
Рефлекс кожи нервная система
Кожные анализаторы делят на анализаторы, рецепторы которых возбуждаются болевым раздражением, и анализаторы, рецепторы которых возбуждаются прикосновением (тактильная чувствительность). В соответствии с этим в клинике различают две основные группы кожных рефлексов спинного мозга: 1) болевые рефлексы с кожи, 2) рефлексы, вызываемые тактильным раздражением кожи.
Болевые рефлексы спинного мозга . Эти рефлексы играют большую роль в деятельности нервной системы, так как сигнализируют о начавшемся повреждении целостности тканей. В зависимости от интенсивности, обширности и топографии повреждения болевой кожный рефлекс чрезвычайно разнообразен. Самый простой болевой рефлекс можно вызвать уколом иглы. В ответ на это болевое раздражение происходит движение, удаляющее часть тела от источника раздражения. Этот рефлекс особенно четко выявляется в случаях, когда спинной мозг освобождается от влияния на него со стороны головного, как, например, у больных, у которых данная связь (главным образом пирамидные пути) нарушаются при патологических процессах в верхних отделах спинного мозга. При этом наблюдается так называемый защитный рефлекс: при уколе в области стопы больной рефлекторно отдергивает ногу тройным сгибанием в голеностопном, коленном и тазобедренном суставах.
Особое место занимают болевые рефлексы при раздражении рецепторов глубокой болевой чувствительности. Повреждение костей, особенно надкостницы, связочного аппарата и суставов, вызывает сильное раздражение заложенных в них рецепторов глубокой болевой чувствительности. Это раздражение вызывает анталгический рефлекс. Так, если возникает патологический процесс в каком-либо сочленении позвонков с одной стороны, то появляется рефлекторное тоническое искривление позвоночника в противоположную сторону, разгружающее заболевший сустав (анталгический сколиоз). При раздражении задних корешков спинного мозга возникает рефлекторное сопротивление натяжению корешков. На этом основан ряд так называемых оболочечных или корешковых симптомов натяжения, или болевых тонических рефлексов.
Симптом Кернига . Если у лежащего на спине больного согнуть ногу под прямым углом в тазобедренном и коленном суставах, то попытка разогнуть ее в коленном суставе не удается из-за рефлекторного сокращения сгибателей голени.
Симптом посадки . Сидящему в кровати больному не удается полностью разогнуть ноги в коленных суставах, так как наступающее вскоре рефлекторное сокращение сгибателей голени препятствует движению.
Симптом Брудзинского . При энергичном наклоне головы у лежащего на спине больного ноги сгибаются в коленных суставах и приводятся к животу (верхний симптом Брудзинского). Если у больного пассивно сгибать одну ногу в коленном и тазобедренном суставах, вторая нога тоже сгибается (нижний симптом Брудзинского). Сюда же следует отнести лобковый симптом Брудзинского (при надавливании на лобок ноги больного приводятся и сгибаются в тазобедренных и коленных суставах), а также симптом Гийена (сжатие рукой исследователя четырехглавой мышцы вызывает ее сокращение на другой ноге). Ригидность затылочных мышц и длинных мышц спины при заболевании оболочек и корешков спинного мозга также относится к болевым тоническим рефлексам в ответ на раздражение рецепторов глубокой чувствительности.
Рефлексы спинного мозга, вызываемые тактильным раздражением кожи . Кожные рефлексы спинного мозга вызываются штриховым раздражением кожи, в ответ на которое происходит сокращение той или иной мышцы или группы их. В отличие от сухожильных рефлексов кожные не являются врожденными. Они возникают у детей в разном возрасте (от 5 месяцев до 3 лет). Очевидно, их формирование в большой степени обусловлено развитием коры полушария головного мозга и пирамидных путей. Двойное замыкание кожных рефлексов (в спинном мозгу и коре головного мозга) обусловливается тем, что их отсутствие может быть вызвано поражением как их спинальной рефлекторной дуги, так и пирамидного пути, который является существенным звеном в эфферентной части дуги кожных рефлексов.
К кожным рефлексам относятся следующие.
Брюшные рефлексы . Они вызываются быстрым штриховым раздражением кожи живота тупым концом иглы или рукояткой молоточка. Ответная реакция состоит в сокращении мышц живота на одноименной стороне. Для вызывания верхнего брюшного рефлекса штриховое раздражение наносится на коже параллельно реберной дуге, для среднего брюшного рефлекса - на уровне пупка в горизонтальном направлении, для нижнего брюшного рефлекса - параллельно паховой складке.
Сюда же относится описанный В. М. Бехтеревым костно-абдоминальный рефлекс, заключающийся в том, что при ударе молотком по краю реберной дуги кнутри от сосковой линии сокращаются брюшные мышцы соответствующей стороны. Это глубокие (периостальные) брюшные рефлексы могут быть использованы для сравнения брюшных рефлексов той и другой стороны.
Рефлекс кремастерной мышцы вызывается при нанесении штрихового раздражения на кожу внутренней поверхности бедра на 1-2 см ниже паховой складки. Ответная реакция при этом выражается в подтягивании яичка кверху.
Подошвенный рефлекс вызывается штриховым раздражением подошвы, в ответ на которое следует подошвенное сгибание пальцев.
Анальный рефлекс вызывается уколом кожи около заднего прохода. В ответ сокращается его круговая мышца.
Особое место занимают так называемые суставные рефлексы. По своей сущности они относятся к глубоким рефлексам, однако с кожными рефлексами их сближает позднее появление и зависимость от пирамидных путей. При целости спинальных рефлекторных дуг суставных рефлексов ослабление или исчезновение их расценивается как признак поражения пирамидных путей. К ним относятся следующие рефлексы.
Рефлекс Майера . Вызывается форсированным сгибанием основной фаланги III или IV пальца супинированной кисти руки. При этом происходит сгибание основной, а также приведение и разгибание ногтевой фаланги большого пальца.
Рефлекс Лери . В положении супинированной кисти и согнутых пальцев производится энергичное сгибание пальцев и кисти в лучезапястном суставе. При этом наблюдается рефлекторное сгибание руки в локтевом суставе.
Понижение или отсутствие кожных и суставных рефлексов в сочетании с повышением сухожильных рефлексов и появлением патологических является достоверным признаком поражения пирамидных путей. Следует иметь в виду, что брюшные рефлексы нередко не вызываются при наличии у больного дряблой брюшной стенки при полной интактности рефлекторных дуг этих рефлексов.
Повышение кожных рефлексов не играет в клинике такой роли, как их понижение или отсутствие. Часто обнаруживается повышение брюшных и подошвенных рефлексов при функциональных заболеваниях нервной системы, при общем повышении ее возбудимости. Обычно у этих больных само исследование рефлексов вызывает общую эмоциональную реакцию (вздрагивание всего тела, вскрикивание и др.).
Некоторое значение в клинике имеет повышение суставного рефлекса Майера. Оно проявляется тем, что большой палец приводится и оппонируется при самом легком сгибании основных фаланг III и IV пальцев, а также тем, что при этом вызываются дополнительные сокращения сгибателей предплечья и дельтовидной мышцы. Повышение рефлекса наблюдается иногда при лобных локализациях процесса, причем на одноименной очагу стороне. Нередко повышение рефлекса Майера сопутствует хватательному рефлексу.
Рефлексы, как суммарное выражение интегральной рефлекторной деятельности, проявляются у животных всех видов, но при заболевании нервной системы они могут резко изменяться, задерживаться или даже полностью выпадать в зависимости от характера патологического процесса.
Например, разрушение нервных волокон периферического нерва вызывает выпадение рефлекторной деятельности той части конечности, которая иннервируется данным моторным нервом. Причины расстройства рефлекторной деятельности могут быть весьма различными: ушибы периферических нервов, сотрясение головного и спинного мозга, авитаминозы, пищевые интоксикации, лучевые поражения, отравления и др. Характер нарушения рефлексов существенно изменяется в зависимости от этиологического фактора, в то же время рефлекс сохраняет известное функционально-морфологическое единство, что следует объяснить длительной приспособительной реакцией животных, обусловившей представительство того или иного органа в коре полушарий мозга в виде особого нервного центра. Павловский принцип динамической локализации функций разрушил узколокалистический и статистический принципы органной локализации тех же функции (И. В. Давыдовский), поэтому в настоящее время нельзя говорить только о рефлекторной дуге. Схематично сущность рефлекса заключается в восприятии раздражений специальными образованиями нервной системы — рецепторами и в распространении волны возбуждения с афферентных на эфферентные, или двигательные, ней ионы, затем на реагирующий орган. С морфологической сто ролы рефлекс представляет цепь двух нейронов или многих нейронов, по которой распространяется возбуждение, причем в невропатологии имеют большое значение не только анимальные, но и вегетативные рефлексы.
К настоящему времени учение о рефлексе обогатилось понятием обратной связи, заключающейся в том, что, как только совершилось действие в каком-либо органе, от него в центральную нервную систему поступают от проприорецепторов центростремительные импульсы, сигнализирующие о его состоянии. В соответствующих центрах (клетки Реншоу) сличается то, что должно было быть и что произошло. В результате такой обратной афферентации центр посылает исправляющие сигналы к данному органу; эти сигналы могут повысить или понизить количество функционирующих рецепторов и даже изменить их чувствительность.
По принципу обратной связи не только осуществляются сложные реакции поведения животных, но также поддерживается постоянство температуры тела, кровяного давления, уровня сахара и др. Рефлекс зрачка может служить примером функционирующей обратной связи: яркий свет вызывает сокращение сфинктера зрачка, в то время как рассеянный свет или затемнение, напротив, расширяют зрачок. Сужение зрачка происходит до тех пор, пока величина сигнала от палочек и колбочек светочувствительного слоя сетчатки не достигнет оптимального значения. Следовательно, благодаря обратной связи центральная нервная система информируется о качественной стороне производимого действия и принимается новая команда, достигающая наибольшей точности и эффективности в соответствии с условиями дайной конкретной обстановки.
С точки зрения кибернетики в структуре нервных клеток и их отростков, образующих субстрат рефлекторной дуги, заложена программа прохождения нервного импульса. Эта программа может изменяться в зависимости от времени и внешних воздействий, поэтому патологическое состояние изменяет программу действий, т. е. изменяется и сам рефлекс.
Используемые в клинике рефлексы могут быть разделены на поверхностные, глубокие и висцеральные. К поверхностным относят кожные рефлексы, возникающие в результате раздражения рецепторов кожи, к глубоким — глубжележащих тканей — сухожилий, мышц, надкостницы. Висцеральными называют рефлексы с внутренних органов, причем многие из них имеют проекции на кожу.
У крупного рогатого скота, лошадей, собак и других животных исследуют корнеальный, или роговичный, рефлекс. Для вызывания его тонким кусочком ваты или бумаги дотрагиваются до роговицы. В ответ на такое прикосновение происходит мигание или смыкание век. Оно возникает в результате раздражения глазной ветви тройничного нерва и обычно исчезает при поражении бульбарной части мозга.
Брюшные рефлексы вызывают штриховым раздражением кожи брюшной стенки, в ответ на которое наблюдается сокращение брюшных мускулов. Ослабление этого рефлекса или даже полное исчезновение связано с перерывом спинальной рефлекторной дуги или поражением пирамидного пути. Следует иметь в виду, что брюшные рефлексы могут исчезать при острых заболеваниях области брюшной полости (метеоризм рубца, воспаление брюшины). У собак мы наблюдали резкое ослабление брюшных рефлексов при огнестрельном ранении дробью спинного мозга на уровне и поясничного позвонка, а также при переломах позвоночника и субдуральной гематоме, появившейся после ушиба. Полагают, что брюшные рефлексы связаны с функциями высших отделов нервной системы и появляются в процессе фило — и онтогенеза сравнительно поздно, поэтому их нет у обезьян и новорожденных животных.
Рефлекс спины вызывается надавливанием пальцами на область поясницы или пощипыванием кожи по ходу сагиттальной линии позвоночника. В ответ на такое раздражение происходит резкое прогибание позвоночника и нередко движение хвостом. Исчезновение рефлекса наблюдается в тех же случаях, что и брюшных рефлексов. В дифференциально-диагностическом отношении нужно иметь в виду острое воспаление почек, так как при нем животные тоже прогибают спину при надавливании на область поясницы в передней ее трети.
Перинеальный рефлекс вызывают раздражением кожи промежности. В ответ на такое раздражение происходит толчкообразное подтягивание корня хвоста, сокращение кругового мускула анального отверстия.
Анальный рефлекс вызывают уколом или штриховым раздражением кожи вблизи заднего прохода или введением в него пальца. В результате такого раздражения происходит выраженное сокращение наружного сфинктера. Н. Ф. Попов наблюдал сохранение анального рефлекса даже в тех случаях, когда у собаки оперативным путем выталкивали спинной мозг. В. Элленбергер (1930) сообщил о сохранении этого рефлекса у коров после глубокого оглушения и обескровливания. Приведенные факты свидетельствуют об исключительной стойкости анально-перинеальных рефлексов. В связи с этим небезынтересно отметить, что восстановление функций и оживление человека (утопленника) иногда достигается уколом именно в области промежности, несколько ниже ануса (чжень-дзю терапия).
Рефлекс кремастера заключается в подтягивании семенника соответствующей стороны при раздражении или уколе кожи на внутренней поверхности бедра той же стороны. Он может отсутствовать при параличах срамного нерва, повреждениях спинного мозга и его оболочек. И. Марек сообщил, что у жеребцов следует раздражать кожу в области внутреннего мыщелка бедренной кости.
На тазовой конечности у собак легко вызывается подошвенный рефлекс, если покалывать булавкой кожу между пальцами. В ответ на такое раздражение происходит разгибание пальцев. Этот рефлекс плохо выражен или отсутствует при послечумных миелитах, субдуральных гематомах спинного мозга. У лошадей отдергивание конечности и сокращение мышц предплечья или голени получается всякий раз, когда сдавливается подошва копытными щипцами, особенно в случаях развития острого пододерматита. После расчистки подошвы можно получить рефлекторное отдергивание конечности, если пальцем надавливать на область подошвы, стрелки, венчика. Этот рефлекс у лошади называют также рефлексом копытной кости и относят к глубоким рефлексам. Его клинико-диагностическое значение связано с возможным выпадением функции двигательной сферы коры, полосатого тела и пирамидного пути, что можно наблюдать при столбняке и энцефаломиелите. При параличах лучевого и седалищного нервов этот рефлекс резко ослабевает.
Из других глубоких рефлексов практическое значение имеют коленный рефлекс и рефлекс ахиллова сухожилия.
Эти рефлексы относятся к так называемым проприорефлексам, так как они вызываются раздражением проприорецепторов мышц, сухожилий, надкостницы или сустава.
Коленный рефлекс вызывается ударом перкуссионного молоточка или краем ладони несколько ниже коленной чашечки, по прямой связке ее. Манипуляции проводят на лежачем животном или на приподнятой конечности, слегка согнутой в коленном суставе. У лошадей и крупного рогатого скота необходимо слегка приподнять конечность и добиться расслабления мускулатуры. В ответ на периодическое постукивание молоточком происходит разгибательное движение коленного сустава, Отсутствие этого рефлекса указывает на нарушение проводимости в сером веществе II—IV поясничных сегментов или в соответствующих корешках спинного мозга. Поражение бедренного нерва (параличи различной этиологии) также сопровождается выпадением или резким ослаблением коленного рефлекса, поэтому его необходимо исследовать при подозрении на поражение бедренного нерва.
Ахиллов рефлекс проявляется относительно слабым разгибанием скакательного сустава при одновременном сгибании путового и венечного суставов в момент ударов по ахиллову сухожилию. Чтобы вызвать рефлекс, нужно поднять конечность и удерживать в слегка отведенном кзади положении (как при ковке), добиваясь расслабления мускулатуры, затем нанести короткий удар по ахиллову сухожилию на 10—15 см выше пяточного бугра. Если разгибательное движение не вызывается или выражено очень слабо, то можно подозревать нарушение рефлекторной связи в сером веществе на уровне последних поясничных сегментов и I—II крестцовых, т. е. в том мосте, где формируется седалищный нерв.
Коленный и ахиллов рефлексы в норме хорошо вызываются и достаточно равномерны. Они могут быть резко усилены при общем возбуждении, неврозах и срывах нервной деятельности, а также при интоксикации и общих инфекционных болезнях. С патогенетической точки зрения ослабление рефлексов является следствием нарушения проводимости нервного пути, усиление их указывает на разобщение функции спинного и головного мозга, например при поперечном повреждении спинного мозга, когда сегментарный аппарат разобщается от корригирующего влияния высших центров. Поражение центрального двигательного неврона (пирамидного пути) вызывает усиление сухожильных и периостальных рефлексов. Кожные рефлексы в отличие от глубоких, являясь не врожденными, а приобретаемыми с началом деятельности коры головного мозга, представляют исключение из этого правила. В этом кроется причина обратной реакции при кожных рефлексах, которые в случаях поражения центрального нейрона резко ослабевают или исчезают (А. В. Синев). Патологические изменения рефлексов проявляются клинически или в виде их усиления (повышения) при хронических заболеваниях головного мозга, столбняке, отравлениях, интоксикации, или в виде снижения (ослабления), особенно поверхностных рефлексов. Последнее связывается с ослаблением коркового торможения при острых заболеваниях головного и спинного мозга.
Паракинез (от греч. parakinesis — нарушение покоя) — особая форма гиперкинезов, характеризующаяся непроизвольными сложными движениями. Паракинез наблюдали при ограниченных заболеваниях головного мозга, особенно премоторной зоны полушария, противоположного пораженной области тела. У собак паракинез наблюдали после ограниченных ушибов головного мозга и переболевания чумой. Характерны периодические подергивания лицевыми мускулами, закидывание головы назад, сгибательные или разгибательные движения конечностями. В патогенетическом отношении нужно учитывать возможность перехода симптомов паракинеза в гемиплегию.
Пирамидные гиперкинезы (от греч. Hyper — чрезмерно и kinesis — движение) — чрезмерные, излишние непроизвольные движения, связанные с поражением эфферентных нейронов и двигательных нервных волокон, посредством которых осуществляются произвольные движения. Эти волокна начинаются от передней центральной извилины и оканчиваются в двигательных клетках передних рогов спинного мозга или в ядрах двигательных черепномозговых нервов. Поражение пирамидных путей у низших млекопитающих не вызывает значительных нарушений двигательных функций. Чем выше организовано млекопитающее животное, тем отчетливее выступают эти нарушения. Пирамидные гиперкинезы представляют большой практический интерес и дифференциальной диагностике центральных выпадений при сотрясении и ушибах головного мозга, а также в случаях двигательных расстройств, вызванных черепномозговыми ранениями.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
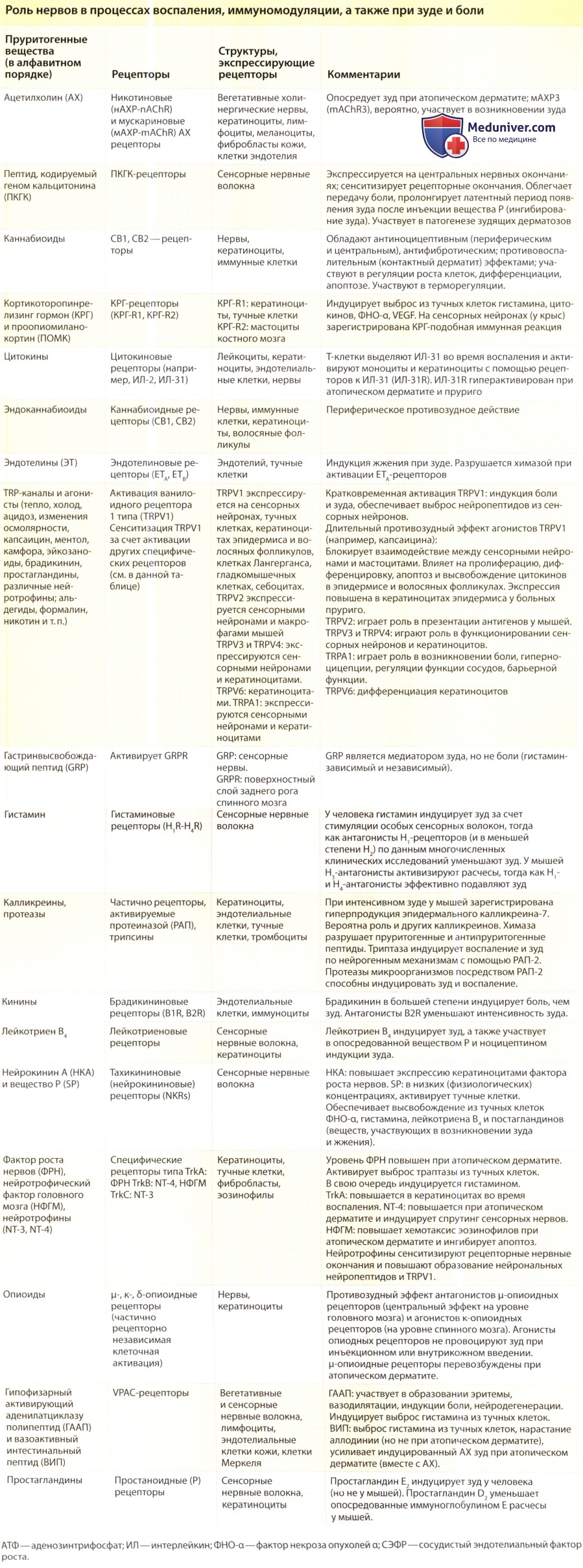
- Нервы кожи, образуя интерактивную сеть, взаимодействуют с различными клетками кожи, эндокринной и иммунной системы.
- Нейродерматологическое взаимодействие влияет на целый ряд таких физиологических и патофизиологических процессов, как рост клеток, воспаление, защитные реакции, зуд и заживление ран.
- Как первичные афферентные, так вегетативные нервные волокна выделяют нейромедиаторы, активирующие специфические рецепторы на многих клетках кожи.
- Клетки кожи экспрессируют ряд специфических рецепторов, которые строго контролируются стимулирующими и подавляющими их активацию сигналами, пептидазами или соседними рецепторами.
- Многочисленные медиаторы (пептиды, протеазы, цитокины, кинины, простаноиды, опиоиды, каннабиноиды, нейтрофины и т.д.), активирующие нейрональные рецепторы, играют важную роль в физиологических и патофизиологических состояниях кожи.
- В спинном мозге и ЦНС осуществляется модуляция сигналов, передаваемых от периферических нервов и относящихся к болевой модальности и зуду; кроме того, происходит обратный процесс—модуляция сигналов, регулирующих кожные функции.
- В настоящее время разрабатываются новые направления лечения различных заболеваний кожи, при которых задействованы нейро-иммуно-эндокринные механизмы.
Эта информация обрабатывается на разных уровнях, в том числе в головном и спинном мозге, дорсальных корневых ганглиях (ДКГ), окончаниях периферических сенсорных нервов, на уровне вегетативных нервов и нейронов и т.д., а также посредством специализированных структур, таких как тельца Пачини, или специализированных клеток, в частности клеток Меркеля.
Эта группа тесно переплетающихся структур и их молекул в итоге играет важнейшую роль в биологии кожи и кожных болезней. Во взаимодействии со спинным и головным мозгом периферические сенсорные нервы выполняют афферентные функции; их окончания воспринимают физические стимулы, такие как прикосновение, тепло или холод, они также выделяют в кожу химические медиаторы и осуществляют эфферентные функции.
Эти сенсорные нервы в значительной мере способствуют развитию кожи до рождения, а также защите и гомеостазу кожи после рождения. Кроме того, вегетативные нервы модулируют как физиологические, так и патофизиологические функции в составе стрессового ответа на экзогенные или эндогенные раздражители и образуют важнейшее звено коммуникации с сосудистой, эндокринной и иммунной системами.
а) Периферические нервы кожи. Сенсорные, а также вегетативные (в коже преимущественно холинергические симпатические) нервы влияют на целый ряд физиологических и патофизиологических функций кожи, таких как эмбриогенез, сужение и расширение сосудов, температура тела, движение мышц, поднимающих волосы; регулировка функции сально-волосяного аппарата, ощущение физических, химических и биологических раздражителей на поверхности кожи; модулирование барьерной функции эпидермиса, клеточная секреция, рост и дифференциация, а также питание и апоптоз клеток; рост нервной ткани, воспаление, иммунная защита и заживление ран.
При отсутствии стимуляции нервов нейромедиаторы в коже практически не определяются. При прямом физическом воздействии — температурном, ультрафиолетовом (УФ), механическом, электрическом, при химической или химико-биологической стимуляции, например, под влиянием аллергенов, гаптенов, микроорганизмов, травмы или воспаления как в условиях in vitro, так и in vivo наблюдается значительное повышение уровня регуляторных нейропептидов, нейротрофинов, нейротрансмиттеров или продуктов окисления (например, оксида озота).
Таким образом, медиаторы, выделяемые сенсорными или вегетативными нервными окончаниями играют в коже важную регуляторную роль при многих физиологических и патологических процессах. Однако наряду с периферической нейромодуляцией, существует сложная, обеспечивающая взаимодействие между спинным мозгом, ЦНС и иммунно-эндокринной системой, сеть, которая также регулирует функции кожи.
Кроме того, нейропептиды и нейтрофины (НТ) могут активироваться и высвобождаться ненейрональными клетками в определенных обстоятельствах, усиливающих или ослабляющих действие нейрогенного раздражителя. В то время как одни нейропептиды [например, вещество Р (SP)] оказывают в ходе воспаления четко выраженное провоспалительное действие, другие нейропептиды, такие как кальцитонин-ген-связанный пептид (CGRP), могут выделяться остро, и, индуцируя (провоспалительное) расширение сосудов, способны одновременно подавлять воспалительные реакции, как при представлении антигенов и иммуносупрессии, чтобы восстановить гомеостаз кожи на более поздней стадии воспаления.
В коже экспрессируется множество рецепторов к этим медиаторам. Большинство из них сопряжены с G-белком. Взаимодействие нейромедиаторов с рецепторами находится под контролем эндопептидазнейтральной эндопептидазы (НЭП), ангиотензинпревращающего фермента (АПФ) и эндотелинпревращающего фермента (ЭПФ), блокирующих индуцированные нейропептидами воспалительные и иммунные реакции. Тесное разнонаправленное взаимодействие между нейромедиаторами, их высокоафинными рецепторами и регуляторными протеазами играет значительную роль в поддержании тканевого гомеостаза и в регуляции патофизиологических процессов в коже.
Ионные каналы неспецифичны и могут быть активированы физическими воздействиями (тепло, холод), химическими веществами (например, капсаицином, ментолом, протонами), а также липидными метаболитами, например простогландинами. Недавно также описана активация ионных каналов каннабиоидами (СВ).
Исходя из этого, очевидно, что чем более разработанной будет сложная проблема нервной системы в коже, тем более доступны окажутся новые перспективные терапевтические подходы к целому ряду дерматологических заболеваний.
Таким образом, доступные данные со всей отчетливостью демонстрируют ключевую роль нейрональной кожной сети как субстрата, влияющего на такие различные физиологические и патофизиологические явления, как защитные реакции, воспаление, зуд, боль, жжение, регенерация и возможно, онкогенез (например, за счет модулирования образования сосудов).
б) Ось головной мозг-кожа. Центральная нервная система соединена с кожей либо прямо, через эфферентные нервы или через производимые в ЦНС медиаторы, либо косвенно, через надпочечники или иммунные клетки. Кожные нервы реагируют также на внутренние стимулы из системного кровообращения (такие как изменения pH, осмотические изменения, брадикинин, цитокины) или на саму кожу и эмоции (внутренние триггерные факторы).
В нормальных условиях сенсорная и вегетативная нервная системы модулируют важные биологические функции, такие как температура тела, кровоток, и рост клеток. Механически индуцированные нервные импульсы передают в ЦНС информацию о давлении. Реагирующие на химические или тепловые стимулы афферентные нервные волокна участвуют в распознании сигналов опасности. Таким образом, в нормальных условиях иннервированная кожа является ключевым барьером, защищающим организм от опасностей во внешней среде. Подтверждением этому служит также факт обильной иннервации не только дермы, но и эпидермиса.
Взаимодействия между центральной и периферической нервной системой играют определенную роль в терморегуляции и патофизиологии различных кожных заболеваний, таких как псориаз, атопический дерматит, акне, заживление ран, а также выпадение волос и возобновление их роста.
в) Спинной мозг и кожа. Наши знания об участии спинного мозга в регулировке специфических ощущений хронической боли в коже человека, о его роли в модулировании зуда или воспалительных стимулов, передаваемых с периферии в головной мозг, очень ограничены. Нам известно, что μ-опиоиды, такие как морфин, могут индуцировать зуд, оказывая обезболивающее действие при интратекальном введении. В отличие от этого, κ-опиоиды оказывают противозудный эффект, вероятно ингибируя рецептор μ-опиоидов. Эти результаты настойчиво указывают на роль опиоидов в регулировке боли и зуда на уровне спинного мозга.
Более того, гастрин-высвобождающий пептид (бомбезин), выделяясь в центральные нервные окончания в спинном мозге, активирует рецептор гастрин-высвобождающего пептида (GRPR) на постсинаптических спинальных нейронах и таким образом селективно регулирует передачу зуда, но не боли. Совокупность этих данных указывает на спинной мозг как на важный регулятор взаимодействий между кожей и нервной системой при воспалении, зудящих заболеваниях и боли, который может стать мишенью для будущих терапевтических модальностей.
г) Анатомия нервной системы кожи:
1. Чувствительные нервы. Большинство нервных структур находится в среднем и в сосочковом слое дермы. Местноспецифические различия наблюдаются на границе слизистых и кожи, в гладкой коже, а также в коже с волосистым покровом. В эпидермисе сенсорные нервные волокна связаны с кератиноцитами, меланоцитами, клетками Лангерганса и Меркеля. Кожные нервы в основном сенсорные, с небольшим включением вегетативных волокон. В отличие от сенсорных вегетативные нервные волокна у млекопитающих никогда не встречаются в эпидермисе. Сенсорные нервы, напротив, иннервируют эпидермис, дерму, а также подкожную жировую клетчатку.
Волокна А-β и А-δ — это в большинстве механочувствительные афференты (типа I) с длительным латентным периодом для теплового раздражения, локализованные на гладкой и имеющей волосяной покров коже. Субпопуляция волокон А-δ на имеющей волосяной покров коже не чувствительна к механическому раздражению (тип II). Около 80% первичных сенсорных нервов кожи представлены А-δ волокнами, распространяющимися из ганглиев дорзальных корешков, тогда как остальные 20% первичных афферентов — это С-волокна.
С-волокна являются либо полимодальными ноцицепторами, которые могут реагировать на химические изменения (с + ), колебания температуры (h + ) или механические стимулы (m + ), либо действуют более специализированно, реагируя только на комбинацию (C-c + h + m + ) или на единственный раздражитель (C-c - h - m + ). Среди периферических нервов человека 45% кожных афферентов относятся к подтипу сенсорных нервов, состоящих из С-волокон, которые одновременно реагируют на механические и термические раздражители (C-m + h + ).
Однако оказалось, что 13% этих нервов являются чувствительными к механическим стимулам (С-m+), 6% — к термическому воздействию (C-h+), 24% — не восприимчивы ни к термическим, ни к механическим стимулам (C-m-h-) и около 12% относятся к вегетативной порции. Среди волокон C-m+h+ 58% реагируют на горчичное масло, в то время как среди С-m + или (С-m - - ) таким свойством обладают лишь 30% волокон, что указывает на существование среди других подтипов также хемо-чувствительных волокон.
Сенсорные нервы воспринимают такие раздражители кожи как тепло, холод или прикосновение. Нервы, воспринимающие тепло, представляют собой преимущественно немиелинизированные С-волокна, субпопуляция волокон А-δ реагирует на легкое охлаждение, в то время как селективные С-волокна активируются при повреждающем холоде. Многие подтипы клеток реагируют на прикосновение и играют важную роль в индукции боли вследствие механического раздражения. Таким образом, система нашего тела имеет как высокоспециализированные, так и менее избирательные ноцицепторы для обеспечения целостности и выживания организма.
Специфическое распределение рецепторов этих разных подтипов сенсорных нервов имеет существенное значение для различных функций (температурной, химической, механической) и воспринимаемых при этом ощущений (пощипывания, покалывания, жжения, боли, зуда). Например, механорецепторы в ганглиях дорзальных корешков (волосяные рецепторы D-типа) экспрессируют исключительно кальциевые каналы Т-типа Ca(v)3.2. Натриевые каналы Nav1.8 (SNS/PN3) и Nav1.9 (SNS/SNS2) экспрессируются как иептидэргическими, так и непептидэргическими нейронами IВ4+(изолектин В4 из Griffonia simplicifolia) и играют решающую роль в определенных подтипах боли. Только первичные афференты небольшого диаметра экспрессируют ванилоидные рецепторы первого типа с транзиторным потенциалом.
Только непептидэргические (слабо пептидэргические) сенсорные волокна экспрессируют пруринергический рецептор Р2х3. Экзогенные факторы, например травма, УФ-излучение, изменения температуры, микробные возбудители, токсины или аллергены, а также эндогенные триггеры воспаления, такие как изменения pH или стрессовые гормональные реакции, способны стимулировать активацию и/или сенситизацию определенных сенсорных нервов. Клеточные процессы, происходящие при передаче действия раздражителя (например, УФ-излучения) до наступления определенной реакции (жгучая боль) в ходе активизации соответствующего механизма (активизация волокон боли, но не волокон зуда, и наоборот) пока еще слабо изучены.
2. Вегетативные (автономные) нервы. В отличие от сенсорных к вегетативным нервам относится меньшая часть кожных нервных волокон. У человека вегетативные нервные волокна кожи практически полностью принадлежат симпатической (холинергической) системе и лишь изредка относятся к парасимпатическим (также холинергическим) структурам. Распространение вегетативных нервов ограничивается дермой, где они иннервируют кровеносные сосуды, артериовенозные анастомозы, лимфатические сосуды, мышцы, подымающие волосы, эккринные и апокринные железы, волосяные фолликулы.
Постганглиарные вегетативные нейроны в коже преимущественно выделяют ацетилхолин, хотя по последним данным дополнительную роль в автономной системе кожи играют нейропептиды. Например, нейропептид Y (НП-Y-NPY) и предсердный натрийуретический пептид образуются исключительно в вегетативных нервных волокнах.
Автономная нервная система кожи регулирует функции потовых желез, регулируя, таким образом, температуру тела, а также водный и электролитный балансы различных органов. Вегетативные нервы участвуют в возникновении целого ряда патологических процессов в коже при гипер- или гипогидрозе, врожденной сенсорной невропатии IV типа, прогрессивном сегментарном гипогидрозе, диабетической нейропатии, сирингомиелии, лепре, дисфункции после симпатэктомии.
Вегетативные нервы осуществляют свое действие в основном путем высвобождения классических нейротрансмиттеров (норадреналина, ацетилхолина) или, в меньшей степени, определенных нейропептидов, таких как вазоактивный интестинальный пептид (ВИП). В отличие от этого, первичные афферентные (сенсорные) нервы выделяют различные классы молекул, в частности, нейропептиды, простаноиды или оксид азота (NO).
Вегетативные нервные волокна активно вовлечены в сосудистые реакции кожи. Симпатические нервы, иннервирующие артериолы, артериовенозные анастомозы и венозные синусоиды, выделяют норадреналин и/или НП-Y (NPY), что приводит к вазоконстрикции, тогда как парасимпатические нервы обусловливают вазодилятацию за счет действия на венозные синусоиды посредством ацетилхолина и вазоактивного интестинального пептида (ВИП)/пептид-гистидин-метионин (ПГМ). Следует отметить, что у ноцицепторов С-волокон может развиться реактивность к адренергическим нейротрансмиттерам в ходе активизации соответствующих рецепторов во время травмы или воспаления. Таким образом, сенсорная и вегетативная нервная системы коммуницируют и взаимодействуют при заболевании на молекулярном уровне.
Мелкие артерии и артериолы, а также артериовенозные анастомозы богато иннервированы норадренергическими нервными окончаниями. В ранее проведенных исследованиях высказывалось предположение, что эта система является холинергической и действует с участием котрансмиттера, вероятно, вазоинтестинального пептида (VIP). Установлено, что холинергические симпатические нервы действуют на мускариновые рецепторы эккринных потовых желез, тогда как высокие концентрации ацетилхолина индуцируют обусловленную аксон-рефлексом эритему за счет никотиновых рецепторов. В системе вазоконстрикции медиатором, скорее всего, является норадреналин и еще один или несколько котрансмиттеров. Лучше других описаны симпатические котрансмиттеры, участвующие в регуляции кровотока, — аденозин трифосфат (АТФ) и NPY.
Примечательно, что даже в отсутствие сенсорной или вегетативной иннервации кожа сохраняет вазодилятационную реакцию, на смену которой приходит неневрогенная вазоконстрикция. Механизмы неневрогенной вазоконстрикции и вазодилятации в ответ на локальное охлаждение остаются неизвестны, однако при этом, вероятно, задействованы оксид азота и сенсорные нервы.
Читайте также:
