
Распространение нервных импульсов по нервным волокнам
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
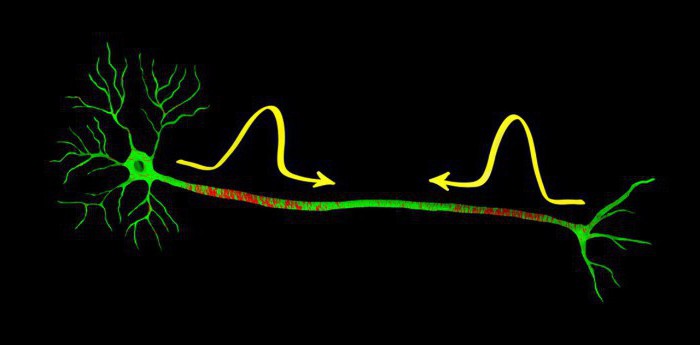
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.
Особенность нервного импульса (потенциала действия) является его самораспространение по нервному или мышечному волокну, в результате которого обеспечивается передача информации от периферических рецепторных окончаний к нервным центрам, а от них к эффекторам. В мышечных клетках нервный импульс оказывает пусковое влияние на процессы, активизирующие сократительный аппарат. Распространение неровного импульса начинается с момента, когда внутренняя часть нервного волокна заряжается положительно, и разность потенциалов между внутренней средой нерва и наружной может достигать 40-50 мВ.
Проведение нервного импульса можно сравнить с распространением пламени по бикфордовому шнуру: импульс возникает при пороговом запале, идет с определенной скоростью без затухания, передаваясь от возбужденного участка к соседнему – невозбужденному.
В основе объяснения этого механизма лежит теория немецкого физика Л. Германа (1879) и затем А. Ходжкина (1937). Согласно этой теории возникший в точке раздражения потенциал действия является источником раздражения соседнего невозбужденного участка волокна. Это происходит вследствие возникновения круговых, или местных токов между возбужденным (т.е. отрицательно заряженным) и соседним (положительно заряженным) участками мембраны. (рис **)
РИС
В результате местного электро-химического сдвига ионной проницаемости мембраны возникает ее деполяризация и достигается критический пороговый потенциал действия. В зоне первоначально возбужденной в это время восстанавливается потенциал покоя. Затем потенциал действия возникает на следующем участке волокна и т.д. Поэтому волна возбуждения проходит вдоль волокна, не затухая и не поворачивая назад, ибо на соседнем пройденном участке находится рефрактерная зона. Важное свойство возбудимых тканей рефрактерность. Она определяет прерывистости импульсов. В естественных условиях по нервам непрерывно бегут нервные импульсы. Частота этих ритмических зарядов зависит от силы раздражения. Двигательные нейроны могут проводить около 500 импульсов в секунду, промежуточные – 1000.
Таким образом, распространение (проведение) возбуждения заключается в последовательном возникновении и исчезновении потенциала действия на протяжении нервного или мышечного волокна.
Непрерывное проведение импульсов характерно для мышечного волокна и безмякотных, безмиелиновых нервных волокон, имеющих только шванновскую оболочку. Таких волокон в нервной системе меньшинство. В мякотных нервных волокнах, имеющих миелиновую оболочку (она является хорошим изолятором) круговые токи могут возникать лишь между двумя соседними (возбужденными и невозбужденными) перехватами Ранвье, где миелин отсутствует. Следовательно, возбуждение в миелинизированных нервных волокнах распространяется скачкообразно, сальтаторно (la salto – скачу, прыгаю). Скорость проведения импульса в нервных волокнах определяется их гистостроением и диаметром. В миелинизированных нервных волокнах она составляет 30-120 м/сек (6 км/мин, 360 км/ч), в безмякотных - 0,5-3 м/сек, в скелетных 5 м/сек. (рис **)
РИС
Особенности проведении нервного импульса:
- возбуждение проводится в обе стороны по нервному волокну от места раздражения;
- проведение возбуждения возможно лишь при целостности волокна;
- более толстые волокна обладают наиболее низким порогом возбуждения;
- волокна, входящие в состав одиночных или смешанных нервов проводят возбуждение изолированно, т.е. не переходя на другие волокна и адресуются лишь своим клеткам;
Проявлением возбуждения нервных волокон является повышение обменных процессов. Возрастает расход кислорода и выделение СО2 , увеличивается расход АТФ, повышается образование молочной кислоты. В окончаниях аксонов нервных клеток при возбуждении секретируются особые химические вещества – медиаторы, оказывающие возбуждающее или тормозящее влияние на иннервируемые ткани.
Дата добавления: 2016-06-29 ; просмотров: 4228 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Если в каком-нибудь участке возбудимой мембраны сформировался потенциал действия, мембрана деполяризована, возбуждение распространяется на другие участки мембраны. Рассмотрим распространение возбуждения на примере передачи нервного импульса по аксону.
Локальные токи при распространении нервного импульса по нервному волокну
Может возникнуть вопрос, почему возбуждение распространяется по аксону не в обе стороны от зоны, до которой дошло возбуждение, ведь локальные токи текут в обе стороны от возбужденного участка. Дело в том, что возбуждение может распространяться только в область мембраны, находящуюся в состоянии покоя, то есть в одну сторону от возбужденного участка аксона. В другую сторону нервный импульс не может распространяться, так как области, через которые прошло возбуждение, некоторое время остаются невозбудимыми - рефракторными.
Повышение мембранного потенциала - величина деполяризующего потенциала V, передаваемого от возбужденных участков вдоль мембраны, зависит от расстояния х (как это следует из электродинамики) по формуле:
Vo - повышение мембранного потенциала в зоне возбуждения, х - расстояние от возбужденного участка; l - константа дины нервного волокна, на котором деполяризующий потенциал уменьшается в е раз.
Зависимость деполяризующего потенциала от расстояния
Рис. Константа длины нервного волокна
где rm - удельное электрическое сопротивление оболочки волокна, l - толщина оболочки, r - радиус нервного волокна, rа - удельное электрическое сопротивление цитоплазмы.
Чем больше константа длины мембраны, тем больше скорость распространения нервного импульса. Величина l тем больше, чем больше радиус аксона и удельное сопротивление мембраны и меньше удельное сопротивление цитоплазмы.
распространение нервного импульса
Сальтаторное распространение потенциала действия по миелинизированному волокну
Синаптическая передача
Нервная клетка (нейрон) состоит изтела клетки (сомы), отростков(аксонов и дендритов) и концевых пластинок. С помощью дендритов нейроны воспринимают, а посредством аксонов передают возбуждение. На периферии аксоны покрытышванновскими клетками, образующими миелиновую оболочку с высокими изолирующими свойствами.
Миелин содержит много холестерина и мало белка; его удельное сопротивление выше удельного сопротивления других биологических мембран. Кроме того, толщина миелиновой оболочки в сотни раз превышает толщину обычной клеточной мембраны.
Передача сигналов между нейронами и от нейронов к мышечным клеткам (так называемая нейронейрональная и нейромышечная трансдукция) происходит в нервных окончаниях (синапсах) с помощью сигнальных веществ, медиаторов.
Нейромедиаторы — короткоживущие вещества локального действия; они выделяются в синаптическую щель и передают сигнал соседним клеткам.Нейрогормоны — долгоживущие вещества дальнего действия, поступающие в кровь. Однако граница между двумя группами достаточно условная, поскольку большинство медиаторов одновременно действует как гормоны.
Сигнальныевещества-нейромедиаторы(или нейромодуляторы) должны удовлетворять ряду критериев. Прежде всего они должны продуцироваться нейронами и храниться в синапсах; при поступлении нервного импульса они должны выделяться в синаптическую щель, избирательно связываться со специфическим рецептором на постсинаптической мембране другого нейрона или мышечной клетки, стимулируя эти клетки к выполнению ими своих специфических функций.
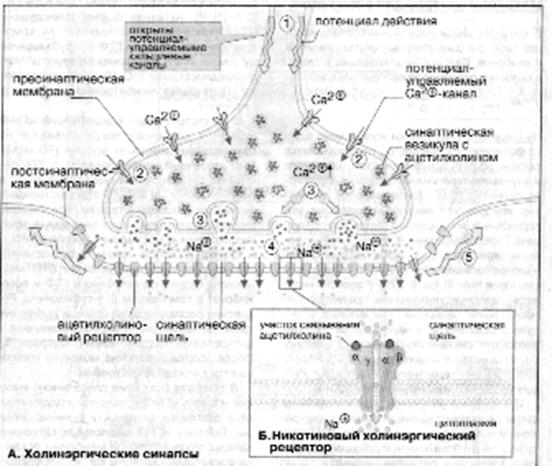
Синапсы образованы мембранами двух контактирующих клеток, пресинаптической и постсинаптической,которые разделены узкойсинаптическойщелью. Медиатор выделяется в синаптическую щель за счет экзоцитоза, диффундирует крецепторам постсинаптической мембраны, связывается с ними и передает сигнал соседней клетке. Белки-рецепторы — это лиганд-активируемые ионные каналы, либо мембранные белки, которые управляют ионными каналами посредством G-белков.
Процесс передачи сигнала включает следующие этапы.Потенциал действия достигает пресинаптической мембраны (1). Это вызывает открываниепотенциал-управляемых Са 2+ -каналов (2). Ионы Са 2+ проникают из внеклеточного пространства в клетку, их уровень в синапсе резко увеличивается, что инициирует процесс экзоцитоза.Синаптические везикулы выделяют содержимое (ацетилхолин) в синаптическую щель (3). Молекулы ацетилхолина диффундируют через синаптическую щель, связываются с постсинаптическими рецепторами и активируют их (4). Поток ионов Na + изменяет потенциал покоя постсинаптической мембраны нервной или мышечной клетки настолько, что открываются соседние потенциал-управляемые Na + каналы и возникаетпотенциал действия.
Дата добавления: 2018-04-15 ; просмотров: 473 ;
Нервный импульс (лат. nervus нерв; лат. impulsus удар, толчок) — волна возбуждения, распространяющаяся по нервному волокну; единица распространяющегося возбуждения.
Нервный импульс обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам — скелетной мускулатуре, гладким мышцам внутренних органов и сосудов, железам внутренней и внешней секреции и т. д.
Распространение Нервных импульсов отождествляется с проведением потенциалов действия (см. Биоэлектрические потенциалы). Возникновение возбуждения может быть результатом раздражения (см.), напр, воздействие света на зрительный рецептор, звука на слуховой рецептор, или процессов, протекающих в тканях (спонтанное возникновение Н. и.). В этих случаях Н. и. обеспечивают согласованную работу органов при протекании какого-либо физиологического процесса (напр., в процессе дыхания Н. и. вызывают сокращение скелетных мышц и диафрагмы, результатом чего являются вдох и выдох, и т. д.).
В живых организмах передача информации может осуществляться и гуморальным путем, посредством выброса в русло крови гормонов, медиаторов и т. п. Однако преимущество информации, передаваемой при помощи Н. и., состоит в том, что она более целенаправленна, передается быстро и может быть точнее закодирована, чем сигналы, посылаемые гуморальной системой.
Факт, что нервные стволы являются путем, по к-рому передаются влияния от мозга к мышцам и в обратном направлении, был известен еще в эпоху античности. В средние века и вплоть до середины 17 в. считалось, что по нервам распространяется некая субстанция, подобная жидкости или пламени. Идея о электрической природе Н. и. возникла в 18 в. Первые исследования электрических явлений в живых тканях, связанных с возникновением и распространением возбуждения, были осуществлены Л. Гальвани. Г. Гельмгольц показал, что скорость распространения Н. и., к-рую ранее считали близкой к скорости света, имеет конечное значение и может быть точно измерена. Германн (L. Hermann) ввел в физиологию понятие потенциала действия. Объяснение механизма возникновения и проведения возбуждения стало возможным после создания С. Аррениусом теории электролитической диссоциации. В соответствии с этой теорией Бернштейн (J. Bernstein) предположил, что возникновение и проведение Н. и. обусловлено перемещением ионов между нервным волокном и окружающей средой. Англ. исследователи А. Ходжкин, Б. Катц и Э. Хаксли детально исследовали трансмембранные ионные токи, лежащие в основе развития потенциала действия. Позже стали интенсивно изучаться механизмы работы ионных каналов, по к-рым происходит обмен ионами между аксоном и окружающей средой, и механизмы, обеспечивающие способность нервных волокон проводить ряды Н. и. разного ритма и продолжительности.
Н. и. распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками нервного волокна. Ток, выходящий из волокна наружу в покоящемся участке, служит раздражителем. Наступающая после возбуждения в данном участке нервного волокна рефрактерность обусловливает поступательное движение Н. и.
Количественно соотношения разных фаз развития потенциала действия можно охарактеризовать, сопоставляя их по амплитуде и длительности во времени. Так, напр., для миелиновых нервных волокон группы А млекопитающих диаметр волокна находится в пределах 1—22 мк, скорость проведения — 5—120 м/сек, длительность и амплитуда высоковольтной части (пика, или спайка) — 0,4—0,5 мсек и 100—120 мв соответственно, следовой негативный потенциал — 12—20 мсек (3—5% от амплитуды спайка), следовой позитивный потенциал — 40—60 мсек (0,2% от амплитуды спайка).
Возможности передачи разнообразной информации расширяются за счет повышения скорости развития потенциала действия, скорости распространения, а также за счет повышения лабильности (см.) — т. е. способности возбудимого образования воспроизводить в единицу времени высокие ритмы возбуждения.
Возникновение Н. и. в нервных клетках (см.) или рецепторах (см.) связано с деполяризацией мембраны, т. е. со снижением величины электрического потенциала на мембране (потенциала покоя, или мембранного потенциала). Если величина мембранного потенциала снижается на 10—20% (пороговый критический уровень), то местный процесс переходит в распространяющийся — возникает потенциал действия (см. Возбуждение).
Конкретные особенности распространения Н. и. связаны со строением нервных волокон (см.). Сердцевина волокна (аксоплазма) обладает низким сопротивлением и, соответственно, хорошей проводимостью, а окружающая аксоплазму плазматическая мембрана — большим сопротивлением. Особенно велико электрическое сопротивление наружного слоя у миелинизированных волокон, у к-рых свободны от толстой миелиновой оболочки только перехваты Ранвье. В безмиелиновых волокнах Н. и. движется непрерывно, а в миелиновых — скачкообразно (сальтаторное проведение).
Различают декрементное и бездекрементное распространение волны возбуждения. Декрементное проведение, т. е. проведение возбуждения с угасанием, наблюдается в безмиелиновых волокнах. В таких волокнах скорость проведения Н. и. невелика и по мере отдаления от места раздражения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания. Декрементное проведение свойственно волокнам, иннервирующим внутренние органы, обладающие низкой функц, подвижностью. Без декрементное проведение характерно для миелиновых и тех безмиелиновых волокон, к-рые передают сигналы к органам, обладающим высокой реактивностью (напр., сердечной мышце). При бездекрементном проведении Н. и. проходит весь путь от места раздражения до места реализации информации без затухания.
Передача Н. и. с нервного волокна на мышечное или какой-либо другой эффектор осуществляется через синапсы (см.). У позвоночных животных в подавляющем большинстве случаев передача возбуждения на эффектор происходит при помощи выделения ацетилхолина (нервно-мышечные синапсы скелетной мускулатуры, синаптические соединения в сердце и др.). Для таких синапсов характерно строго одностороннее проведение импульса и наличие временной задержки передачи возбуждения.
В синапсах, в синаптической щели которых сопротивление электрическому току благодаря большой площади контактирующих поверхностей мало, происходит электрическая передача возбуждения. В них нет синаптической задержки проведения и возможно двустороннее проведение. Такие синапсы свойственны беспозвоночным животным.
Регистрация Н. и. нашла широкое применение в биол, исследованиях и клин, практике. Для регистрации используют шлейфные и чаще катодные осциллографы (см. Осциллография). При помощи микроэлектродной техники (см. Микроэлектродный метод исследования) регистрируют Н. и. в одиночных возбудимых образованиях — нейронах и аксонах. Возможности исследования механизма возникновения и распространения Н. и. значительно расширились после разработки метода фиксации потенциала. Этим методом были получены основные данные о ионных токах (см. Биоэлектрические потенциалы).
Нарушение проведения Н. и. происходит при повреждении нервных стволов, напр, при механических травмах, сдавливании в результате разрастания опухоли или при воспалительных процессах. Такие нарушения проведения Н. и. зачастую бывают необратимы. Следствием прекращения иннервации могут быть тяжелые функциональные и трофические расстройства (напр., атрофия скелетных мышц конечностей после прекращения поступления Н. и. вследствие необратимой травмы нервного ствола). Обратимое прекращение проведения Н. и. может быть вызвано специально, в терапевтических целях. Напр., с помощью анестезирующих средств блокируют импульсацию, идущую от болевых рецепторов в ц. н. с. Обратимое прекращение проведения Н. и. вызывает и новокаиновая блокада. Временное прекращение передачи Н. и. по нервным проводникам наблюдается и во время общего наркоза.
Библиография: Бpеже М. А. Электрическая активность нервной системы, пер. с англ., М., 1979; Жуков Е. К. Очерки по нервно-мышечной физиологии, Л., 1969; Коннели К. Восстановительные процессы и обмен веществ в нерве, в кн.: Совр, пробл. биофизики, пер. с англ., под ред. Г. М. Франка и А. Г. Пасынского, т. 2, с. 211, М., 1961; Костюк П. Г. Физиология центральной нервной системы, Киев, 1977; Латманизова Л. В. Очерк физиологии возбуждения, М., 1972; Общая физиология нервной системы, под ред. П. Г. Костюка, Л., 1979; Тасаки И. Нервное возбуждение, пер. с англ., М., 1971; Ходжкин А. Нервный импульс, пер. с англ., М., 1965; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975.
Проведение нервных импульсов осуществляется при помощи отростков нервных клеток – нервных волокон. По особенностям строения и расположения осевых цилиндров в составе нерва (по отношению к шванновским клеткам) различают волокна мякот-ного (миелинизированные) и безмякотного типа. Как правило, нервы включают волокна обоих типов в разном количественном соотношении. Миелиновые волокна преобладают в чувствительных и двигательных нервах органов чувств, скелетных мышц и вегетативной нервной системы. Безмиелиновые преимущественно встречаются в составе нервов симпатического отдела.
Волокна мякотного и безмякотного типов состоят из одинаковых структурных компонентов: отростка нервной клетки (осевого цилиндра), шванновских клеток (леммоцитов) и базальной мембраны (тонкой пластинки, состоящей из рыхлой волокнистой соединительной ткани и покрывающей каждое нервное волокно снаружи). Миелиновое волокно представлено осевым цилиндром, окруженным миелиновой оболочкой. Шванновские клетки, окружающие отросток нейроцита, образуют дупликатуры цитоплазма-тической мембраны, называемые мезаксонами. Многократно обертываясь вокруг осевого цилиндра, мезаксон формирует миелиновую оболочку нервного волокна, представляющую собой плотный липидно-белковый футляр. На протяжении мякотного нервного волокна через примерно равные интервалы имеются участки истончения миелинового слоя, ширина которых составляет около 1 мкм. Эти участки получили название перехватов Ранвье. Они имеют большое функциональное значение при проведении нервного импульса. Расстояние от одного до другого перехвата называется межузловым сегментом. Мякотные волокна могут располагаться как в пределах центральной нервной системы, так и вне ее. В зависимости от этого среди них различают центральные и периферические. Миелиновые волокна характеризуются высокой скоростью проведения импульсов (60-120 м/с). Эта особенность обусловлена изолирующей функцией миелино-вого слоя и наличием перехватов Ранвье.
Безмякотные нервные волокна построены более примитивно. Они представлены цепочкой шванновских клеток, в которую вдавлено от одного и более осевых цилиндров. В среднем их количество составляет от 5 до 20. Миелинового слоя в этих клетках нет, леммоциты образуют между осевыми цилиндрами мезаксо-ны, не играющие существенной роли в проведении нервного возбуждения. Отростки нейроцитов расположены довольно рыхло и плохо изолированы. Поэтому они могут переходить из одного безмиелинового волокна в другое. Снаружи волокно покрыто соединительно-тканной базальной пластинкой. В связи с перечисленными особенностями строения безмякотные нервные волокна называют волокнами кабельного типа. Скорость проведения в них (3–5 м/с) гораздо ниже, чем в миелиновых.
Функцией нервного волокна является проведение нервных импульсов, способ которого в мякотных и безмякотных нервных волокнах несколько различается. В этом процессе большое значение имеют структуры нервного волокна.
Главный элемент с функциональной точки зрения – цито-плазматическая мембрана осевого цилиндра. Именно на мембране возникает нервный импульс, а затем распространяется по ней. Миелин одновременно служит изолятором и выполняет трофическую функцию по отношению к осевому цилиндру. Являясь веществом липидно-белковой природы, миелин благодаря своему высокому сопротивлению ограничивает передвижение ионов. В связи с этим потенциал действия, возникновение которого связано с перераспределением ионов, может возникать не на любом участке волокна, а в местах наименьшего сопротивления – истончениях миелина, перехватах Ранвье. Это имеет большое значение в проведении нервного возбуждения и обуславливает разницу механизмов проведения в мякотных и безмякотных волокнах. Трофическая функция миелиновой оболочки заключается в регуляции роста и обмена веществ осевого цилиндра. Кроме того, шванновские клетки при повреждении нерва осуществляют дегенерацию дистальной от тела клетки отсеченной части осевого цилиндра, а затем образуют ложе для нового волокна, обеспечивая направление продвижения колбы роста.
Свою роль в распространении импульса играют и внутриклеточные структуры осевого цилиндра, в частности нейрофибрил-лы, микротубулы и транспортные филаменты, соединяющиеся с различными веществами или органеллами и осуществляющие их транспорт по нервным волокнам.
Проведение импульса протекает при определенных условиях и подчиняется ряду законов. Во-первых, проведение нервного импульса по волокну может осуществляться полноценно лишь при условии его анатомической и физиологической целостности, или непрерывности. Анатомическая целостность может нарушаться при механических повреждениях, например при перерезке или сдавливании волокна, нарушение физиологической целостности может быть результатом сильного охлаждения, действия веществ – блокаторов натриевых каналов мембраны (местных анестетиков) или других подобных факторов. Все это вызывает частичное или полное нарушение проводимости нервного волокна.
С помощью многочисленных опытов (А. И. Бабухин, 1877 г., В. Кюне, 1886 г.) было доказано, что возникшее на мембране возбуждение распространяется в двух направлениях: в центробежном и в центростремительном. Потенциал действия возникает на нейроне в так называемой триггерной зоне (в месте перехода тела нервной клетки в нейрит), и распространяется по аксону и по телу к дендритам. В условиях целостного организма наблюдать двустороннее проведение нельзя из-за наличия клапанного аппарата синаптических соединений, передающих импульс только в одном направлении, и специфики места возникновения возбуждения (рецепторного аппарата).
В одном нерве могут находиться волокна разных видов – двигательные, чувствительные, вегетативные – различающиеся иннервируемыми структурами. В связи с этим очень важным свойством является изолированное проведение возбуждения в нервных волокнах. В первую очередь, роль изолятора выполняет миелиновая оболочка, обладающая высоким сопротивлением. Большое значение имеет также наличие жидкости в межклеточных пространствах. За счет более низкого ее сопротивления по сравнению с мембраной волокна ток между деполяризованным и покоящимся участками мембраны идет по межклеточным щелям и не затрагивает другие волокна.
Таким образом, проведение нервного импульса подчиняется трем основным законам: закону анатомической и физиологической целостности, двустороннего проведения и изолированного проведения возбуждения.
Возникновение его начинается со снятия потенциала покоя. Потенциалом покоя, или мембранным потенциалом, называют разницу потенциалов между наружной (положительно заряженной) и внутренней (отрицательно заряженной) поверхностями клеточной мембраны в состоянии относительного физиологического покоя. Эта величина составляет примерно 60–90 мВ.
В соответствии с мембранно-ионной теорией, предложенной в 1902 г. немецким физиологом Н. А. Берштейном, в поддержании постоянства потенциала покоя участвует ряд факторов.
Среди них называют в первую очередь особенности строения и функционирования клеточных мембран. Разность потенциалов существует за счет различного содержания ионов натрия, калия, кальция и хлора в клетке и вне ее и, следовательно, различной проницаемости мембраны для них. Важным свойством цитоплаз-матической мембраны является наличие в ней белков (60 %); среди них различают периферические, погруженные (полуинтегральные) и трансмембранные (интегральные), которые помимо рецепторной и ферментативной функции выполняют транспортную функцию и образуют поры, предназначенные для перемещения определенных ионов.
Поры, или каналы, имеют диаметр около 1 нм, образованы молекулами белков и работают в соответствии с воротным механизмом, регуляция которого обеспечивается разностью потенциалов или взаимодействием белка-рецептора с адекватным медиатором.
В соответствии с последним, различают поры электровозбудимые (преобладают в нервных и мышечных клетках) и хемовозбу-димые. Функционирование воротного механизма представляет собой последовательную смену состояний канала. В закрытом состоянии, которое существует в относительном физиологическом покое, пора не может пропустить ион. Из закрытого состояния канал переходит в открытое, соответствующее началу деполяризации. Это, собственно, время работы поры. После открытого следует состояние инактивации, когда канал все еще открыт, но ион не пропускает. После этого канал вновь закрывается.
Поры могут осуществлять транспорт только определенного иона, таким образом, обладая специфичностью (избирательностью), что объясняется разницей или сходством диаметра иона и канала, зарядом внутри него и вышеописанным воротным механизмом.
В покое клеточная мембрана обладает хорошей проницаемостью для ионов калия и хлора. Проницаемость для натрия гораздо ниже. Анионы и молекулы органических веществ не могут проникнуть через мембрану.
Следующим фактором, обусловливающим наличие и постоянство величины мембранного потенциала, является ионная асимметрия внутри клетки и снаружи. В клетке содержатся преимущественно ионы калия и анионы органических веществ, а вне ее больше ионов натрия, хлора и кальция. Ионную асимметрию можно считать главной причиной существования мембранного потенциала. Она поддерживается за счет избирательной проницаемости цитоплазматической мембраны и физико-химического равновесия Доннана.
Большое значение в поддержании мембранного потенциала имеют так называемые пассивные силы, объединяющие силы простой диффузии и электростатического взаимодействия ионов. Пассивными их называют потому, что для своей реализации они не требуют энергетических затрат. Действие этих сил можно проследить, например, на ионе калия. Клеточная мембрана хорошо проницаема для калия, в клетке содержание калия больше, чем вне ее. По градиенту концентрации калий выходит из клетки. Он заряжен положительно, следовательно, выходя на поверхность, он попадает под действие сил электростатического отталкивания, так как в состоянии относительного физиологического покоя наружная мембрана также заряжена положительно, и снова возвращается в клетку. Таким образом, калий находится преимущественно внутри нее.
Для хлора, находящегося большей частью снаружи, клеточная мембрана тоже легко проницаема. Аналогично калию, на хлор действуют силы простой диффузии, под воздействием которых он устремляется в клетку. Отрицательный заряд внутренней поверхности мембраны отталкивает анионы, и хлор остается снаружи.
Клеточная мембрана не пропускает ионы натрия, находящегося вне клетки, и анионы органических веществ (внутри), они не могут пройти через нее при помощи диффузии и соответственно, остаются за пределами клетки и в ней.
Фактически мембранный потенциал основывается на электрохимическом равновесии по иону калия (количество калия, вышедшего из клетки благодаря диффузии и вернувшегося путем электростатического отталкивания, одинаково). Это равновесие развивается при условии, если создается равенство сил диффузии и отталкивания в клетке.
Важным звеном в формировании и поддержании мембранного потенциала является работа натрий-калиевого насоса, активного транспорта ионов через цитоплазматическую мембрану против градиента концентрации с затратой энергии. Функционирование насоса происходит при участии натрий-калиевой АТФ-азы, которая за один цикл работы насоса выводит за пределы клетки три иона натрия и возвращает два калия.
При возникновении потенциала действия происходит снятие мембранного потенциала и перезарядка мембраны, что является результатом постепенного изменения проницаемости для определенных ионов. Он способен генерироваться только в определенных участках мембраны нервной клетки, называемых электровозбудимыми. Они располагаются в аксональном холмике (триггерная зона), нейрите, иногда в теле (но не на постсинапти-ческих мембранах), в перехватах Ранвье. Наиболее значимая из этих структур – аксональный холмик. Потенциал действия возникает лишь при достижении критического уровня деполяризации (влияние распространяющегося возбуждающего постсинап-тического потенциала из синапсов или рецепторного потенциала с чувствительных нейронов). Мембрана аксонального холмика содержит большое количество натриевых и калиевых каналов и, кроме того, имеет сравнительно низкий с другими структурами порог возбуждения. Это обусловливает более легкое достижение критического уровня деполяризации. Таким образом, этот участок является наиболее легко возбудимым в клетке.
С точки зрения мембранно-ионной теории, потенциал действия развивается благодаря повышению мембранной проницаемости для двух видов ионов – натрия и калия. Сначала открываются натриевые каналы, проницаемость увеличивается в 400–500 раз, и натрий устремляется в клетку. Для калия проницаемость становится возможна позже, к началу реполяризации, и увеличивается всего в 10–15 раз. В итоге в процессе формирования потенциала действия натрия выходит гораздо больше, чем выходит калия.
Потенциал действия – достаточно сложное комплексное явление, включающее несколько фаз. В первую очередь возникает локальный ответ, затем следует пик, или спайк, состоящий из фазы деполяризации и реполяризации, позже наблюдаются следовые потенциалы – отрицательный и положительный.
Локальный ответ (начальная фаза) возникает, когда сила действующего раздражителя еще не достигла пороговой (подпорого-вый раздражитель). Этот этап характеризуется увеличением ионной проницаемости мембраны для всех ионов (неспецифически). Мембранный потенциал начинает смещаться в положительную сторону. В процессе увеличения силы воздействия при достижении 50–70 % от порога уже имеет место специфическое увеличение проницаемости для натрия, который силами простой диффузии входит в клетку и частично деполяризует мембрану. Мембранный потенциал стремится к критическому уровню деполяризации, который в разных клетках может составлять примерно от 50 до 40 мВ
При достижении раздражителем пороговой силы начинается первый этап фазы спайка: деполяризация (восходящее колено). Мембранный потенциал достигает критического уровня, что вызывает открытие всех натриевых каналов и лавинообразный ток ионов внутрь клетки. Это становится причиной увеличения мембранного потенциала до нуля, а затем и перезарядки мембраны. Деполяризация прекращается после развития нового электрохимического равновесия по натрию и инактивации натриевых каналов.
В процессе реполяризации (нисходящее колено) мембрана начинает пропускать ионы калия. Он покидает клетку, удаляя таким образом положительный заряд. Перераспределение ионов активирует работу натрий-калиевого насоса.
За реполяризацией следует отрицательный следовой потенциал. Он характеризуется повторным незначительным увеличением проницаемости для натрия, который в небольшом количестве входит в клетку и вызывает приближение мембранного потенциала к критическому уровню – частичную деполяризацию. Эта фаза сменяется положительным следовым потенциалом, основным явлением которого называют следовую гиперполяризацию (формирование положительного заряда на наружной поверхности мембраны, возникающее в результате продолжения тока калия из клетки). В итоге величина мембранного потенциала вновь отдаляется от критического уровня.
Таким образом, возбуждение включает локальный ответ, который не распространяется и быстро затухает, и волновой ответ, распространяющийся далее по волокнам.
Осуществляясь в соответствии с одинаковыми закономерностями, процессы проведения в миелиновых и безмиелиновых волокнах отличается рядом особенностей. В безмиелиновых волокнах импульс распространяется от возбужденного участка к соседним, расположенным рядом с ним. Распространение потенциала действия происходит в обе стороны без изменения амплитуды с помощью круговых токов.
Различная скорость проведения, продолжительность фаз потенциала действия, строение волокон позволяет подразделять их на 3 вида: А, В, С.
Волокна типа А – миелиновые, они, в свою очередь, подразделяются еще на 4 подгруппы: α-, β-, γ– и δ-волокна. Альфа-волокна, или, как их еще называют, А-а, среди этой группы имеют самый большой диаметр (12–22 мкм) и высокую скорость проведения импульса (70-120 м/с). В организме они достаточно распространены. К этой группе относятся двигательные и чувствительные волокна скелетной мускулатуры. Что касается длительности различных фаз потенциала действия, то продолжительность пика составляет 0,4–0,5 мс, следовой деполяризации – 15–20 мс, следовой гиперполяризации – 40–60 мс. Следующие подгруппы типа А – А-β, А-γ, А-δ – отличаются меньшим диаметром и скоростью проведения возбуждения, но их потенциал действия более продолжителен, чем А-α. Эти волокна участвуют в основном в проведении импульсов от различных рецепторов внутренних органов в нервные центры. А-β волокна идут от тактильных рецепторов, А-γ – от тактильных и барорецепторов, а также к мышечным веретенам как двигательные волокна. В А-δ подгруппу входят афферентные волокна, несущие импульс от термо-, барорецепторов и ноцицепторов. Проведение в трех последних подгруппах составляет 1-12 м/с. Самое быстрое проведение – в А-β волокнах. Пик потенциала действия может продолжаться до 0,9–1,0 мс, (наиболее длительное – в А-β волокнах). Следовая деполяризация и гиперполяризация продолжаются примерно в течение того же времени, что и в А-α.
Волокна типа В также покрыты миелиновой оболочкой, имеют достаточно низкую скорость проведения и находятся в составе вегетативных отделов нервной системы. В основном, это преганг-лионарные вегетативные волокна. Проведение осуществляется со скоростью от 3 до 18 м/с. Наряду с этим, в В-волокнах потенциал действия по длительности превосходит более чем в 3 раза это значение в А-волокнах. Важнейшим отличием является тот факт, что в этих волокнах не наблюдается фаза следовой гиперполяризации. Следовая гиперполяризация может продолжаться до 100 мс.
Волокна типа С по строению безмиелиновые, очень тонкие, всего 0,5–2,0 мкм в диаметре. Они также обнаруживаются в вегетативной нервной системе в составе постганглионарных волокон, осуществляя проведение от рецепторов тепла, холода, давления и боли. Эти волокна отличаются наиболее медленной скоростью проведения (не больше 3 м/с). Их потенциал действия отличается самой большой (по сравнению с другими типами) длительностью развития потенциала действия: у теплокровных животных и человека – до 2 мс.
Волокна способны существовать и выполнять свою функцию только тогда, когда они связаны с телом нейрона. Повреждение волокна ведет к нарушению или утрате способности проводить возбуждение. Перерезка ведет к гибели отсеченной части волокна. Однако волокна способны регенерировать за счет гипертрофии (эндорепродукции). Разрушение отделенного волокна и образование нового является сложным и длительным процессом и протекает в несколько последовательных стадий. После перерезки миелиновый слой перерождается в жировые капли. Леммо-циты осуществляют дегенерацию осевого цилиндра с помощью гидролитических ферментов лизосом. В результате на месте волокна остается цепь леммоцитов. Дальше начинается регенерация волокна. От места его отделение в ложе, образованной шваннов-скими клетками, начинает прорастать колба роста. Регенерация осуществляется с примерной скоростью 0,5–4,5 мм за сутки в зависимости от строения волокна и местоположения его в организме.
Читайте также:
