого сгибателя запястья, а С2 — на уровне выхода нерва у медиального надмыщелка. Активный электрод устанавливают над мышцами гипотенара у медиального края ладони.
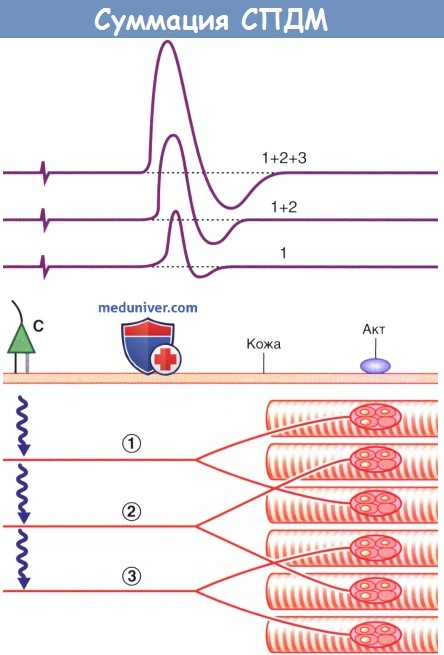
Суммация СПДМ. Двигательные единицы представлены перекрещивающимися парами мышечных волокон.
При легкой (1), средней (2) и максимальной (3) стимуляции на экране возникают волны все большей амплитуды, хотя они и являются разными с точки зрения физиологии феноменами. 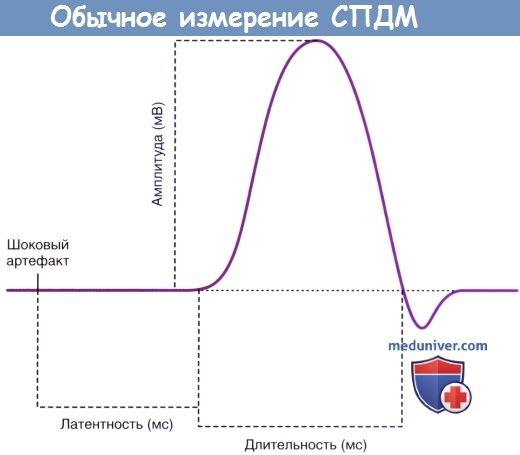
Обычное измерение СПДМ 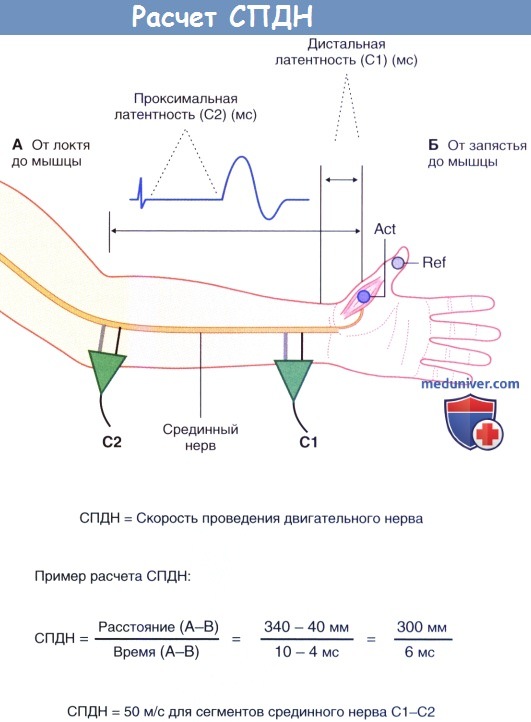
Расчет скорости проведения двигательного нерва (СПДН). Нерв стимулируют дважды: С1—первый стимул, С2 — второй стимул.
Двунаправленными стрелками отмечены два измерения длины.
Исходное значение времени не представлено. Внизу показан пример расчета нормальной скорости проведения.
2. Проведение по чувствительным нервам. Для измерения скорости проведения чувствительных нервов (СПЧН) также удобнее всего использовать срединный нерв. Точно также при стимуляции происходит возбуждение крупных миелинизированных нервных волокон, места и характер стимуляции в локтевой ямке и у запястья остаются прежними. Однако в данном случае избирательно записывают антидромные потенциалы кожных чувствительных волокон, в частности пальцевых ветвей срединного нерва, идущих к коже указательного пальца. Для этого на указательный палец надевают активное записывающее устройство в форме кольца.
1. Физиологическая временная дисперсия. Подобно тому, как во время гонки расстояние между бегунами прогрессивно увеличивается, наиболее быстрый проводник импульса выходит вперед, а самый медленный остается позади. Из-за этого при увеличении расстояния измерения кривая СЧПД удлиняется. Данное явление и называют временной дисперсией (рассеивание во времени).
- Скорость проведения чувствительных нервов. Основные режимы и принципы измерения остаются теми же, что и при исследовании суммарных потенциалов действия мышцы (СПДМ). На рисунке ниже изображен пример, на котором показан феномен отмены фазы при наличии физиологической временной дисперсии.
- Повторное измерение. Обычно проводят с локтевым нервом. Как и ранее, нерв стимулируют у запястья и у локтевой ямки, кольцо надевают на мизинец.
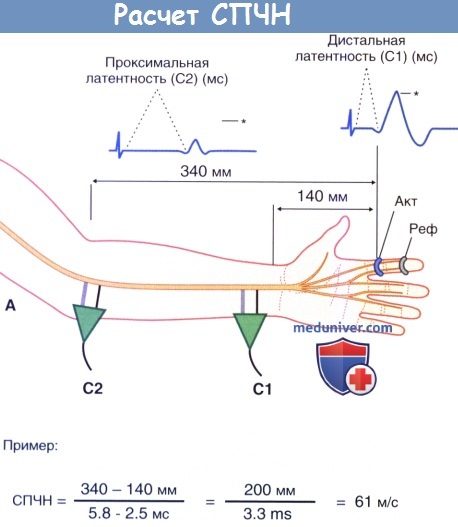
Расчет скорости проведения чувствительных нервов (СПЧН). Представлены пальцевые ветви срединного нерва. Основные принципы расчета те же, что и в случае скорости проведения двигательного нерва (СПДН).
б) Проведение по нервам нижней конечности:
1. Проведение по двигательным нервам. При исследовании СПДМ на нижней конечности чаще всего используют глубокий малоберцовый нерв, регистрацию осуществляют с короткого разгибателя пальцев на подошвенной поверхности стопы. Сначала глубокий малоберцовый нерв стимулируют на передней поверхности лодыжки, а затем на уровне шейки малоберцовой кости. Иногда регистрацию осуществляют также с передней большеберцовой мышцы; в этом случае общий малоберцовый нерв сначала стимулируют у шейки малоберцовой кости, а затем у латерального края подколенной ямки вблизи сухожилия двуглавой мышцы бедра.
Повторное исследование СПДМ проводят со стимуляцией большеберцового нерва, запись осуществляют с мышцы, приводящей большой палец, расположенной у медиального края стопы.
2. Проведение по чувствительным нервам. Для исследования СЧПД используют икроножный нерв. Он начинается от большеберцового нерва и получает ветвь от общего малоберцового нерва; данный нерв иннервирует кожу вдоль латерального края стопы. Запись осуществляют с кожи, расположенной немного ниже латеральной лодыжки, далее нерв антидромно стимулируют в точках, показанных на рисунке ниже.
• на уровне шеи, где корешки спинномозговых нервов С6 и С7 предрасположены к сдавливанию остеофитами, возникающими при шейном спондилезе;
• на уровне поясницы, где корешки спинномозгового нерва S1 могут сдавливаться при пролапсе межпозвоночного диска L5/S1;
• радикулопатия может быть компонентом генерализованной периферической нейропатии.
Н-рефлекс. Поскольку корешки спинномозговых нервов залегают глубоко, оценить проводимость по ним можно лишь косвенно за счет активации сенсомоторных чувствительных дуг соответствующего уровня. Стандартный тест, названный в честь Hoffman, который впервые его описал, называют Н-ответом или Н-рефлексом. Его часто используют для оценки общей скорости проведения по рефлекторной дуге S1; оценивают те же самые нейроны, которые клинически исследуют путем ахиллова рефлекса. Большеберцовый нерв стимулируют длительным током минимальной силы, достаточной для сокращения мышцы. Цель исследования — возбуждение самых крупных афферентных миелинизированных волокон (предпочтительно длительными стимулами), преимущественно тех, которые иннервируют аннулоспиральные окончания мышечных веретен. За счет этого активируется моносинаптический рефлекс с минимальной латентностью, происходит сокращение трехглавой мышцы голени (икроножной/камбаловидной). Минимальная латентность оказывается достаточно долгой — до 35 мс в зависимости от роста пациента, поскольку сегмент S1 спинного мозга расположен за телом позвонка L1; за счет этого нервный импульс должен пройти путь порядка 130-150 см вниз и вверх. При последовательном повышении силы тока возникает точка, в которой появляются М-волны. М-волны возникают благодаря прямой ортодром-ной активации двигательных концевых пластинок. Антидромное проведение обусловливает постепенное затухание потенциалов действия, нисходящих по эфферентному звену Н-рефлекса.
На верхней конечности проведение по нервным корешкам нерва С6 можно исследовать за счет стимуляции срединного нерва и регистрации сокращений лучевого сгибателя запястья. Корешок С7 можно исследовать путем стимуляции заднего кожного нерва предплечья и регистрации сокращений трехглавой мышцы плеча.
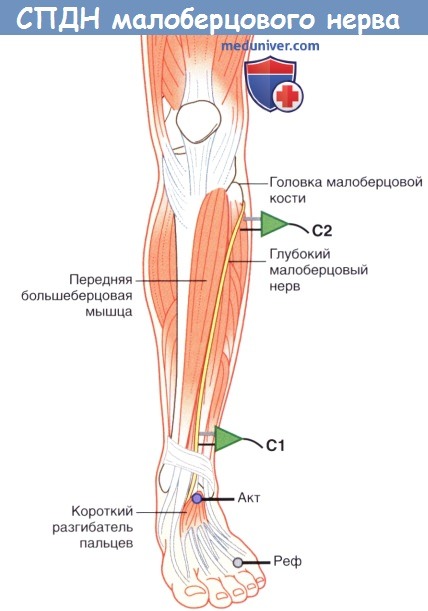
Положение датчика и усилителя при измерении скорости проведения двигательного нерва (СПДН) малоберцового нерва. 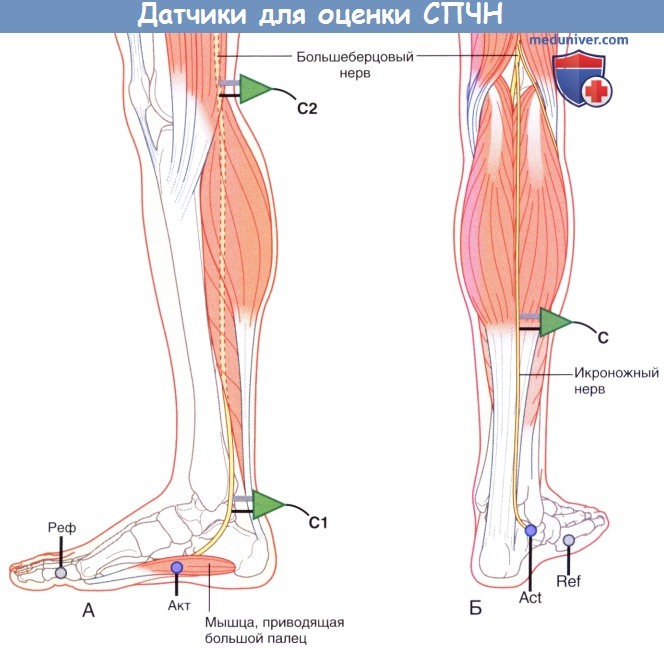
(А) Положение датчика и стимулятора при измерении скорости проведения чувствительного нерва (СПЧН) с большеберцового нерва.
(Б) Положение электродов при записи антидромной скорости проведения чувствительного нерва (СПЧН) с икроножного нерва. 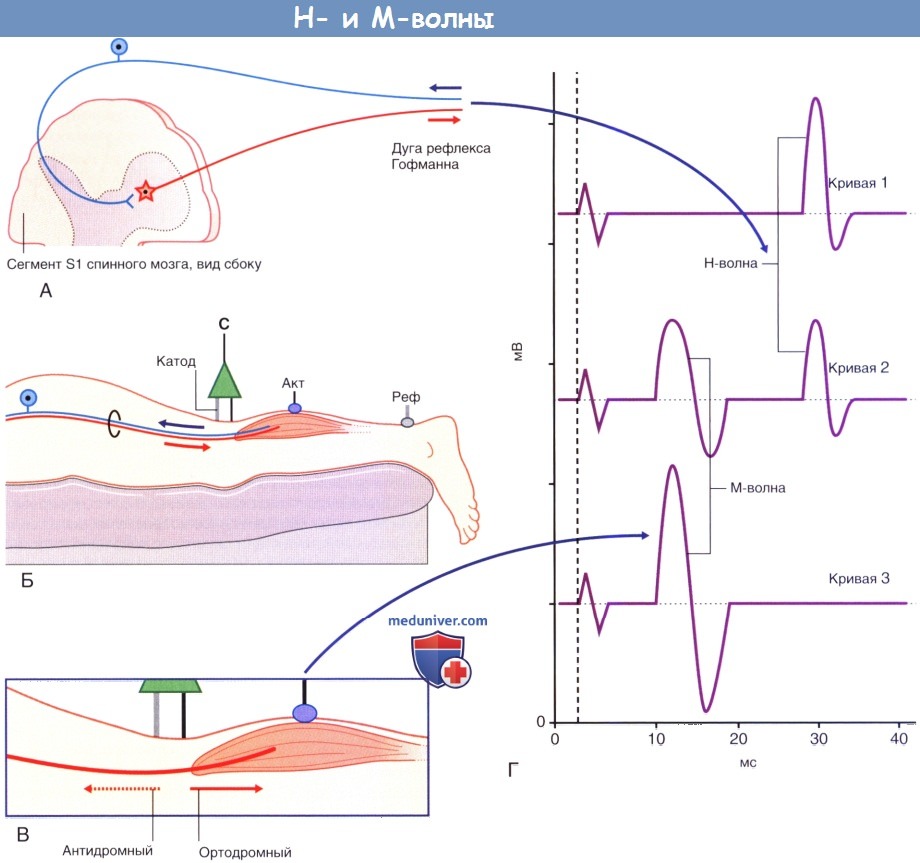
Анатомические предпосылки появления Н- и М-волн.
(А) Появление Н-волны опосредованно моносинаптической рефлекторной дугой, как показано на рисунке.
(Б) Запись СПДМ с S1 сегмента рефлекторной дуги Ахиллова рефлекса. Стимулирующий электрод расположен над большеберцовым нервом; записывающий электрод расположен над трехглавой мышцей.
Оба звена рефлекторной дуги образованы большеберцовой ветвью седалищного нерва.
(В) При повышении силы тока происходит непосредственная активация аксонов, иннервирующих мышцу, из-за чего возникают коротколатентные М-волны.
(Г) Обратите внимание, что по мере увеличение интенсивности стимула Н-волна постепенно исчезает (от кривой 1 до кривой 3) вследствие того,
что ортодромные двигательные импульсы из (А) подавляются антидромными импульсами катода, которые представлены в (В) пунктирной линией.
Редактор: Искандер Милевски. Дата публикации: 13.11.2018
В этой книге предельно сжато изложен курс лекций по нормальной физиологии. Благодаря четким определениям основных понятий студент может сформулировать ответ, за короткий срок усвоить и переработать важную часть информации, успешно сдать экзамен. Курс лекций будет полезен не только студентам, но и преподавателям.
- ЛЕКЦИЯ № 1. Введение в нормальную физиологию
- ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
- ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
- ЛЕКЦИЯ № 4. Физиология мышц
- ЛЕКЦИЯ № 5. Физиология синапсов
- ЛЕКЦИЯ № 6. Физиология центральной нервной системы
- ЛЕКЦИЯ № 7. Физиология различных разделов ЦНС
Приведённый ознакомительный фрагмент книги Нормальная физиология: конспект лекций (С. С. Фирсова) предоставлен нашим книжным партнёром — компанией ЛитРес.
ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
1. Физиология нервов и нервных волокон. Типы нервных волокон
Физиологические свойства нервных волокон:
1) возбудимость – способность приходить в состояние возбуждения в ответ на раздражение;
2) проводимость – способность передавать нервные возбуждение в виде потенциала действия от места раздражения по всей длине;
3) рефрактерность (устойчивость) – свойство временно резко снижать возбудимость в процессе возбуждения.
Нервная ткань имеет самый короткий рефрактерный период. Значение рефрактерности – предохранять ткань от перевозбуждения, осуществляет ответную реакцию на биологически значимый раздражитель;
4) лабильность – способность реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом импульсов возбуждения за определенный период времени (1 с) в точном соответствии с ритмом наносимых раздражений.
Нервные волокна не являются самостоятельными структурными элементами нервной ткани, они представляют собой комплексное образование, включающее следующие элементы:
1) отростки нервных клеток – осевые цилиндры;
2) глиальные клетки;
3) соединительнотканную (базальную) пластинку.
Главная функция нервных волокон – проведение нервных импульсов. Отростки нервных клеток проводят сами нервные импульсы, а глиальные клетки способствуют этому проведению. По особенностям строения и функциям нервные волокна подразделяются на два вида: безмиелиновые и миелиновые.
Безмиелиновые нервные волокна не имеют миелиновой оболочки. Их диаметр 5–7 мкм, скорость проведения импульса 1–2 м/с. Миелиновые волокна состоят из осевого цилиндра, покрытого миелиновой оболочкой, образованной шванновскими клетками. Осевой цилиндр имеет мембрану и оксоплазму. Миелиновая оболочка состоит на 80 % из липидов, обладающих высоким омическим сопротивлением, и на 20 % из белка. Миелиновая оболочка не покрывает сплошь осевой цилиндр, а прерывается и оставляет открытыми участки осевого цилиндра, которые называются узловыми перехватами (перехваты Ранвье). Длина участков между перехватами различна и зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами. При диаметре 12–20 мкм скорость проведения возбуждения составляет 70—120 м/с.
В зависимости от скорости проведения возбуждения нервные волокна делятся на три типа: А, В, С.
Наибольшей скорость проведения возбуждения обладают волокна типа А, скорость проведения возбуждения которых достигает 120 м/с, В имеет скорость от 3 до 14 м/с, С – от 0,5 до 2 м/с.
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
- ЛЕКЦИЯ № 1. Введение в нормальную физиологию
- ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
- ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
- ЛЕКЦИЯ № 4. Физиология мышц
- ЛЕКЦИЯ № 5. Физиология синапсов
- ЛЕКЦИЯ № 6. Физиология центральной нервной системы
- ЛЕКЦИЯ № 7. Физиология различных разделов ЦНС
Приведённый ознакомительный фрагмент книги Нормальная физиология: конспект лекций (С. С. Фирсова) предоставлен нашим книжным партнёром — компанией ЛитРес.
Потенциалы действия (импульсы возбуждения) обладают способностью распространяться вдоль по нервным и мышечным волокнам. Потенциалы могут быть локальными, они распространяются на небольшие расстояния 1-2 мм с затуханием (декрементом) и импульсными. Импульсные потенциалы распространяются без декремента на значительные расстояния – до нескольких десятков сантиметров. Локальные потенциалы возникают в ответ на действие подпорогового раздражителя, например, на мембране рецепторной клетки. Если локальное возбуждение попадает в участок мембраны, способной генерировать ПД, и амплитуда локального тока достигает критического уровня деполяризации, формируется ПД, который распространяется по всей длине нервного волокна.
Передача информации на большие расстояния в пределах нервной системы осуществляется с помощью нервных импульсов по аксонам нейронов. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциал чувствительных ионных каналов. В зависимости от расположения и концентрации ионных каналов в мембране волокна выделяют два способа проведения нервного импульса.
1. Непрерывное проведение нервного импульсаосуществляется в безмиелиновых волокнах, объясняется равномерным распределением потенциал чувствительных ионных каналов, участвующих в генерации ПД.
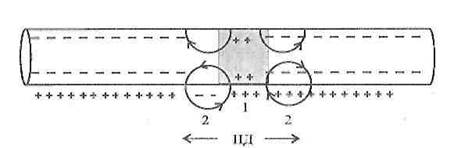
Рисунок 3 - Непрерывное распространение ПД в нервном волокне
Возникший ПД обеспечивает открытие потенциал зависимых Na-каналов на соседнем участке мембраны нервного волокна и движение ионов Na + внутрь волокна, что обеспечивает развитие критического уровня деполяризации на соседнем участке нервного волокна и возникновение нового ПД. Непрерывное распространение нервного импульса идет через генерацию новых импульсов по эстафете, когда каждый возникший импульс является раздражителем для соседнего участка нервного волокна и обеспечивает возникновение нового ПД.
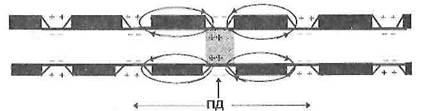
Рисунок 4 – Сальтаторное распространение ПД в нервных волокнах
2. Сальтаторное проведение нервного импульса (ПД) осуществляется в миелиновых волокнах, так как у них потенциал чувствительные ионные каналы локализованы только в участках мембраны перехватов Ранвье, где их плотность достигает 12 000 на 1 мкм 2 . В области межузловых сегментов, обладающих высокими изолирующими свойствами, потенциал чувствительных каналов нет, вследствие чего мембрана осевого цилиндра там практически невозбудима. Поэтому ПД, возникший в одном перехвате Ранвье распространяется через межузловой сегмент до соседнего перехвата, деполяризует мембрану до критического уровня и вызывает возникновение потенциала действия.
Сальтаторное проведение нервных импульсов является эволюционно более поздним механизмом, возникшим впервые у позвоночных в связи с миелинизацией нервных волокон. Оно имеет два важных преимущества по сравнению с непрерывным механизмом проведения возбуждения:
-более экономично по затрате энергии, так как возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% от площади мембраны волокна, следовательно, требуется меньше энергии для восстановления трансмембранных градиентов ионов натрия и калия;
-возбуждение проводится с большей скоростью (до 120 м/с), чем в безмиелиновых волокнах (0,5-2,0 м/с).
В связи с этими приемуществами миелиновые волокна в нервной системе сформировались там, где необходима наиболее быстрая регуляция функций. Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей.
Проведение нервных импульсов по волокнам нейронов починяется определенным законам:
Закона 1: нервный импульс распространяется в обе стороны от места раздражения.
Закон 2: проведение нервного импульса по волокну происходит изолировано и не распространяется на параллельные волокна. Объяснение этого закона заключается в том, что аксолемма имеет очень высокое сопротивление и не пропускает петли тока на невозбужденные волокна, расположенные рядом. Изолированное проведение обеспечивает высокую точность регуляторной деятельности ЦНС.
Закон 3: скорость проведения возбуждения по нервному волокну определяется его диаметром. Отсюда следствие: чем толще нервное волокно, тем больше скорость проведения нервного импульса по этому волокну.
Закон 4: нерв сохраняет способность к проведению возбуждения в течение 6-8 часов непрерывного раздражения (закон Н.Е. Введенского, 1883).
Закон 5: действие веществ, блокирующих работу ионных каналов, без нарушения целостности нервного волокна вызывает состояние обратимого парабиоза, (закон парабиоза, Введенский, 1901).
Кроме того, выделяют несколько видов распространения возбуждения:
1. Иррадиация (дивергенция) возбуждения в ЦНС, которая объясняется ветвлением аксонов и наличием вставочных нейронов, аксоны которых также ветвятся. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов.
Рисунок 4 – Дивергенция афферентных дорсальных корешков на нейроны головного мозга (а – дивергенция, б – конвергенция)
2. Конвергенция возбуждения представляет собой схождение нескольких нервных импульсов, идущих по разным путям к одному и тому же нейрону. Явление конвергенции распространения возбуждения описал Э. Шеррингтон, поэтому явление было названо принцип шеррингтоновской воронки или принцип общего конечного пути. Примером может служить конвергенция возбуждения на спинальном мотонейроне. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг.
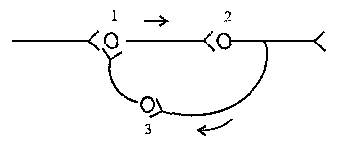
Рисунок 5 – Циркуляция возбуждения в замкнутых нейронных цепях
3. Циркуляция возбуждения по замкнутым нейронным цепям, возникает в результате замыкания группы нейронов в кольцевую структуру. Циркуляция возбуждения – одна из причин явления последействия. Считают, что циркуляция возбуждения в замкнутых нейронных цепях наиболее вероятный механизм феномена кратковременной памяти. Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом.
Литература
1. Смирнов, В.М. Физиология сенсорных систем и высшая нервная деятельность / В.М. Смирнов, С.М. Будылина. - М.: Медицина, 2003. - 304 с.
2. Шульговский, В. В. Основы нейрофизиологии: Учебное пособие для студентов вузов. - М.: Аспект Пресс, 2000. - с. 277.
3. Батуев, А.С. Физиология поведения. Нейрофизиологические закономерности / А.С. Батуев. - Л.: Наука, 1986. - 340 с.
4. Александров, Ю.И. Психофизиология / Ю.И.Александров. - М.: Медицина, 2001. - 230 с.
Данилова, Н.Н. Физиология высшей нервной деятельности / Н. Н. Данилова, А.Л. Крылова. - Ростов н/Д: Феникс, 1999. – 480 с.
[1] ионоселективные каналы и ионные насосы
[2] правило действует и в обратном направлении
[3] 1780 год, профессор анатомии Болонского университета
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
Читайте также:
