
Рефрактерность это отражательная способность цнс
Рефрактерный период обусловлен особенностями поведения потенциал-зависимых натриевых и потенциал-зависимых калиевых каналов возбудимой мембраны.
В ходе ПД, потенциал-зависимые натриевые (Na+) и калиевые (К+) каналы переходят из состояния в состояние. У Na+ каналов основных состояний три - закрытое, открытое и инактивированное. У K+ каналов два основных состояния - закрытое и открытое.
При деполяризации мембраны во время ПД, Na+ каналы после открытого состояния (при котором и начинается ПД, формируемый входящим Na+ током) временно переходят в инактивированное состояние, а K+ каналы открываются и остаются открытыми некоторое время после окончания ПД, создавая выходящий К+ ток, приводящий мембранный потенциал к исходному уровню.
В результате инактивации Na+ каналов, возникает абсолютный рефрактерный период. Позже, когда часть Na+ каналов уже вышла из инактивированного состояния, ПД может возникнуть. Однако для его возникновения требуются очень сильные стимулы, так как, во-первых, "рабочих" Na+ каналов всё ещё мало, а во-вторых, открытые К+ каналы создают выходящий К+ ток и входящий Na+ ток должен его перекрыть, чтобы возник ПД - это относительный рефрактерный период.
Расчёт рефрактерного периода
Рефрактерный период можно рассчитать и описать графически, рассчитав предварительно поведение потенциал-зависимых Na+ и К+ каналов. Поведение этих каналов, в свою очередь, описывается через проводимость и вычисляется через коэффициенты трансфера.

,
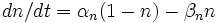
,

- коэффициент трансфера из закрытого в открытое состояние для K+ каналов [1/s];

- коэффициент трансфера из открытого в закрытое состояние для K+ каналов [1/s];
n - фракция К+ каналов в открытом состоянии;
(1 - n) - фракция К+ каналов в закрытом состоянии

,

,

,
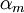
- коэффициент трансфера из закрытого в открытое состояние для Na+ каналов [1/s];

- коэффициент трансфера из открытого в закрытое состояние для Na+ каналов [1/s];
m - фракция Na+ каналов в открытом состоянии;
(1 - m) - фракция Na+ каналов в закрытом состоянии;
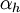
- коэффициент трансфера из инактивированного в не-инактивированное состояние для Na+ каналов [1/s];
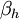
- коэффициент трансфера из не-инактивированного в инактивированное состояние для Na+ каналов [1/s];
h - фракция Na+ каналов в не-инактивированном состоянии;
(1 - h) - фракция Na+ каналов в инактивированном состоянии.
Wikimedia Foundation . 2010 .
- Реформы и порядок (партия)
- Рефрактор
РЕФРАКТЕРНОСТЬ — (от франц. refractaire невосприимчивый) в физиологии отсутствие или снижение возбудимости нерва или мышцы после предшествующего возбуждения. Рефрактерность лежит в основе торможения. Рефрактерный период длится от нескольких десятитысячных (во… … Большой Энциклопедический словарь
рефрактерность — невосприимчивость Словарь русских синонимов. рефрактерность сущ., кол во синонимов: 1 • невосприимчивость (5) Словарь синоним … Словарь синонимов
РЕФРАКТЕРНОСТЬ — (от франц. refractaire невосприимчивый), снижение возбудимости клеток, сопровождающее возникновение потенциала действия. Во время пика потенциала действия возбудимость полностью исчезает (абсолютная Р.) вследствие инактивации натриевых и… … Биологический энциклопедический словарь
рефрактерность — и, ж. refractaire adj. невосприимчивый. физиол. Отсутствие или снижение возбудимости нерва или мышцы после предшествующего возбуждения. СЭС … Исторический словарь галлицизмов русского языка
рефрактерность — (от франц. réfractaire невосприимчивый) (физиол.), отсутствие или снижение возбудимости нерва или мышцы после предшествующего возбуждения. Рефрактерность лежит в основе торможения. Рефракторный период длится от нескольких десятитысячных (во… … Энциклопедический словарь
Рефрактерность — (от франц. геfractaire невосприимчивый) кратковременное снижение возбудимости (См. Возбудимость) нервной и мышечной тканей непосредственно вслед за потенциалом действия (См. Потенциал действия). Р. обнаруживается при стимуляции нервов и… … Большая советская энциклопедия
рефрактерность — (франц. refractaire невосприимчивый) преходящее состояние пониженной возбудимости нервной или мышечной ткани, возникающее после их возбуждения … Большой медицинский словарь
РЕФРАКТЕРНОСТЬ — (от франц. refractaire невосприимчивый) (физиол.), отсутствие или снижение возбудимости нерва или мышцы после предшествующего возбуждения. Р. лежит в основе торможения. Рефракторный период длится от неск. десятитысячных (во ми. нерв. волокнах) до … Естествознание. Энциклопедический словарь
рефрактерность — рефракт ерность, и … Русский орфографический словарь
РЕФРАКТЕРНОСТЬ — [от фр. refraktaire невосприимчивый; лат. refraktarius упрямый] отсутствие или снижение возбудимости нерва или мышцы после предшествующего возбуждения. Р. лежит в основе нервного процесса торможения … Психомоторика: cловарь-справочник
РЕФРАКТЕРНОСТЬ (лат. refractorius невосприимчивый) — состояние возбудимых образований после предшествующего возбуждения, характеризующееся снижением или отсутствием возбудимости. Впервые Р. была обнаружена в мышце сердца Э. Мареем в 1878 г., а в нервах — Готчем и Берком (F. Gotch, С. J. Burck) в 1899 г.
Изменения возбудимости (см.) нервных и мышечных клеток связаны с изменениями уровня поляризации их мембран при возникновении процесса возбуждения (см.). При уменьшении величины мембранного потенциала возбудимость незначительно повышается, а если вслед за уменьшением мембранного потенциала возникает потенциал действия, то возбудимость полностью исчезает и мембрана клетки становится нечувствительной (рефрактерной) к каким бы то ни было воздействиям. Это состояние полной невозбудимости получило название фазы абсолютной Р. Для быстропроводящих нервных волокон теплокровных животных ее продолжительность составляет 0,4 мсек, для скелетных мышц 2,5—4 мсек, для мышц сердца — 250—300 мсек. Восстановление исходного уровня мембранного потенциала сопровождается повышением уровня возбудимости и мембрана приобретает способность реагировать на сверхпороговые раздражители (фаза относительной Р.). В нервных волокнах относительная Р. длится 4—8 мсек, в мышце сердца — 0,03 мсек. Фаза относительной Р. сменяется фазой повышенной возбудимости (экзальтационная фаза Р.), к-рая характеризуется повышением возбудимости против исходного уровня и связана со следовой деполяризацией (отрицательный следовой потенциал). Последующая следовая гиперполяризация (положительный следовой потенциал) сопровождается вторичным снижением возбудимости, к-рая затем сменяется нормальной возбудимостью при восстановлении величины потенциала покоя мембраны.
Все фазы Р. связаны с механизмами возникновения и изменения мембранных потенциалов и обусловлены кинетикой проницаемости мембран для ионов (см. Биоэлектрические потенциалы). Продолжительность фаз Р. можно определить, применяя метод парных раздражений при разных интервалах между ними. Первое раздражение называется кондиционирующим — оно вызывает процесс возбуждения в возбудимой ткани; второе — тестирующее — показывает уровень возбудимости ткани и фазу Р.
На возбудимость и, следовательно, на продолжительность и выраженность отдельных фаз Р. могут оказывать влияние возрастные изменения, воздействие нек-рых лекарственных веществ, температурных и других факторов. Этим пользуются с целью управления возбудимостью ткани при лечении нек-рых заболеваний. Напр., удлинение фазы относительной Р. в мышце сердца приводит к снижению частоты его сокращений и устранению аритмии. Изменения Р., обусловленные нарушением ионных механизмов возникновения возбуждения, наблюдаются при ряде заболеваний нервной системы и мышц.
Библиография: Бериташвили И. С. Общая физиология мышечной и нервной системы, т. 1, М., 1959; Б p е ж e М. А. Электрическая активность нервной системы, пер. с англ., М., 1979; Оке С. Основы нейрофизиологии, пер. с англ., М., 1969; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975, библиогр.; Gotch F. а. В u г с k С. J. The electrical response of nerve to two stimuli, J. Physiol. (Lond.), v. 24, p. 410, 1899.
Нейрофизиология Тесты Синергия
Гиппокамп – это …
основная часть неокортекса
основная часть архикортекса
структура, участвующая в процессах обучения и сохранения памяти
образование, участвующее в высшей координации эмоционального поведения
Функциональные контакты между клетками, передачу сигналов в которых осуществляют специальные вещества посредники – медиаторы – это …
потенциалы действия
стволовые потенциалы
химические синапсы
Поверхность коры больших полушарий человека примерно равна …
500 см. кв.
1700 см. кв.
1000 см. кв.
Ацетилхолин – это …
один из нейромедиаторов
химическое вещество, вызывающее наркотическую зависимость
структура мозга, участвующая в явлениях памяти
Роль натрий-калиевого канала – …
поддерживать клеточный потенциал и регулировать клеточный объём
перемещение ионов в клетку и из клетки
проведение нервного импульса
По функциональному значению выделяют три группы нервных окончаний:
чувствительные, амплитудные, двигательные
чувствительные, двигательные, межнейронные
чувствительные, амплитудные, межнейронные
Места функциональных контактов, образуемых нейронами, называются …
рецепторами
нейронами
синапсами
медиаторами
Кора большого мозга имеет преимущественно…
двухслойное строение
шестислойное строение
трехслойное строение
В коре больших полушарий содержится свыше …
25 млрд. нервных клеток
5 млрд. нервных клеток
14 млрд. нервных клеток
Синапс – это …
протоплазматический мостик-контакт между нейронами
специализированный функциональный контакт между клетками
прорастание аксона в мембрану соседнего нейрона
Нейромедиаторы – это …
вещества, которые выделяются из специальных нейросекреторных клеток
специальные вещества, которые выделяются из постсинаптического нейрона специальные
вещества, которые выделяются из пресинаптической терминали
Нейрофизиология – это …
наука, раскрывающая закономерности функционирования органов и систем
наука, изучающая механизмы, лежащие в основе поведения
наука, изучающая особенности строения нервной ткани
наука о жизнедеятельности нейронов, их ансамблей и нервных сетей
Нейросекреция – это …
образование и выделение нейрогормонов, синтезируемых гипоталамусом
метод регистрации электрической активности головного мозга через неповрежденные покровы головы
выделение медиаторов терминалями аксонов
Мультиполярные (биполярные) клетки – это …
нейроны, имеющие два аксона
элемент сетчатки
нейроны, имеющие аксоны и дендриты
Нервные волокна, отростки нервных клеток, имеющие миелиновую оболочку, – это …
дендрит
серое вещество
белое вещество мозга
аксон
Основателем отечественной школы нейрофизиологии является …
И.П. Павлов
Л.А. Орбели
В.М. Бехтерев
Монополярные клетки – это …
элемент сетчатки
нейроны, имеющие два аксона
нейроны, имеющие только один отросток – аксон
Рефрактерность – это …
кратковременное понижение возбудимости
взаимодействие нейронов в нервных сетях
отражательная способность ЦНС
Часть головного мозга, состоящая из двух полушарий и включающая серое вещество коры, подкорковые ядра, нервные волокна, образующие серое вещество, называются … мозгом
передним
средним
задним
промежуточным
В этой книге предельно сжато изложен курс лекций по нормальной физиологии. Благодаря четким определениям основных понятий студент может сформулировать ответ, за короткий срок усвоить и переработать важную часть информации, успешно сдать экзамен. Курс лекций будет полезен не только студентам, но и преподавателям.
- ЛЕКЦИЯ № 1. Введение в нормальную физиологию
- ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
- ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
- ЛЕКЦИЯ № 4. Физиология мышц
- ЛЕКЦИЯ № 5. Физиология синапсов
- ЛЕКЦИЯ № 6. Физиология центральной нервной системы
- ЛЕКЦИЯ № 7. Физиология различных разделов ЦНС
Приведённый ознакомительный фрагмент книги Нормальная физиология: конспект лекций (С. С. Фирсова) предоставлен нашим книжным партнёром — компанией ЛитРес.
ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
1. Физиологическая характеристика возбудимых тканей
Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей.
Раздражители – это факторы внешней или внутренней среды, действующие на возбудимые структуры.
Различают две группы раздражителей:
1) естественные (нервные импульсы, возникающие в нервных клетках и различных рецепторах);
2) искусственные: физические (механические – удар, укол; температурные – тепло, холод; электрический ток – переменный или постоянный), химические (кислоты, основания, эфиры и т. п.), физико-химические (осмотические – кристаллик хлорида натрия).
Классификация раздражителей по биологическому принципу:
1) адекватные, которые при минимальных энергетических затратах вызывают возбуждение ткани в естественных условиях существования организма;
2) неадекватные, которые вызывают в тканях возбуждение при достаточной силе и продолжительном воздействии.
К общим физиологическим свойствам тканей относятся:
1) возбудимость – способность живой ткани отвечать на действие достаточно сильного, быстрого и длительно действующего раздражителя изменением физиологических свойств и возникновением процесса возбуждения.
Мерой возбудимости является порог раздражения. Порог раздражения – это та минимальная сила раздражителя, которая впервые вызывает видимые ответные реакции. Так как порог раздражения характеризует и возбудимость, он может быть назван и порогом возбудимости. Раздражение меньшей интенсивности, не вызывающее ответные реакции, называют подпороговым;
2) проводимость – способность ткани передавать возникшее возбуждение за счет электрического сигнала от места раздражения по длине возбудимой ткани;
3) рефрактерность – временное снижение возбудимости одновременно с возникшим в ткани возбуждением. Рефрактерность бывает абсолютной (нет ответа ни на какой раздражитель) и относительной (возбудимость восстанавливается, и ткань отвечает на подпороговый или сверхпороговый раздражитель);
4) лабильность – способность возбудимой ткани реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом волн возбуждения, возникающих в ткани в единицу времени (1 с) в точном соответствии с ритмом наносимых раздражений без явления трансформации.
2. Законы раздражения возбудимых тканей
Законы устанавливают зависимость ответной реакции ткани от параметров раздражителя. Эта зависимость характерна для высоко организованных тканей. Существуют три закона раздражения возбудимых тканей:
1) закон силы раздражения;
2) закон длительности раздражения;
3) закон градиента раздражения.
Для совокупности клеток (для ткани) эта зависимость иная, ответная реакция ткани прямо пропорциональна до определенного предела силе наносимого раздражения. Увеличение ответной реакции связано с тем, что увеличивается количество структур, вовлекающихся в ответную реакцию.
Закон длительности раздражений. Ответная реакция ткани зависит от длительности раздражения, но осуществляется в определенных пределах и носит прямо пропорциональный характер. Существует зависимость между силой раздражения и временем его действия. Эта зависимость выражается в виде кривой силы и времени. Эта кривая называется кривой Гоорвега—Вейса—Лапика. Кривая показывает, что каким бы сильным ни был бы раздражитель, он должен действовать определенный период времени. Если временной отрезок маленький, то ответная реакция не возникает. Если раздражитель слабый, то бы как длительно он ни действовал, ответная реакция не возникает. Сила раздражителя постепенно увеличивается, и в определенный момент возникает ответная реакция ткани. Эта сила достигает пороговой величины и называется реобазой (минимальной силой раздражения, которая вызывает первичную ответную реакцию). Время, в течение которого действует ток, равный реобазе, называется полезным временем.
Закон градиента раздражения. Градиент – это крутизна нарастания раздражения. Ответная реакция ткани зависит до определенного предела от градиента раздражения. При сильном раздражителе примерно на третий раз нанесения раздражения ответная реакция возникает быстрее, так как она имеет более сильный градиент. Если постепенно увеличивать порог раздражения, то в ткани возникает явление аккомодации. Аккомодация – это приспособление ткани к медленно нарастающему по силе раздражителю. Это явление связано с быстрым развитием инактивации Na-каналов. Постепенно происходит увеличение порога раздражения, и раздражитель всегда остается подпороговым, т. е. порог раздражения увеличивается.
Законы раздражения возбудимых тканей объясняют зависимость ответной реакции от параметров раздражителя и обеспечивают адаптацию организмов к факторам внешней и внутренней среды.
3. Понятие о состоянии покоя и активности возбудимых тканей
О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма, нет видимого функционального отправления ткани. Состояние активности наблюдается в том случае, когда на ткань действует раздражитель, при этом изменяется уровень метаболизма, и наблюдается функциональное отправление ткани.
Основные формы активного состояния возбудимой ткани – возбуждение и торможение.
Возбуждение – это активный физиологический процесс, который возникает в ткани под действием раздражителя, при этом изменяются физиологические свойства ткани, и наблюдается функциональное отправление ткани. Возбуждение характеризуется рядом признаков:
1) специфическими признаками, характерными для определенного вида тканей;
2) неспецифическими признаками, характерными для всех видов тканей (изменяются проницаемость клеточных мембран, соотношение ионных потоков, заряд клеточной мембраны, возникает потенциал действия, изменяющий уровень метаболизма, повышается потребление кислорода и увеличивается выделение углекислого газа).
По характеру электрического ответа существует две формы возбуждения:
1) местное, нераспространяющееся возбуждение (локальный ответ). Оно характеризуется тем, что:
а) отсутствует скрытый период возбуждения;
б) возникает при действии любого раздражителя, т. е. нет порога раздражения, имеет градуальный характер;
в) отсутствует рефрактерность, т. е. в процессе возникновения возбуждения возбудимость ткани возрастает;
г) затухает в пространстве и распространяется на короткие расстояния, т. е. характерен декремент;
2) импульсное, распространяющееся возбуждение. Оно характеризуется:
а) наличием скрытого периода возбуждения;
б) наличием порога раздражения;
в) отсутствием градуального характера (возникает скачкообразно);
г) распространением без декремента;
д) рефрактерностью (возбудимость ткани уменьшается).
Торможение – активный процесс, возникает при действии раздражителей на ткань, проявляется в подавлении другого возбуждения. Следовательно, функционального отправления ткани нет.
Торможение может развиваться только в форме локального ответ.
Выделяют два типа торможения:
1) первичное, для возникновения которого необходимо наличие специальных тормозных нейронов. Торможение возникает первично без предшествующего возбуждения;
2) вторичное, которое не требует специальных тормозных структур. Оно возникает в результате изменения функциональной активности обычных возбудимых структур.
Процессы возбуждения и торможения тесно связаны между собой, протекают одновременно и являются различными проявлениями единого процесса. Очаги возбуждения и торможения подвижны, охватывают большие или меньшие области нейронных популяций и могут быть более или менее выражены. Возбуждение непременно сменяется торможением, и наоборот, т. е. между торможением и возбуждением существуют индукционные отношения.
4. Физико-химические механизмы возникновения потенциала покоя
Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов К, снаружи его мало. Ионов Na и ионов Cl больше снаружи, чем внутри. Такое распределение ионов называется ионной асимметрией;
2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ.
За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концентрации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил:
1) силы диффузии;
2) силы электростатического взаимодействия.
Значение электрохимического равновесия:
1) поддержание ионной асимметрии;
2) поддержание величины мембранного потенциала на постоянном уровне.
В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентрационно-электрохимическим.
Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ. Работа натрий-калиевого насоса обеспечивает:
1) высокую концентрацию ионов К внутри клетки, т. е. постоянную величину потенциала покоя;
2) низкую концентрацию ионов Na внутри клетки, т. е. сохраняет нормальную осмолярность и объем клетки, создает базу для генерации потенциала действия;
3) стабильный концетрационный градиент ионов Na, способствуя транспорту аминокислот и сахаров.
5. Физико-химические механизмы возникновения потенциала действия
Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Компоненты потенциала действия:
1) локальный ответ;
2) высоковольтный пиковый потенциал (спайк);
3) следовые колебания:
а) отрицательный следовой потенциал;
б) положительный следовой потенциал.
Пока раздражитель не достиг на начальном этапе 50–75 % от величины порога, проницаемость клеточной мембраны остается неизменой, и электрический сдвиг мембранного потенциала объясняется раздражающим агентом. Достигнув уровня 50–75 %, открываются активационные ворота (m-ворота) Na-каналов, и возникает локальный ответ.
Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку. Если сила раздражения недостаточна, то локального ответа не происходит.
Высоковольтный пиковый потенциал (спайк).
Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз:
1) восходящей части – фазы деполяризации;
2) нисходящей части – фазы реполяризации.
Лавинообразное поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Постепенно заряд с мембраны снимается, а потом возникает с противоположным знаком. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na. Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K, натрий-калиевый насос вступает в действие и восстанавливает заряд клеточной мембраны. Полного восстановления мембранного потенциала не происходит.
В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный. Следовые потенциалы являются непостоянными компонентами потенциала действия. Отрицательный следовой потенциал – следовая деполяризация в результате повышенной проницаемости мембраны к ионам Na, что тормозит процесс реполяризации. Положительный следовой потенциал возникает при гиперполяризации клеточной мембраны в процессе восстановления клеточного заряда за счет выхода ионов калия и работы натрий-калиевого насоса.
- ЛЕКЦИЯ № 1. Введение в нормальную физиологию
- ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
- ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
- ЛЕКЦИЯ № 4. Физиология мышц
- ЛЕКЦИЯ № 5. Физиология синапсов
- ЛЕКЦИЯ № 6. Физиология центральной нервной системы
- ЛЕКЦИЯ № 7. Физиология различных разделов ЦНС
Приведённый ознакомительный фрагмент книги Нормальная физиология: конспект лекций (С. С. Фирсова) предоставлен нашим книжным партнёром — компанией ЛитРес.
Рефрактерность. Рефрактерность - временное снижение возбудимости ткани, возникающее при появлении потенциала действия. В этот момент повторные раздражения не вызывают ответной реакции (абсолютная рефрактерность). Она длится не более 0,4 миллисекунды, а затем наступает фаза относительной рефракторности, когда раздражение может вызвать слабую реакцию. Эта фаза сменяется фазой повышенной возбудимости - супернормальности. Показатель рефрактерности (рефрактерный период) - время, в течение которого возбудимость ткани снижена. Рефрактерный период тем короче, чем выше возбудимость ткани.
Процесс возбуждения сопровождается изменением возбудимости. Таков смысл свойства рефрактерности. Это слово, в переводе означающее невпечатлительность, ввел в науку Э. Ж. Марей, обнаруживший в 1876 г. угнетение возбудимости миокарда в момент его возбуждения. Позднее рефрактерность была выявлена во всех возбудимых тканях. В1908 г. Н. Е. Введенский установил, что вслед за угнетением наступает некоторое повышение возбудимости возбужденной ткани.
Выделяют три основные стадии рефрактерности, их принято называть фазами:
- Развитие возбуждения вначале сопровождается полной утратой возбудимости (е = 0). Это состояние называют абсолютно рефрактерной фазой. Она соответствует времени деполяризации возбудимой мембраны. В течение абсолютно рефрактерной фазы возбудимая мембрана не может генерировать новый потенциал действия, даже если на нее подействовать сколь угодно сильным раздражителем (S„-> оо). Природа абсолютно рефрактерной фазы состоит в том, что во время деполяризации все потенциалзависимые ионные каналы находятся в открытом состоянии, и дополнительные стимулы не могут вызвать воротный процесс (им просто не на что действовать).
- Относительно рефрактерная фаза – возвращает возбудимость от нуля к исходному уровню (е0). Относительно рефрактерная фаза совпадает с реполяризацией возбудимой мембраны. С течением времени во все большем числе потенциалзависимых ионных каналов завершаются воротные процессы, с которыми было связано предшествующее возбуждение, и каналы вновь обретают способность к следующему переходу из закрытого в открытое состояние под действием очередного стимула. Вовремя относительно рефрактерной фазы пороги возбуждения постепенно снижаются (S„o
- Фаза экзальтации, для которой характерна повышенная возбудимость (е> е0). Она, очевидно, связана с изменением свойств сенсора напряжения во время возбуждения. За счет перестройки конформации белковых молекул изменяются их дипольные моменты, что приводит к повышению чувствительности сенсора напряжения к сдвигам мембранного потенциала (критический мембранный потенциал приближается к потенциалу покоя).
Разным возбудимым мембранам присуща неодинаковая продолжительность каждой фазы рефрактерности. Так, в скелетных мышцах АРФ длится в среднем 2,5 мс, ОРФ — около 12 мс, ФЭ — приблизительно 2 мс. Миокард отличается гораздо более продолжительной АРФ — 250—300 мс, что обеспечивает четкую ритмичность сердечных сокращений и является необходимым условием жизни. В типичных кардиомиоцитах относительно рефрактерная фаза длится около 50 мс, а в сумме продолжительность абсолютно рефрактерной и относительно рефрактерной фаз примерно равна длительности потенциала действия. Различия в длительности рефракторных фаз обусловлены неодинаковой инерционностью потенциалзависимых ионных каналов. В тех мембранах, где возбуждение обеспечивается натриевыми каналами, рефрактерные фазы наиболее быстротечны и потенциал действия наименее продолжителен (порядка единиц миллисекунд). Если же за возбуждение ответственны кальциевые каналы (например, в гладких мышцах), то рефрактерные фазы затягиваются до секунд. В сарколемме кардиомиоцитов присутствуют и те, и другие каналы, вследствие чего длительность рефрактерных фаз занимает промежуточное значение (сотни миллисекунд).
Рефрактерный период в возбудимых клетках
В фазу деполяризации потенциала действия потенциалзависимые натриевые ионные каналы кратковременно открываются, но затем инактивируются h-ворота. В период инактивации натриевых ионных каналов возбудимые клетки не способны реагировать повышением натриевой проницаемости на повторный стимул. Поэтому во время фазы деполяризации мембрана не может генерировать потенциал действия в ответ на действие пороговых или сверхпороговых раздражителей. Это состояние называется абсолютной рефрактерностью, время которой составляет в нервных волокнах 0,5—1,0 мс, а в скелетных мышечных клетках —в среднем 2 мс. Период абсолютной рефрактерности заканчивается после того, как снижается количество инактивированных натриевых каналов и постепенно увеличивается количество натриевых каналов, находящихся в закрытом состоянии. Происходят эти процессы во время фазы реполяризации, когда уменьшению количества потенциалзависимых натриевых ионных каналов, находящихся в состоянии инактивации, соответствует период относительной рефрактерности. Период относительной рефрактерности характеризуется тем, что лишь некоторая часть потенциалзависимых натриевых ионных каналов переходит в закрытое состояние, а в силу этого порог возбудимости мембраны клетки имеет более высокие значения, чем в исходном состоянии. Поэтому возбудимые клетки в период относительной рефрактерности могут генерировать потенциалы действия, но при воздействии на них раздражителей сверхпороговой силы. Однако из-за небольшого количества потенциалзависимых натриевых ионных каналов, находящихся в закрытом состоянии, амплитуда генерируемых при этом потенциалов действия будет меньше, чем в условиях исходной возбудимости нервной или мышечной клетки.
В клетках возбудимых тканей максимальное количество генерируемых потенциалов действия в единицу времени обусловлено двумя факторами: длительностью потенциала действия и длительностью периода абсолютной рефрактерности после каждого импульса. На этом основании в физиологии формулируется современное понятие лабильности: чем меньше период абсолютной рефрактерности при возбуждении возбудимой ткани, тем выше ее функциональная подвижность или лабильность, тем больше в ней генерируется потенциалов действия в единицу времени.
При непрерывной стимуляции нерва электрическим током лабильность нерва зависит от частоты и силы раздражения. В зависимости от частоты и силы раздражения нерва сокращение иннервируемой им мышцы может быть максимальной или минимальной амплитуды. Эти явления были названы соответственно оптимумом и пессимумом (Н. Е. Введенский). Максимальное (оптимально большое) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв в периоде его состояния супернормальной возбудимости после предыдущего потенциала действия. Минимальное (или пессимальное) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв, находящийся в периоде относительной рефрактерности после предыдущего потенциала действия. Поэтому значения оптимальной частоты раздражения нерва всегда меньше, чем значения пессимальной частоты раздражения.
К мерам возбудимости относятся:
- Порог раздражения - первая базисная мера раздражителя любой природы. Но для количественной оценки возбудимости в медицине используют не любой раздражитель, а используют электрический ток. Именно с помощью электрического тока тестируют мышцы, нервы, синапсы. Электрический ток точно дозируется - электрический ток можно легко дозировать, при чем по двум показателям: по силе и по времени действия. С другими раздражителями иначе: например, химический - можно дозировать по силе (концентрации), но нельзя - по длительности, так как для его отмывания нужно время. С помощью электрического тока получены еще 3 меры возбудимости, одна из которых используется в медицине:
- Базисная мера - это реобаза - минимальная сила постоянного тока, которая, действуя длительное, но определенное время, способна вызвать ответную реакцию. Недостаток этой меры - определение времени трудно определимо - оно расплывчато.
- Полезное время - то время, которое должна действовать сила тока в 1 реобазу, чтобы вызвать ответную реакцию. Но и эта мера возбудимости не нашла своего применения в медицинской практике, потому что, как показывает график, она находится на очень пологой части кривой "сила - время" и любая неточность (небольшая неточность) вела к большой ошибке.
- Хронаксия - минимальное время, в течение которого должна действовать сила тока в 2 реобазы, чтобы вызвать ответную реакцию. На графике - это тот участок кривой, где зависимость между силой и временем точно прослеживается. Посредством хронаксии определяют возбудимость нервов, мышц, синапсов. Этим методом определяют, где же наступило поражение нервно-мышечной системы: на уровне мышцы, нервов, синапсов или центральных образований.
Дата добавления: 2015-08-12 ; просмотров: 11468 . Нарушение авторских прав
Читайте также:
